Abstract
Objectives: Sepsis is a life-threatening condition characterized by multi-organ dysfunction due to host immune system dysregulation in response to an infection. During sepsis, neutrophils release neutrophil extracellular traps (NETs) as part of the innate immune response. However, excessive NETs play a critical role in the development of organ failure during sepsis. Although recombinant human soluble thrombomodulin (rTM) can inhibit NET formation in the lungs and liver of a mouse model of endotoxin shock, its effects on the kidneys are unclear.
Methods: The specific effects of NETs and rTM on the renal cortex and renal medulla were examined in a mouse model of endotoxin shock generated by intraperitoneal (i.p.) injection of lipopolysaccharide (LPS), followed by i.p. injection of rTM or an identical volume of saline 1 h later.
Results: LPS injection increased serum creatinine, blood urea nitrogen, and histone H3 levels. However, rTM administration significantly decreased histone H3 and citrullinated histone H3 (citH3) levels. Immunohistochemical analysis revealed no significant changes in citH3 quantity in the renal cortex of any group. However, in the renal medulla, the increase in citH3 induced by LPS was abolished in the LPS+rTM group.
Conclusions: Our findings demonstrate that rTM can suppress NETs in the renal medulla of mice with endotoxin-induced acute kidney injury.
Introduction
Sepsis, a type of multi-organ dysfunction, results from infection-induced dysregulation of immune responses.1 A recent study revealed that hospital mortality rates associated with sepsis and severe sepsis in the United States are 17% and 26%, respectively.2 In addition, sepsis can cause acute kidney injury (AKI),3,4 and considerably increases the duration of hospital stays and in-hospital mortality rates compared with non-septic AKI.5 However, the pathophysiology of AKI in sepsis remains unknown, and better understanding could improve treatment strategies for sepsis.
As a part of the innate immune response, neutrophils release substances such as histones and nuclear DNA, which form reticular structures called neutrophil extracellular traps (NETs).6 As these reticular structures trap and kill bacteria,7 they affect coagulation and thrombogenesis.8,9 NET formation contributes to immune thrombosis during the pathogenesis of sepsis.9 However, excessive NET formation can lead to capillary circulation disorders.10,11 During sepsis, rapid systemic inflammation causes these immune responses to affect both infected and non-infected regions of the body, leading to multiple-organ dysfunction.12 NETs release citrullinated histones, which induce platelet aggregation13,14 and inflammation.15 Direct histone injection into the kidney has been shown to cause neutrophil migration, microvascular leakage, and renal inflammation.16
Thrombomodulin (TM) is an important cofactor in the anticoagulant pathway that exerts anticoagulation effects in vivo, however, its expression is decreased during the development of septic-disseminated intravascular coagulation.17 Recombinant human soluble TM (rTM) can inhibit NET formation by binding to histones.18 Previously, we reported that injecting rTM into a mouse model of endotoxin shock improved the survival rate and inhibited NET formation in both lung and liver tissues.19 However, the effects of rTM on the kidney remain controversial. Hayase et al. found that rTM administration inhibited histone accumulation and NET formation in the lungs, but not kidneys, of a mouse model of renal ischemia-reperfusion injury.20 In contrast, Nozaki et al. and Akatsuka et al. reported that rTM reduces renal damage.21,22 The kidney has multiple physiological roles: the glomerulus and glomerular capillary produce primary urine in the renal cortex, whereas the nephron and peritubular capillary reabsorb nutrients from primary urine in the renal medulla. Notably, previous studies examined the kidney as a single organ, with no distinction made between the renal cortex and renal medulla. The immune function of healthy renal medulla is more developed than that of the renal cortex, as antigen-presenting dendritic cells are more prevalent in the peritubular interstitium.23 Moreover, the high-sodium environment of the renal medulla induces chemokines that promote immune function.24 We suspected that the renal medulla plays an important role in the immune response during sepsis as a result of its well-developed immune function. Therefore, we hypothesized that NETs are abundant in the renal medulla of endotoxin-shocked mice, and rTM can effectively reduce NETs. In this study, we aimed to evaluate the specific effects of rTM on the renal cortex and renal medulla in an endotoxin shock mouse model.
Methods
Animals and study design
Female C57BL/6J Jms mice were obtained from SLC (Hamamatsu, Japan) and maintained in our conventional mouse and rat facility with free access to food and water. Mice aged 6–8 weeks were used after at least 6 days of acclimation. Lipopolysaccharide (LPS; 125-05201, Wako Pure Chemical Industries, Osaka, Japan) and rTM (ART-123; Asahi Kasei Pharma, Tokyo, Japan) were used in the present study. To create the endotoxin shock model, 10 mg/kg LPS was intraperitoneally (i.p.) injected into mice. One hour after LPS injection, 6 mg/kg rTM (rTM group) or an equal volume of saline (non-rTM group) was administered (i.p.). Control mice were injected with an identical volume of saline instead of LPS or rTM. Eight hours after LPS/saline injection, mice were anesthetized with isoflurane and euthanized by cardiac puncture. Blood and kidney samples were collected for analysis. Symptoms of sepsis, such as lethargy, piloerection, decreased appetite, and a hunched position, were also observed.
Ethics statement
All mice were handled according to Regulations for the Management of Laboratory Animals at Fujita Health University (Toyoake, Japan). All animal protocols were approved by the Animal Care and Use Committee of Fujita Health University (Approval No. APU19079-MD1, 5 January 2022). The point when mice displayed loss of the righting reflex while in a dorsal position was classified as the humane endpoint, at which point mice were culled.
Measurement of histone H3 (H3) and citrullinated H3 (citH3) levels
Serum was separated from collected blood samples by centrifugation at 1470 × g for 10 min at 4°C. Serum H3 levels were measured using enzyme-linked immunosorbent assay (ELISA) with human anti-H3 antibodies (Shino-Test Corporation, Sagamihara, Japan) as previously described.25 citH3 was detected using an ELISA kit (501620; Cayman Chemical, Ann Arbor, MI, USA).
Measurement of blood creatinine (CRE) and blood urea nitrogen (BUN) levels
Dri-CHEM (NX600; Fujifilm, Tokyo, Japan) was used to measure CRE (v-CRE-P, Fujifilm) and BUN (v-BUN-P, Fujifilm) levels. Slides were placed in the machine and 10 μL of serum was dropped onto the slides. Spotting was carried out manually using a pipette because the sample volume was small.
Immunofluorescence staining
Excised kidneys were immediately fixed in 4% paraformaldehyde phosphate buffer solution (Fujifilm), and 3-μm-thick sections were prepared after paraffin embedding. Sections were incubated with primary antibodies against lymphocyte antigen 6 complex locus G6D (Ly-6G; 551459; BD Bioscience, Franklin Lakes, NJ, USA) and histone H3 (citrulline R2+R8+R17, citH3; ab5103; Abcam, Cambridge, UK), followed by Alexa Fluor 488- (ab172332, Abcam) and Alexa Fluor 594-conjugated (A21207; Thermo Fisher Scientific, Waltham, MA, USA) secondary antibodies. Nuclei were stained using 4',6-diamidino-2-phenylindole dihydrochloride solution (DAPI; Thermo Fisher Scientific). Tissue sections were observed and imaged using a confocal laser-scanning microscope (LSM 980; Zeiss, Oberkochen, Germany).
Quantification of citH3 under immunohistochemical staining
Stained kidneys were imaged at 400× magnification with an LSM 980 (Zeiss). Numbers of citH3-positive cells were quantified in the renal cortex and renal medulla. In the renal cortex, three images were taken per sample, citH3-positive cells in the glomerulus were counted, and the mean value was calculated. In the renal medulla, a single image was taken per sample and citH3-positive cells in the field of view were counted. Fiji (ImageJ, version 1.53q; http://imagej.nih.gov)26 was used for manual quantification.
Statistical analysis
Serum CRE, BUN, H3, and citH3 levels were analyzed using the Mann–Whitney U test. Numbers of citH3-positive cells evaluated by immunohistochemical staining were analyzed using an unpaired t-test. Results with P<0.05 were considered statistically significant. All statistical analyses were conducted using GraphPad Prism version 9.3.1 (GraphPad Software, San Diego, CA, USA).
Results
Effects of rTM on Serum H3 and citH3 Levels during Endotoxin Shock
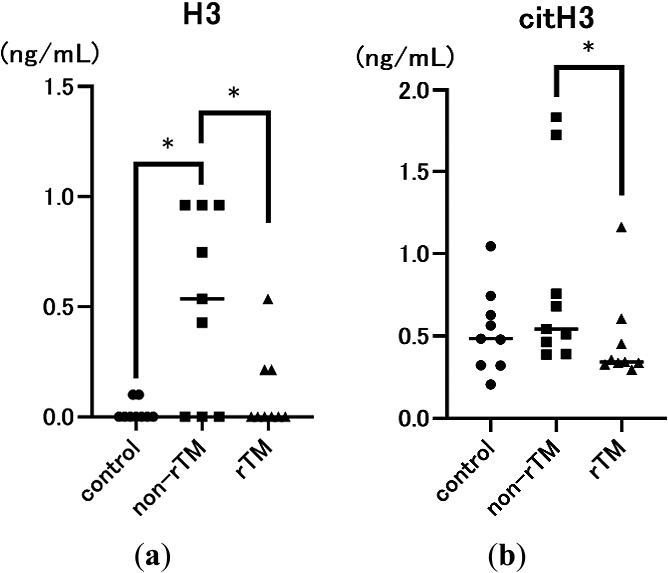
First, we examined the effects of rTM on serum H3 and citH3 levels in the mouse model 8 h after LPS injection. LPS significantly increased serum H3 levels (H3: 0.535 mg/dL, p=0.0202; citH3: 0.542 mg/dL) compared with the control (H3: 0.000 mg/dL; citH3: 0.484 mg/dL). However, rTM administration significantly decreased serum H3 and citH3 levels (H3: 0.000 mg/dL, p=0.0493; citH3: 0.342 mg/dL, p=0.0179; Figure 1) following LPS induction.
Effects of rTM on blood CRE and BUN levels during endotoxin shock
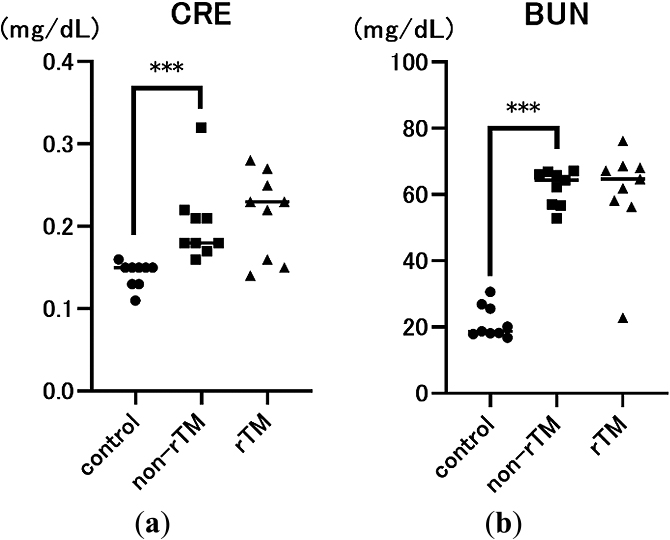
To determine the effects of LPS and rTM, we also measured two indicators of endotoxin-induced renal injury, serum CRE and BUN levels, 8 h after LPS administration. As expected, CRE and BUN levels were significantly higher in the non-rTM group (CRE: 0.18 mg/dL, p<0.0001; BUN: 64.4 mg/dL, p<0.0001) compared with the control group (CRE: 0.15 mg/dL; BUN: 18.7 mg/dL). However, there was no significant difference in CRE or BUN levels between non-rTM and rTM groups (CRE: 0.23 mg/dL; BUN: 64.7 mg/dL, Figure 2).
Effects of rTM on NETs in kidneys during endotoxin shock
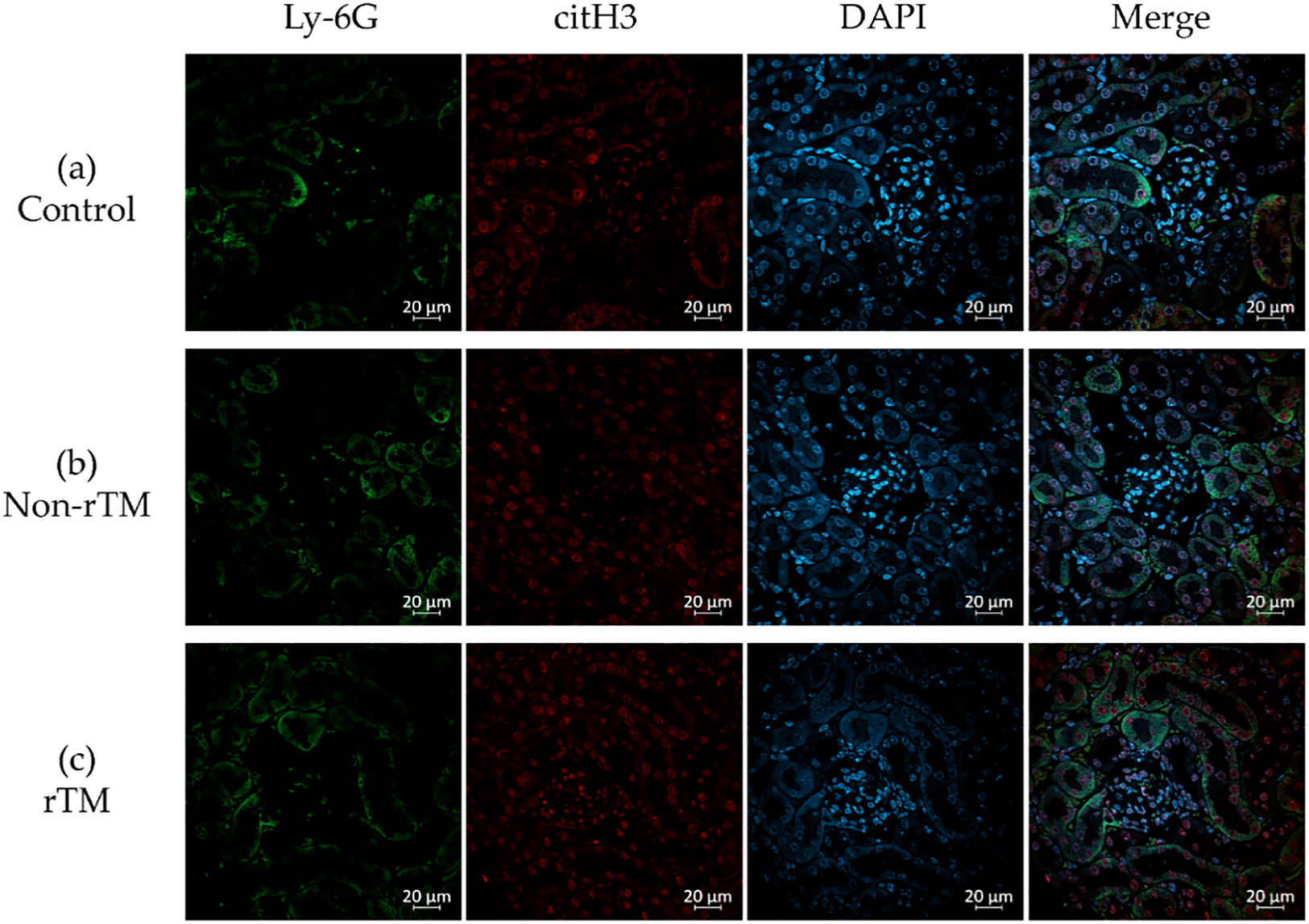
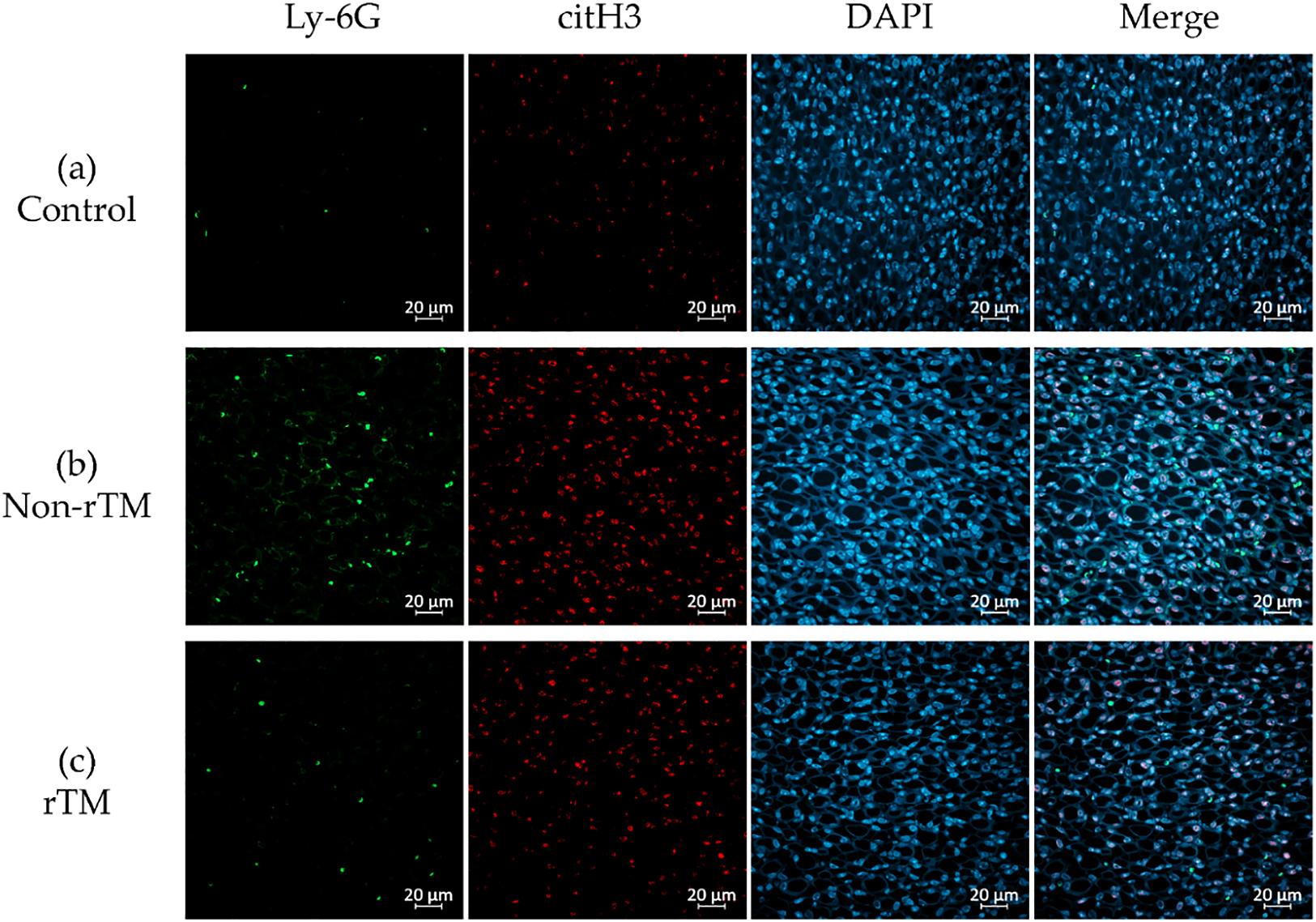
To confirm the effects of rTM on NETs in the kidney, the left kidney was removed 8 h after LPS administration and subjected to immunofluorescence staining to identify neutrophils (Ly-6G, green) and NETs (citH3, red). Immunohistochemical analysis revealed no significant changes in citH3 in the renal cortex between control (21.87 count/glomerulus) and non-rTM groups (23.57 count/glomerulus), or between non-rTM and rTM groups (24.77 count/glomerulus, Figures 3 and 5). In the renal medulla, citH3 was increased in the non-rTM group (317.3 count/HPF, p=0.0144) compared with the control group (193.0 count/HPF) and decreased in the rTM group (253.0 count/HPF, p=0.0352, Figures 4 and 5). Notably, citH3 luminescence was observed in both the renal cortex and renal medulla of the control group, suggesting the possibility of autoluminescence.
Discussion
Sepsis is a life-threatening condition characterized by multi-organ dysfunction due to dysregulation of the host immune system in response to an infection.1 During sepsis, neutrophils release NETs as part of the innate immune response, which can trap pathogens and prevent them from spreading. However, an excessive NET response can induce microvascular occlusion and tissue damage.8,27 Previously, we reported that rTM inhibits LPS-induced NETs in vitro in human neutrophils and platelets,28 and rTM injection can decrease lethality in mice with endotoxin shock to inhibit NET formation in the lungs and liver.19 However, the effects of rTM on the kidneys remain unclear. Because the kidney has various physiological roles, we evaluated the specific effects of rTM on the renal cortex and renal medulla of mice with endotoxin shock. First, we confirmed that LPS significantly increased serum CRE, BUN, and H3 levels compared with the control, suggesting that LPS successfully induced AKI in mice (Figures 1a and 2). In addition, we found that rTM significantly decreased serum H3 and citH3 levels in LPS-treated mice, suggesting that it effectively alleviated endotoxin shock (Figure 1).
Organs that are in contact with the external environment take up essential substances and excrete unwanted substances; however, bacteria and toxins can inadvertently be taken up at the same time. To protect organisms from external bacteria and toxins, their organs have well-developed immune functions.29,30 For instance, the lungs take up oxygen while discharging carbon dioxide, and contain both innate and adaptive immune cells.29. Similarly, the liver both absorbs and excretes substances from/into the intestinal tract via the enterohepatic circulatory system, and is therefore continuously exposed to antigenic stimuli including exogenous pathogens from the intestinal tract, dietary components, and foreign biological substances such as drugs and toxins.30 Although the lungs and liver have well-developed immune functions and rich blood flow, they are both low-pressure organs (Figure 6);31–37 this characteristic is hypothesized to facilitate interactions with foreign substances to allow leukocyte function. This hypothesis is consistent with the findings of our previous study, in which we demonstrated that NETs form in the lungs and liver of mice with endotoxin shock.19 Although pulmonary, sinusoidal, and peritubular capillaries have low perfusion pressure, glomerular capillaries have high perfusion pressure (Figure 6),31–40 suggesting that the kidney is a high-pressure organ. However, at the sub-organ level, the renal cortex is associated with high pressure and the renal medulla is associated with low pressure. Indeed, approximately 1 L of blood flows into the kidneys every minute, with 90% of blood flowing through the renal cortex,41 which filters out unwanted toxins as primary urine; in contrast, the medulla reabsorbs essential substances. In this study, no significant changes in citH3 quantity were observed in the renal cortex of any group (Figure 3, 5a), likely because the renal cortex is a high-pressure organ specialized for filtering urine.
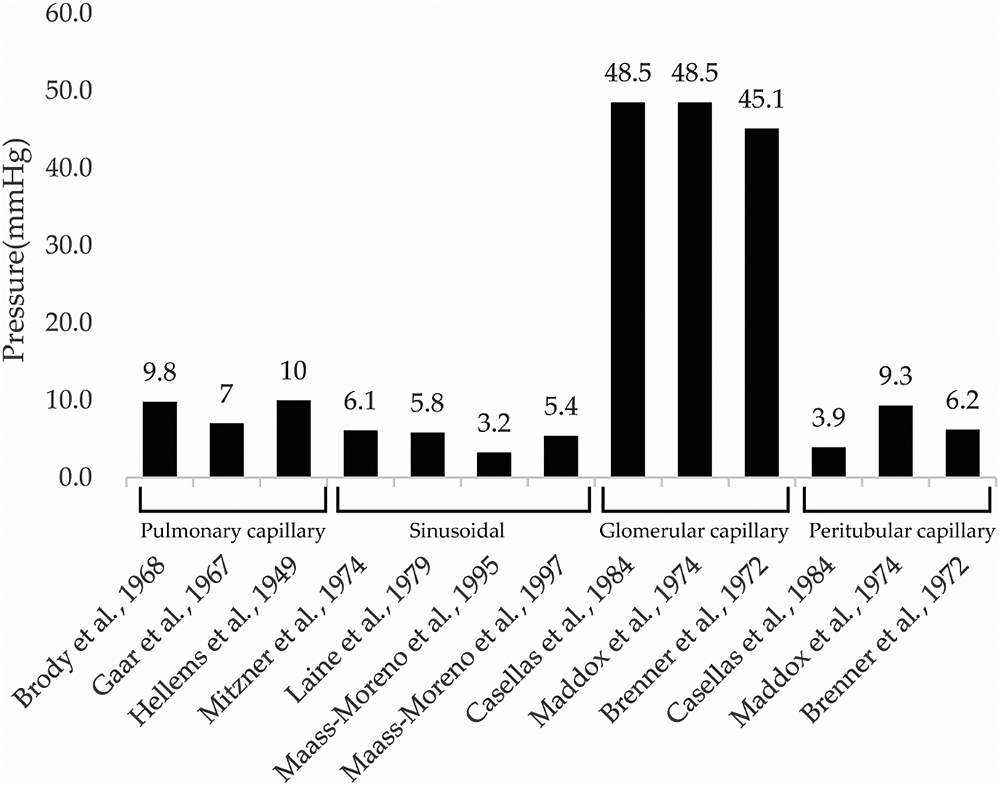
Unlike the renal cortex, the renal medulla absorbs essential substances that constitute around 99% of the primary urine (100 L) produced by the glomerulus per day. Glomerular capillaries have pores with an approximately 60-nm diameter that rapidly filter out molecules <20 kDa, while partially or completely filtering out larger molecules. Previous studies reported that LPS accumulates in the proximal tubules,42 whereas damage-associated molecular patterns (DAMPs), such as histones, are filtered out by the glomerulus. The receptors of these molecules, Toll-like receptors 2 and 4, are located in the tubular epithelium and can cause inflammation upon binding.43,44 Peritubular capillaries wrap around the tubules of the renal medulla and reabsorb substances from the primary urine. The renal medulla has a highly developed immunity, largely due to the presence of many antigen-presenting dendritic cells in the peritubular interstitium.23 Accordingly, the renal medulla can induce high chemokine levels24 to prevent DAMPs and LPS from re-entering the peritubular capillaries during advanced sepsis. Here, we found that LPS increased citH3 in the renal medulla (Figures 4a, b; 5b), as hypothesized. Thus, we believe that the strong immune function of the renal medulla, as well as its low pressure, may facilitate the accumulation of NETs. Furthermore, we found that rTM suppressed citH3 (Figures 4b, c; 5b), suggesting that rTM can effectively inhibit NETs in the renal medulla. One limitation of this study is that the effect of rTM was explored only with regard to neutrophils, although it can bind to other immune cells. Although an endotoxin shock model was used, this study focused on renal injury, and only the effect on renal NETs on rTM was evaluated. Therefore, it is possible that other immune effectors not targeted by rTM may contribute to kidney injury after LPS administration. Indeed, this study did not reveal whether NETs formed in the kidney, but instead focused on whether rTM could suppress NETs in the kidney following induction by LPS.
Acknowledgments
We would like to thank Kanako Kumamoto (Fujita Health University, Toyoake, Japan) for her technical assistance with immunohistochemistry experiments. We are grateful to Kazuya Shiogama (Fujita Health University) for his advice on the pathology findings of the kidney.
Notes
Funding
This research was funded by JSPS KAKENHI (Grant Nos. 17K11064, 19K18369, 20K09233, and 20K17878).
References
- 1. Singer M, Deutschman CS, Seymour CW, et al. The third international consensus definitions for sepsis and septic shock (sepsis-3). JAMA 2016; 315: 801–810.
- 2. Fleischmann C, Scherag A, Adhikari NKJ, Hartog CS, Tsaganos T, Schlattmann P, Angus DC, Reinhart K. Assessment of global incidence and mortality of hospital-treated sepsis current estimates and limitations. Am J Respir Crit Care Med 2016; 193: 259–272.
- 3. Uchino S, Kellum JA, Bellomo R, Doig GS, Morimatsu H, Morgera S, Schetz M, Tan I, Bouman C, Macedo E, Gibney N, Tolwani A, Ronco C. Acute renal failure in critically ill patients: A multinational, multicenter study. JAMA 2005; 294: 813–818.
- 4. Bouchard J, Acharya A, Cerda J, Maccariello ER, Madarasu RC, Tolwani AJ, Liang X, Fu P, Liu ZH, Mehta RL. A prospective international multicenter study of AKI in the intensive care unit. Clin J Am Soc Nephrol 2015; 10: 1324–1331.
- 5. Bagshaw SM, Uchino S, Bellomo R, Morimatsu H, Morgera S, Schetz M, Tan I, Bouman C, Macedo E, Gibney N, Tolwani A, Oudemans-van Straaten HM, Ronco C, Kellum JA. Septic acute kidney injury in critically ill patients: Clinical characteristics and outcomes. Clin J Am Soc Nephrol 2007; 2: 431–439.
- 6. Dąbrowska D, Jabłońska E, Garley M, Ratajczak-Wrona W, Iwaniuk A. New aspects of the biology of neutrophil extracellular traps. Scand J Immunol 2016; 84: 317–322.
- 7. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y, Zychlinsky A. Neutrophil extracellular traps kill bacteria. Science 2004; 303: 1532–1535.
- 8. Denning NL, Aziz M, Gurien SD, Wang P. Damps and nets in sepsis. Front Immunol 2019; 10: 2536.
- 9. Kapoor S, Opneja A, Nayak L. The role of neutrophils in thrombosis. Thromb Res 2018; 170: 87–96.
- 10. Grayson PC, Kaplan MJ. At the bench: Neutrophil extracellular traps (NETs) highlight novel aspects of innate immune system involvement in autoimmune diseases. J Leukoc Biol 2016; 99: 253–264.
- 11. Ito T. PAMPs and DAMPs as triggers for DIC. J Intensive Care 2014; 2: 67.
- 12. Cheng Z, Abrams ST, Toh J, Wang SS, Wang Z, Yu Q, Yu W, Toh C-H, Wang G. The critical roles and mechanisms of immune cell death in sepsis. Front Immunol 2020; 11: 1918.
- 13. Fuchs TA, Bhandari AA, Wagner DD. Histones induce rapid and profound thrombocytopenia in mice. Blood 2011; 118: 3708–3714.
- 14. Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD, Wrobleski SK, Wakefield TW, Hartwig JH, Wagner DD. Extracellular DNA traps promote thrombosis. Proc Natl Acad Sci U S A 2010; 107: 15880–15885.
- 15. Silk E, Zhao H, Weng H, Ma D. The role of extracellular histone in organ injury. Cell Death Dis 2017; 8: e2812.
- 16. Allam R, Scherbaum CR, Darisipudi MN, et al. Histones from dying renal cells aggravate kidney injury via TLR2 and TLR4. J Am Soc Nephrol 2012; 23: 1375–1388.
- 17. Öhlin AK, Larsson K, Hansson M. Soluble thrombomodulin activity and soluble thrombomodulin antigen in plasma. J Thromb Haemost 2005; 3: 976–982.
- 18. Shrestha B, Ito T, Kakuuchi M, Totoki T, Nagasato T, Yamamoto M, Maruyama I. Recombinant thrombomodulin suppresses histone-induced neutrophil extracellular trap formation. Front Immunol 2019; 10: 2535.
- 19. Kato Y, Nishida O, Kuriyama N, Nakamura T, Kawaji T, Onouchi T, Hasegawa D, Shimomura Y. Effects of thrombomodulin in reducing lethality and suppressing neutrophil extracellular trap formation in the lungs and liver in a lipopolysaccharide-induced murine septic shock model. Int J Mol Sci 2021; 22: 4933.
- 20. Hayase N, Doi K, Hiruma T, Matsuura R, Hamasaki Y, Noiri E, Nangaku M, Morimura N. Recombinant thrombomodulin prevents acute lung injury induced by renal ischemia-reperfusion injury. Sci Rep 2020; 10: 289.
- 21. Nozaki Y, Ri J, Sakai K, Niki K, Funauchi M, Matsumura I. Protective effects of recombinant human soluble thrombomodulin on lipopolysaccharide-induced acute kidney injury. Int J Mol Sci 2020; 21: 2519.
- 22. Akatsuka M, Masuda Y, Tatsumi H, Yamakage M. Recombinant human soluble thrombomodulin is associated with attenuation of sepsis-induced renal impairment by inhibition of extracellular histone release. PLoS One 2020; 15: e0228093.
- 23. Kaissling B, Hegyi I, Loffing J, Hir M Le. Morphology of interstitial cells in the healthy kidney. Anat Embryol (Berl) 1996; 193: 303–318.
- 24. Wyatt CM, Crowley SD. A novel role for renal epithelial cells and the medullary sodium gradient in the local immune response. Kidney Int 2017; 92: 1308–1311.
- 25. Ito T, Nakahara M, Masuda Y, Ono S, Yamada S, Ishikura H, Imaizumi H, Kamikokuryo C, Kakihana Y, Maruyama I. Circulating histone H3 levels are increased in septic mice in a neutrophil-dependent manner: Preclinical evaluation of a novel sandwich ELISA for histone H3. J Intensive Care 2018; 6: 79.
- 26. Schindelin J, Arganda-Carreras I, Frise E, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods 2012; 9: 676–682.
- 27. Bitto N, Liguori E, Mura V La. Coagulation, microenvironment and liver fibrosis. Cells 2018; 7: 85.
- 28. Shimomura Y, Suga M, Kuriyama N, Nakamura T, Sakai T, Kato Y, Hara Y, Yamashita C, Nagasaki H, Kaneki M, Nishida O. Recombinant human thrombomodulin inhibits neutrophil extracellular trap formation in vitro. J Intensive Care 2016; 4: 48.
- 29. Kumar V. Pulmonary innate immune response determines the outcome of inflammation during pneumonia and sepsis-associated acute lung injury. Front Immunol 2020; 11: 1722.
- 30. Bogdanos DP, Gao B, Gershwin ME. Liver immunology. Compr Physiol 2013; 3: 567–598.
- 31. Brody JS, Stemmler EJ, DuBois AB. Longitudinal distribution of vascular resistance in the pulmonary arteries, capillaries, and veins. J Clin Invest 1968; 47: 783–799.
- 32. Gaar KA, Taylor AE, Owens LJ, Guyton AC. Pulmonary capillary pressure and filtration coefficient in the isolated perfused lung. Am J Physiol 1967; 213: 910–914.
- 33. Hellems HK, Haynes FW, Dexter L. Pulmonary capillary pressure in man. J Appl Physiol 1949; 2: 24–29.
- 34. Mitzner W. Hepatic outflow resistance, sinusoid pressure, and the vascular waterfall. Am J Physiol 1974; 227: 513–519.
- 35. Laine GA, Hall JT, Laine SH, Granger J. Transsinusoidal fluid dynamics in canine liver during venous hypertension. Circ Res 1979; 45: 317–323.
- 36. Maass-Moreno R, Rothe CF. Nonlinear resistances in hepatic microcirculation. Am J Physiol 1995; 269: H1922–H1930.
- 37. Maass-Moreno R, Rothe CF. Distribution of pressure gradients along hepatic vasculature. Am J Physiol 1997; 272: H2826–H2832.
- 38. Casellas D, Navar LG. In vitro perfusion of juxtamedullary nephrons in rats. Am J Physiol - Ren Fluid Electrolyte Physiol 1984; 246: F349–F358.
- 39. Maddox DA, Deen WM, Brenner BM. Dynamics of glomerular ultrafiltration: VI. Studies in the primate. Kidney Int 1974; 5: 271–278.
- 40. Brenner BM, Troy JL, Daugharty TM, Deen WM, Robertson CR. Dynamics of glomerular ultrafiltration in the rat. II. Plasma-flow dependence of GFR. Am J Physiol 1972; 223: 1184–1190.
- 41. Cowley AW. Long-term control of arterial blood pressure. Physiol Rev 1992; 72: 231–300.
- 42. Nakano D, Doi K, Kitamura H, Kuwabara T, Mori K, Mukoyama M, Nishiyama A. Reduction of tubular flow rate as a mechanism of oliguria in the early phase of endotoxemia revealed by intravital imaging. J Am Soc Nephrol 2015; 26: 3035–3044.
- 43. Shigeoka AA, Holscher TD, King AJ, Hall FW, Kiosses WB, Tobias PS, Mackman N, McKay DB. TLR2 is constitutively expressed within the kidney and participates in ischemic renal injury through both MyD88-dependent and -independent pathways. J Immunol 2007; 178: 6252–6258.
- 44. Nakano D, Kitada K, Wan N, Zhang Y, Wiig H, Wararat K, Yanagita M, Lee S, Jia L, Titze JM, Nishiyama A. Lipopolysaccharide induces filtrate leakage from renal tubular lumina into the interstitial space via a proximal tubular Toll-like receptor 4–dependent pathway and limits sensitivity to fluid therapy in mice. Kidney Int 2020; 97: 904–912.