ORIGINAL ARTICLES
Ionomic Analysis of Horticultural Plants Reveals Tissue-specific Element Accumulation
2015 Volume 84 Issue 4 Pages 305-313

Details
2015 Volume 84 Issue 4 Pages 305-313
Ionomic analysis was performed in three diverse fruit crops in order to characterize element concentrations in edible parts and to compare element concentrations between edible parts and vegetative organs and between species. Nineteen elements, including elements essential for human health, toxic elements, and elements with radioisotopes from nuclear reactors, were analyzed in apple (Malus domestica), Japanese pear (Pyrus pyrifolia), eggplant (Solanum melongena), and edamame (Glycine max). The concentrations of elements were shown as basic information needed for biofortification, and high mobilities depending on plant species were found for some elements whose mobilities are believed to be low. Classification of element profiles into the leaf (or the leaf blade) and other organs containing edible parts, except the calyx, by principal component analysis revealed similarity in the element concentrations across species. The present study may contribute to progress in the ionomic studies of fruit crops.
Ionomic analysis is a type of omics study in which elemental analysis is performed in order to characterize the ionome, the inorganic component of cellular and organismal systems (Salt et al., 2008); the ionomic data of cereals is available in the ionomicsHUB database (http://www.ionomicshub.org/). Ionomic analysis has mainly focused on genetic studies and the effects of soil environment and fertilization on the ionome using vegetative tissues of model plants such as Arabidopsis thaliana and Lotus japonicus (Conn et al., 2012; Quadir et al., 2011). In non-model plant species, the genetic aspects of the ionome and the effects of fertilization and environmental stress on the ionome have been studied in herbage, wildflowers, and beech (Lowry et al., 2012; Peuke and Rennenberg, 2011; White et al., 2012).
Ionomes of edible parts have been reported for rice and soybean (Norton et al., 2010; Sha et al., 2012). In soybean, dried mature seeds are used, and there are no reports of ionome analysis using immature soybean as a vegetable, edamame, as described in the present study. Among horticultural crops, there are reports of ionome analysis in tomato, pear, bilberry, and grape as wine (Elisabetta et al., 2013; Horii et al., 2011; Reuscher et al., 2014; Sánchez-Rodríguez et al., 2010). In tomato, the fruit ionome has not been reported, but the effect of environmental stress on the ionome has been investigated in leaves. In pear and bilberry, although ionomic analysis has been proposed to be useful for the evaluation of fruit development and quality, ionomes have not been compared between fruit and other organs or between different tissues within the fruit. In addition, ionomic analysis using diverse plant species, as in this study, sheds light on the species specificity of ionomes and their similarity among species. This type of study has not been performed in horticultural crops, but it has been performed in herbage (White et al., 2012).
Fruit is important as a source of mineral elements essential for human health as well as vitamins. Deficiencies in some essential elements are common in many developed and developing countries, and biofortification, which increases the concentration and/or bioavailability of elements in food crops, is receiving much attention (Karley and White, 2009; White and Broadley, 2009). Therefore, ionomic studies may contribute to research progress in biofortification as well as in soil science and plant nutrition. Biofortification in fruit crops could be more important for the promotion of human health compared with the use of mineral supplements. Minerals can be absorbed from fruit along with other nutrients, such as vitamins and fibers. For example, the beneficial effect of dietary Ca on colon cancer reportedly depends on the level of dietary fiber (Galas et al., 2013).
Uptake and transport of the radioisotopes of Cs and Sr become a problem in vegetables and fruit trees cultivated in the open field when nuclear fallout from a nuclear blast or nuclear reaction occurs, but information is lacking on within-plant concentrations of Cs and Sr in vegetables and fruit trees. Because apple/pear, eggplant, and edamame are usually cultivated in open fields, they were chosen for analysis. Although radioactive Sr and Cs have been analyzed for characterization of the behavior of their radioisotopes in the environment, including soil and plants, after the nuclear accidents of Chernobyl and Fukushima, the concentrations of non-radioactive Sr and Cs within a plant could be meaningful for an understanding of their behavior after uptake into the plant.
Ionomic studies of edible parts of crops and mineral distribution and movement are now important for the promotion of global health (White and Brown, 2010). Therefore, we performed ionomic analysis in three diverse fruit crops, including fruit trees, fruit vegetables, and leguminous vegetables, grown in the same field, and ionomes were compared between species and between plant organs.
All plants were grown in a field (fine-textured yellow soil; Cultivated Soil Classification Committee, 1995) at the Graduate School of Agricultural Science, Tohoku University, Sendai, Japan, in 2011. Leaves and fruit were sampled from 15-year-old apple trees (Malus domestica ‘Fuji’ and ‘Himekami’ grafted onto M.9/M. prunifolia) and 15-year-old Japanese pear trees (Pyrus pyrifolia ‘Hosui’ and ‘Kosui’ grafted onto P. betulaefolia). They were collected from four trees of each cultivar at the optimum harvesting stage, and the skin and flesh from the fruit and leaves were dried at 60°C for analysis. Eggplants (Solanum melongena ‘Shikon Sendai Naganasu’, ‘Binannasu’, and ‘Shinsen Chunaganasu’ grafted onto S. torvum) and edamame plants (Glycine max ‘Ichibancha’, ‘Sakenotomo Sango’, and ‘Kurogoyou’) were grown in the field with slow-release fertilizer (100 g·m−2; N:P2O5:K2O = 15:15:15; CDU Tamagokasei S555; Co-op Chemical, Tokyo, Japan) and granular dolomite (100 g·m−2; Magutan; Gausukogyo, Tochigi, Japan). Leaves, stems, and fruit were collected from five plants of each cultivar at the optimum harvesting stage. Fruit skin, calyces, fruit flesh, leaves, and stems from eggplants and seeds, pods, leaf blades, petioles, and stems from edamame plants were dried at 60°C for analysis.
AnalysisDried samples were digested with HNO3 and subjected to an elemental analysis by inductively coupled plasma mass spectrometry (ELAN, DRC-e; Perkin-Elmer, Waltham, MA, USA), according to the method of Sha et al. (2012). Five or more samples were collected from each individual and pooled for each individual for extraction. Subsequently, 19 elements were analyzed and the average of four or five individuals per cultivar was calculated.
StatisticsThe calculation of correlation coefficients and principal component analysis (PCA) were performed using Excel (Microsoft, Redmond, WA, USA) and Ekuseru-Toukei 2010 (Social Survey Research Information, Tokyo, Japan). Multiple comparisons by the Tukey-Kramer test were performed using the “Tukey HSD” function of the R statistical software package (ver. 3.0.2, http://www.r-project.org/).
Concentrations of nutritionally important elements in different tissues are shown in Figure 1A. Element concentrations were compared among organs in apple and Japanese pear (apple/pear), eggplant, and edamame. The data from apple and Japanese pear, which are closely related species in the family Rosaceae, are shown in the same figure as apple/pear. The concentrations of elements were similar in apple and Japanese pear.

Concentrations of nutritionally important elements (A), elements with toxicity and radioisotopes (B), and other elements as supplementary data (C) in apple/pear (white), eggplant (gray), and edamame (black). Apple cultivars are ‘Fuji’ ( ) and ‘Himekami’ (
) and ‘Himekami’ ( ) and Japanese pear cultivars are ‘Hosui’ (
) and Japanese pear cultivars are ‘Hosui’ ( ) and ‘Kosui’ (
) and ‘Kosui’ ( ). Eggplant cultivars are ‘Binannasu’ (
). Eggplant cultivars are ‘Binannasu’ ( ), ‘Shinsen Chunaganasu’ (
), ‘Shinsen Chunaganasu’ ( ), and ‘Shikon Sendai Naganau’ (
), and ‘Shikon Sendai Naganau’ ( ) and edamame cultivars are ‘Kurogoyou’ (
) and edamame cultivars are ‘Kurogoyou’ ( ), ‘Ichibancha’ (
), ‘Ichibancha’ ( ), and ‘Sakenotomo Sango’ (
), and ‘Sakenotomo Sango’ ( ). Element concentrations were determined in the fruit flesh (Fls), fruit skin (Skn), leaves (Lf), calyces (Cly), stems (Stm), seeds (Sd), pods (Pd), leaf blades (Bld), and petioles (Ptl). Values are presented as the mean for each cultivar with SE (n = 4 to 5). Values with different letters were significantly different in each of apple/pear, eggplant, and edamame at P < 0.01, according to the Tukey-Kramer test, in A and B.
). Element concentrations were determined in the fruit flesh (Fls), fruit skin (Skn), leaves (Lf), calyces (Cly), stems (Stm), seeds (Sd), pods (Pd), leaf blades (Bld), and petioles (Ptl). Values are presented as the mean for each cultivar with SE (n = 4 to 5). Values with different letters were significantly different in each of apple/pear, eggplant, and edamame at P < 0.01, according to the Tukey-Kramer test, in A and B.

Continued.
Differences in Ca concentrations among organs were large, and edible parts, including the fruit flesh of apple/pear, eggplant, and the seeds of edamame, contained much lower concentrations of Ca than did vegetative organs. Although the calyx is a reproductive organ, Ca concentrations in eggplant calyces were relatively high. In contrast to Ca, differences among organs in K concentration were small, and K concentrations in edible parts were high. The mean K concentrations in all organs were significantly different among apple/pear, eggplant, and edamame at P < 0.01, as determined by the analysis of variance (ANOVA), and the K concentration in eggplant was the highest in every organ among all species. Differences in Ca and K concentrations were not found among cultivars in apple/pear and eggplant, whereas Ca concentrations in the leaf blades and petioles of ‘Ichibancha’ and K concentrations in its petioles differed from those of the other edamame cultivars.
Mg concentrations were species-specific. They were much lower in the edible parts than in the leaves of apple/pear, whereas the differences in Mg concentrations between the edible parts and leaves (or leaf blades) were small in both eggplant and edamame. Similar results were found for Fe, Mn, and Zn (but not Fe in eggplant). The concentrations of these elements were much lower in the fruit flesh of apple/pear than in the leaves, whereas they were not low in the edible parts of eggplant and edamame. Differences in the concentrations of Mg, Zn, and Mn in leaves were found between apple and Japanese pear. Differences in Mg concentrations in edamame leaf blades and petioles, Fe concentrations in eggplant leaves, and Mn concentrations in edamame leaf blades were found among cultivars.
The highest concentrations of Se among all samples were found in the leaves of Japanese pear cultivars. Although the Se concentration differences between the edible parts and vegetative organs in eggplant and edamame were small, the difference in Se concentrations between the fruit flesh and leaves of apple/pear was large.
The concentrations of Cu differed from those of other elements, that is, the concentrations in edible parts were the highest among organs in edamame. Cu concentrations substantially differed between apple and Japanese pear; Cu concentrations in the fruit flesh and skin of the Japanese pear were more than threefold those of apple.
Concentrations of elements with toxicity and radioisotopesThe concentrations of toxic, non-essential elements like Cd and As, and elements present as radioisotopes from nuclear reactors, such as Sr and Cs, are shown in Figure 1B. Cd concentrations were very low compared with concentrations of most other elements in all organs, and a difference was found between apple and Japanese pear; that is, higher Cd concentrations were found in Japanese pear. Differences in Cd concentrations were found among cultivars in edamame seed, leaf blade, and petiole. Concentrations of As were also very low in all organs compared with those of most other elements. Concentrations in edible parts were mostly lower than those in leaves (or leaf blades) and were almost undetectable. A difference in As concentrations was observed between apple and Japanese pear leaves. Differences in As concentrations were also found among edamame cultivars in leaf blades and petioles.
The concentrations of Sr were similar to those of Ca, being low in edible parts. Sr concentrations in ‘Hosui’ leaves were higher than those of the other apple cultivars, and Sr concentrations in ‘Ichibancha’ leaf blades and petioles were higher than those of the other edamame cultivars. Cs concentrations were very low in all organs, similar to those of Cd and As, and concentrations in edible parts were similar to or lower than those in leaves and leaf blades. Cs concentrations in ‘Fuji’ leaves were higher than in those of other Japanese pear cultivars. Differences in Cs concentrations were found among cultivars in eggplant leaves and edamame pods and leaf blades.
Ratios of element concentrations between fruit flesh (or seed) and leaf (or leaf blade) and between fruit flesh (or seed) and skin (or pod)The flesh (or seed) to leaf ratio of important elements could be useful information for biofortification of fruit crops, helping breeders to improve the transport of minerals into the fruit as well as the uptake into plants. The skin (or pod) to flesh (seed) ratios of elements are also useful information for consumers when choosing which parts to eat. The ratios of element concentrations in the fruit flesh (or seeds) to those in the leaves (or leaf blades; i.e., flesh to leaf or seed to leaf ratios) are shown in Figure 2. Concentrations of elements that are not described above are shown in Figure 1C. In apple/pear, the log2 value of the flesh to leaf ratio of Na was approximately 1, and those of K, Cu, Cs, and B were between 0 and −1. The log2 values of the flesh to leaf ratios of other elements were lower than −1, and that of Ca was particularly low. In eggplant, the flesh to leaf ratio of Na was also high, and the log2 values of the flesh to leaf ratios of K, Cu, and B were higher than −1, similar to the case of apple/pear. Those of Zn and P were approximately 0. In edamame, the log2 value of the seed to leaf ratio of Na was lower than 0, unlike in other species, and those of Mo and Cu were uniquely high.
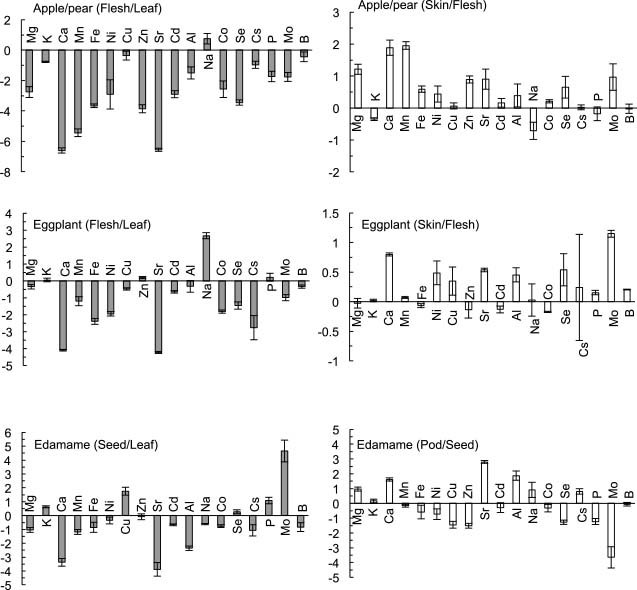
The (log2-transformed) ratios of element concentrations in the fruit flesh (or seeds) to those in leaves (or leaf blades) and the ratios of element concentrations in fruit skin (or pods) to those in fruit flesh (or seeds). Values are presented as mean ± SE (n = 3 to 4) for apple/pear, eggplant, and edamame. The ratios were calculated from element concentrations in each cultivar and averaged. The ratios of As are not shown because of the extremely low concentrations of As in edible parts.
The ratios of element concentrations in fruit skin (or pods) to those in fruit flesh (or seeds; i.e., skin to flesh or pod to seed ratio) are shown in Figure 2. In apple/pear, the log2 values of the skin to flesh ratios of Ca and Mn were higher than around 2, whereas those of other elements were between −1 and 1.5. The log2 values of the skin to flesh ratios of all elements were between −0.5 and 1.5 in the eggplant. The pod to seed ratios in edamame exhibited a wide range: the log2 values of the ratios of Ca, Sr, and Al were higher than 1.5, but those of Cu, Zn, Se, P, and Mo were lower than −1.
Differences in ionome profiles among organsIn order to characterize the differences in ionome profiles among organs across species, the results of 19 elements in different organs of diverse plant species were subjected to PCA (Fig. 3). The cumulative contribution ratio of PCA was 53.4% up to the second principal component (PC2). The first principal component (PC1) was mainly based on the concentrations of Ca, Co, Ni, Sr, and Fe, and their means in all species were significantly different among organs at P < 0.05, according to ANOVA. PC2 was mainly based on the concentrations of K, Al, Na, P, and Mo, and their means in all species were not significantly different among organs at P < 0.05, according to ANOVA. Although each species tended to cluster, the results of PCA indicated that the ionome profiles of these fruit crops could be classified into three groups according to organs: leaf (and leaf blade), calyx, and other organs containing edible parts.

Principal component analysis of element concentrations in various organs from cultivars of apple/pear, eggplant, and edamame. The mean value in each organ of each cultivar was used for the analysis. Ionome profiles were classified into three groups according to organs: leaf and leaf blade of apple (), pear ( ), eggplant (), and edamame (); calyx of eggplant (x); other organs, including fruit flesh, skin, seed, and pod of apple (), pear (
), eggplant (), and edamame (); calyx of eggplant (x); other organs, including fruit flesh, skin, seed, and pod of apple (), pear ( ), eggplant (
), eggplant ( ), and edamame (
), and edamame ( ) as well as stem and petiole of eggplant () and edamame ().
) as well as stem and petiole of eggplant () and edamame ().
Correlation coefficients among element concentrations in all samples are shown in Table 1. The highest correlation was found between Fe and Ni (R = 0.88), and the next highest correlation was between Ca and Sr (R = 0.87). Correlations were also relatively high (R > 0.7) between K and Al, Ca and Fe, and Ca and Ni. Some combinations of elements showed significant correlations (0.57 < R < 0.7). Negative correlations were found among fewer combinations of elements.

Correlation coefficients between elements.
High (> 0.8) correlation coefficients were found between Ca and Sr and between Fe and Ni, and both pairs consist of chemically similar elements. Correlation coefficients of approximately 0.7 were also found between chemically similar elements such as K and Na as well as Mn and Zn. Concentrations of these chemically similar elements are likely correlated because they share pathways and those pathways are conserved across species.
A positive correlation was also found between the chemically similar elements K and Na, but this correlation was not significant in previous studies (Baxter et al., 2012; Sha et al., 2012). Given that this correlation was previously analyzed on the basis of variation under different cultivation conditions, the absence of significance could have been due to a difference in the response to such cultivation conditions.
Although the mobility of Fe, Zn, and Cu in the phloem is generally considered to be low, similar to that of Ca (White and Broadley, 2009), the differences in the concentrations of these elements between edible parts and vegetative organs were found to be small for Fe in edamame, Zn in eggplant and edamame, and Cu in all species. The result of Cu in apple/pear is supported by a previous study that examined apple fruit and leaves (Johnson, 2000). The phloem mobility of Zn in eggplant and edamame described here may be controlled by the proposed mechanism of increased mobility, which involves the expression of proteins responsible for Zn mobilization (White and Broadley, 2009). Moreover, active Zn transport to seeds has been shown in tracer experiments involving maturing soybean (Shinonaga et al., 1998). The seed to leaf ratio of Cu in edamame (log2 = 1.76) was much higher than that in dried mature soybean (log2 ≈ 0; Duke et al., 2012), suggesting the existence of an active Cu accumulation mechanism in immature seeds. Because Cu mobility could be enhanced by a complex of Cu with small proteins or nicotinamine (White and Broadley, 2009), the relationship between this mechanism and Cu concentrations in edible parts should be investigated further.
Se is proposed to be transported as selenate and/or organoselenium compounds in the phloem (White and Broadley, 2009) and seemed to be phloem-mobile in edamame and eggplant in this study. However, the flesh to leaf ratio of Se was very low (log2 = −3.45) in apple/pear, suggesting that Se mobility in the phloem is species-dependent. Apple Se concentration is known to be comparatively high among vegetables and fruit (Diaz-Alarcon et al., 1994), and Se concentration in apple/pear leaves was the highest among all samples in this study. Thus, biofortification of Se in apple/pear could be increased by an increase in mobility within the plant.
Mo concentration was unique in edamame, with a seed to leaf ratio of 4.66 (as a log2 value), suggesting the existence of an edamame-specific mechanism of Mo accumulation in seeds. This suggestion is supported by a previous study of mineral redistribution regulation using soybean explants (Mauk and Noodén, 1992). In another soybean study, Mn and Fe interactions were reported (Heenan and Campbell, 1983), and our study also found a high correlation between these two elements in edamame (R = 0.85, P < 0.001), although a strong correlation was not found in such a comparison in all species (Table 1).
Concentrations of As, a toxic element, differed substantially between organs, similar to Ca, and edible parts contained very low or undetectable levels of As. The result in apple is supported by a previous study (Peryea, 1998). Low concentrations of As in fruit or seeds are desirable in food crops. Cd concentrations were also low, and differences among organs were small, except in Japanese pear. The result in eggplant is also supported by a previous study (Qadir et al., 2000). Cd concentrations in Japanese pear were higher than those in all apple tissues, suggesting that apple and Japanese pear have different mechanisms for Cd uptake and transport despite being closely related species. Cd concentrations in edible parts were lower than the limit of 0.4 mg·kg−1 for rice in Japan (Ministry of Health, Labour, and Welfare).
The concentrations of Sr were similar to those of Ca, which is a chemically similar element, and the correlation coefficient between these two elements was high (Table 1). Cs concentrations showed relatively small differences among organs, suggesting that Cs is phloem-mobile like K, which is a chemically similar element. A similarity in Ca and Sr concentrations was also found in the comparison among edamame cultivars; that is, the concentrations of both elements were higher in ‘Ichibancha’ than in the other cultivars in leaf blades and petioles, suggesting that Ca and Sr are distributed via the same pathways. On the other hand, the similarity between K and Cs concentrations was not found in the comparison of cultivars in each species. To the best of our knowledge, there are no reports on the concentrations of Sr and Cs in various organs from Japanese pear, eggplant, and edamame. Although soil–plant transfer factors of radioactive Cs and Sr have been reported in apple (Green et al., 1997), the concentrations of these elements among different tissues within plants have not yet been reported. Our results identify the parts in which these elements accumulate.
The flesh (or seed) to leaf ratios of most elements were less than 0 (as a log2 value), suggesting that elements preferentially accumulate in leaves and that the ratios depend on their mobility. In fact, Ca, Mn, and Zn, of which the log2 values of the flesh to leaf ratios were very low, accumulate in leaves during fruit development in Japanese pear, while the other elements decrease (Buwalda and Meekings, 1990). As a result, although both Ca and K were the most abundant elements in leaves, only K concentration was predominantly high in fruit. Leaf K concentrations are consistent indicators of fruit K in apple (Casero et al., 2004), and our apple/pear data supported this (R = 0.97). Increasing the flesh (or seed) to leaf ratios of essential elements as well as their uptake is desirable for the biofortification of fruit crops. In particular, the fruit to leaf ratios of Ca, Mn, and Zn in apple/pear were much lower than those in eggplant and edamame. This low ratio of Ca (log2 = −6.6) in apple is supported by a previous study (log2 = −5.5; Vang-Petersen, 1980). Because concentrations in leaves did not substantially differ among species, improving their mobility within the plant should be the focus of biofortification of these elements in apple/pear.
The mobility of B is generally believed to be low because of its cell wall-bound form; however, its flesh to leaf ratio was comparatively high in apple/pear and eggplant. This high ratio in apple/pear could be related to the increase of B mobility by sorbitol, which is one of the main transport forms of photosynthate in apple/pear (Kanayama, 2009). In contrast, a novel mechanism of B phloem mobility that is not dependent on sorbitol may be present in eggplant, whose main transport form of photosynthate is sucrose.
The skin to flesh ratios of elements are important information, helping consumers to choose whether to remove the skin before consumption; however, there have been only a few reports on these ratios. Although pods of edamame are not edible generally, pod to seed ratios were estimated for reference. The skin (or pod) to flesh (or seed) ratios were mostly higher than the flesh (or seed) to leaf ratios, and those of most elements were between 0 and 1 (as a log2 value). The log2 values of ratios of around 0 appear reasonable, given that the fruit flesh and skin as well as seeds and pods are similar organs. In contrast, the skin (or pod) to flesh (or seed) ratios were found to be around 2 or higher in Ca and Mn in apple/pear and Sr and Al in edamame, suggesting the presence of skin (or pod)-specific accumulation mechanisms for these elements. The concentrations of Mg, K, Ca, Mn, Fe, Cu, and Zn in apple flesh and the skin to flesh ratios of apple/pear were roughly the same as those of a previous study (Gorinstein et al., 2001). The concentrations of the elements in Japanese pear flesh were also roughly the same as those of a previous study (Buwalda and Meekings, 1990). These similarities suggest that their concentrations are relatively constant. As an exception, the Mn concentration in Japanese pear fruit observed by Buwalda and Meekings (1990) was approximately 3 times higher than that observed in this study, whereas the flesh (fruit) to leaf ratios were similar. Therefore, although Mn concentrations depend on environmental conditions, the ratios are most likely to be relatively constant.
While the concentrations of Mg, K, Fe, Cu, and Zn in eggplant were roughly the same as those of a previous study (Arivalagan et al., 2012), the Cd concentration was one order of magnitude lower than that observed in another previous study (Demirezen and Aksoy, 2006). While the concentrations of 10 essential elements in edamame were also roughly equivalent to those observed in a previous study, Se concentrations in it were much lower than the average value in that study (Wolnik et al., 1983). These patterns suggest that the concentrations of non-essential elements, such as Cd and Se, are variable, while those of essential elements are relatively constant.
Classification of element profiles into leaf (and leaf blade) and other organs containing edible parts, except for the calyx, in PCA revealed similarity in element concentration across species, including different families, true and false fruit, and herbaceous and woody crops. Furthermore, the concentrations of elements that are important as dietary minerals are basic data needed for future biofortification, and high, species-dependent mobilities were found in some elements whose phloem mobilities are believed to be low. All findings obtained in this study reveal the utility of ionomic analysis. Its utility in fruit crops has been recently shown by a comparison between bilberry fruit in different habitats as well as a comparison between mutant and wild-type pear fruit (Elisabetta et al., 2013; Reuscher et al., 2014). Although future ionomic analysis should be performed in crops grown under different conditions as well as in other fruit crops, the present study may broadly contribute to progress in the ionomic studies of fruit crops.
We thank the members of the laboratory of Horticultural Science at Tohoku University for help in plant cultivation and sample preparation.