ORIGINAL ARTICLES
Effects of Interrupted Lighting on the Spray Formation of Summer-to-autumn-flowering Small-flowered Spray-type Chrysanthemum Cultivars ‘Haruka’ and ‘Subaru’
2016 Volume 85 Issue 3 Pages 264-271


Details
2016 Volume 85 Issue 3 Pages 264-271
Summer-to-autumn-flowering small-flowered spray-type (SAFS) chrysanthemums (Chrysanthemum morifolium Ramat.) are commercially important in Japan, and there is an occasional increased demand for these flowers. However, under current cultivation methods, the quality of spray formations is compromised in order to maintain a sufficient production rate at times of peak demand. This study investigated the effect of interrupted lighting (IL), the intercalation of a period of long days (by night interruption) into the natural day length (NDL) period of growing, in order to regulate spray formations in SAFS chrysanthemum cultivars ‘Haruka’ and ‘Subaru’, in which lighting can markedly delay flowering. The effects of the NDL period before IL and those of the IL period were investigated. First, we investigated the effect of 2–12 days of NDL followed by 12 days of IL. Plants subjected to 2–6 days of NDL had longer flower clusters and a greater number of flower buds on the upper lateral flower stems and developed a broom-like spray. Next, we investigated the effect of 4–20 days of IL after 4 days of NDL. Plants subjected to 8–20 days of IL had longer flower clusters, more flower buds on the upper lateral flower stems, and broom-like sprays. These changes were more pronounced when the IL periods were prolonged. Our results showed that spray formation in SAFS chrysanthemum cultivars ‘Haruka’ and ‘Subaru’ can be regulated by controlling the timing and period of IL.
In Japan, small-flowered spray-type chrysanthemums are used in many religious festivals, and hence, their demand increases at particular times of the year. About 40% of these flowers are marketed from July to September. During periods of high demand, summer-to-autumn-flowering small-flowered spray-type (SAFS) chrysanthemums are used; these are usually grown under natural day length (NDL). Since the flowering time of SAFS chrysanthemums is strongly influenced by climatic conditions, maintaining their production to meet peak demand is difficult. Thus, we investigated a light culture using 9 small-flowered spray-type chrysanthemum cultivars that flower from June to July under NDL; we found that flowering could be controlled in 4 cultivars (‘Haruka’, ‘Subaru’, ‘Seikomaki’, and ‘Seichigusa’) by modifying lighting regimens, allowing growers to meet the peak demand supply from July to September (Mori et al., 2014). However, in these 4 cultivars, the terminal capitulum tended to be set above the lateral capitula, and spray formation was different from that of the traditional SAFS chrysanthemum cultivars (the terminal capitulum is set under the lateral capitula, making the spray formation appear broom-like). Moreover, in ‘Haruka’, only few capitula were present on the upper lateral flower stem under both light and no-light culture conditions; ‘Subaru’ and ‘Seikomaki’ occasionally have only a few capitula on the upper lateral flower stem under light culture. Therefore, some customers want growers to regulate spray formation produced using light culture. However, the demand for different spray formations differs across customers; at present, growers change the cultivar type to meet the demands of customers. If growers can regulate spray formation, the demands for various spray formations can be met by using the same cultivar.
Osuga et al. (1978) showed that standard-type chrysanthemums developed small upper leaves under light culture, and that these could be restored to their normal length by using interrupted lighting (IL). IL is the intercalation of a period of long days (LD) by night interruption (NI, a short exposure to light during the middle of the night) into the short day (SD) phase of growing. This method has since been used widely in Japan to improve the quality of standard-type chrysanthemum cut flowers.
In spray-type chrysanthemums, IL has been shown to regulate the cut flower characteristics (Naka and Maeda, 2006; Takahashi and Nakano, 1999; Tjia and Glasser, 1976; Yamagata et al., 2008). In particular, IL has been investigated in winter- and autumn-flowering small-flowered spray-type chrysanthemums (Sato, 1981; Yamagata et al., 2008); however, few studies have determined its effectiveness in summer-to-autumn-flowering types. In the autumn-flowering small-flowered spray-type chrysanthemum cultivar ‘Tsubasa’ cultivated using IL, the spray formation of plants became broom-like because the length of lateral flower stems increased and axillary buds developed on lateral flower stems (Yamagata et al., 2008). This indicates that IL could also regulate spray formation in SAFS chrysanthemums. However, a previous study showed that the effect of IL on spray formation varied markedly according to its timing (Yamagata et al., 2008). This study aimed to investigate the effects of the timing and period of IL on spray formation in two SAFS cultivars ‘Haruka’ and ‘Subaru’.
‘Haruka’ and ‘Subaru’ were grown in an open field at the Okayama Prefectural Technology Center for Agriculture, Forestry and Fisheries (Akaiwa, Okayama) in 2013 and 2014. In both years, 6 rooted cuttings were transplanted on May 14 into a plastic container (width, 23 cm × length, 64 cm × height, 18.5 cm) filled with a mixture of silty loam, perlite, and peat moss (1:1:1 volume ratio). To all the plants, 900 mg N, 705 mg P2O5, and 835 mg K2O of slow-release fertilizer (Ecolong 413-100; Jcam Agri. Co., Ltd., Tokyo, Japan) was applied. After 10 days, plants were pinched at the uppermost leaf, and each plant was allowed to grow three shoots. Plants were grown under NI conditions, i.e., NDL plus a daily 4 h (22:00–2:00, 2013) or 6 h (22:00–4:00, 2014) of NI with incandescent bulbs (K-RD110V75W; Panasonic Corp., Osaka, Japan), to maintain the plants in a vegetative state until the treatments were started. Incandescent bulbs were set at every 4 m2, at a height of 1.5 m from the soil surface of the container, since horizontal illuminance at the soil surface became higher than 50 lx.
Experiment 1: Effect of timing of IL on flowering and cut flower characteristicsAll plants were transferred to NDL treatment on June 20, 2013. SAFS cultivars have longer critical day length for flowering than autumn-flowering cultivars (Oda et al., 2010; Sumitomo et al., 2013, 2014); therefore, NDL induced flowering in the 2 cultivars. Plants were transferred to NI (22:00–2:00) at intervals of 2, 4, 6, 8, or 12 days, and then subjected to NI for 12 days after NDL treatment (IL treatments, Fig. 1). Subsequently, the plants were transferred back to NDL until flowering. Control plants were not subjected to IL. Six plants were used for each treatment.

Interrupted lighting (IL) treatments in Experiments 1 and 2. White and grey horizontal bars represent days of natural day length (NDL) with or without night interruption, respectively.
At the beginning of each IL treatment, 1 shoot apex of the second lateral shoot was collected from each plant and dissected under a stereoscopic microscope to observe the stage of flower development. For the remaining plants, the date of flowering was recorded, and this was defined as the appearance of a visible disk floret on the terminal capitulum. At flowering, each shoot was collected, and the following characteristics of the cut flowers were recorded (Fig. 2): cut flower length, stem length, flower cluster length, neck length of the terminal capitulum, number of primary lateral flower stems, total number of capitula per cut flower, flower cluster width, number of capitula on each lateral flower stem (1st to 10th lateral flower stems down from the terminal capitulum), and length of each lateral flower stem (3rd, 6th, and 9th lateral flower stems). Chrysanthemums have a condensed indeterminate inflorescence (capitulum) structure composed of many individual florets, which bears a single flower-like architecture. Thus, the capitulum of chrysanthemums apparently mimics a large single flower. Therefore, in this report, we refer to the mass of capitula as a “flower cluster” (Fig. 2).
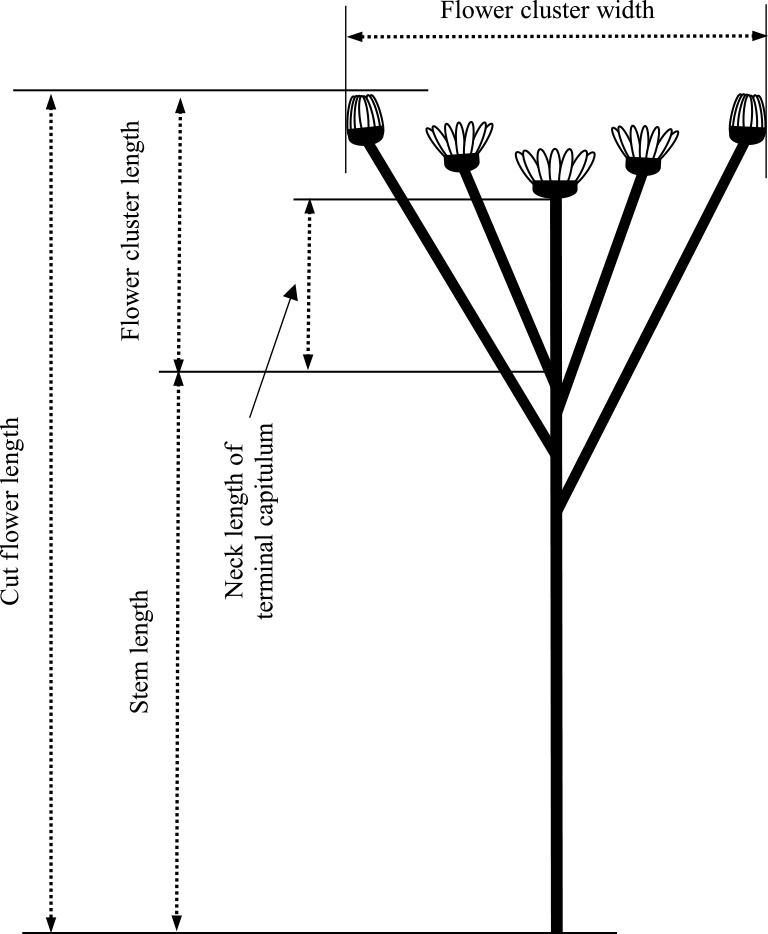
Measurements of cut flower characteristics.
From June 20, 2014, plants were grown for 4 days under NDL. Plants were then transferred to NI (22:00–2:00) on June 24, and grown for 4, 8, 12, or 20 days (IL treatments, Fig. 1). Control plants were not subjected to IL. After IL, plants were returned to NDL. Twelve plants were used in each treatment. The stage of flower development, flowering date, and cut flower characteristics (including leaf number besides those described above) were measured.
Average temperature from the date of first transfer to NDL (June 20) to the final end date of IL (July 14) was 25.2°C at the Okayama Prefectural Agriculture, Forestry and Fisheries Research Center. There were no morphological changes (such as flower initiation) at the shoot apex until 4 days after transfer to NDL (Fig. 3). At 6, 8, and 12 days after transfer to NDL, the shoot apex reached the dome-formation stage, and the first and latter stages of involucre primordia differentiation, respectively. Flower development at 12 days after transfer to NDL was slightly earlier in ‘Haruka’ than in ‘Subaru’, but not significantly (P = 0.061).

The effect of natural day length (NDL) period on flower development of summer-to-autumn-flowering small-flowered spray-type chrysanthemums. Flower development stages are (0) vegetative growth, (1) dome formation, (2) first stage of involucre primordia differentiation, and (3) latter stage of involucre primordia differentiation. One dot indicates one shoot. There was no significant difference between the 2 cultivars at P < 0.05 by using Fisher’s exact test.
IL after 2–12 days of NDL significantly delayed flowering in both the cultivars compared with that in the controls (Table 1). Flowering occurred latest when subjected to IL after 2 and 4 days of NDL in the ‘Haruka’ and ‘Subaru’ plants, respectively. With a decrease in NDL period, cut flower length and stem length increased. Compared with those of controls, cut flower length and stem length were significantly greater when subjected to IL after 2–6 and 2 days of NDL, respectively, in ‘Haruka’, and after 2–8 and 2–4 days of NDL, respectively, in ‘Subaru’. Flower clusters were significantly longer when subjected to IL after 2–4 days of NDL in ‘Haruka’ and 4–12 days of NDL in ‘Subaru’, with the longest clusters formed in plants subjected to IL after 4 and 6 days of NDL treatment, respectively. Thus, earlier application of IL resulted in a strong plant growth stimulus. However, the effect varied among the 2 cultivars. The neck length of the terminal capitulum was significantly greater with IL after 4 days of NDL in ‘Haruka’ and with IL after 6–12 days of NDL in ‘Subaru’. In both cultivars, the number of primary lateral flower stems, total number of capitula per cut flower, and flower cluster width did not differ from those of the controls.

Effects of natural day length (NDL) period before 12 days of interrupted lighting (IL) on flowering and cut flower characteristics of summer-to-autumn-flowering small-flowered spray-type chrysanthemums.
The effect of IL on the number of capitula on each lateral flower stem varied among the cultivars. The number of capitula on the 1st to 7th lateral flower stems increased with IL after 2–4 days of NDL in ‘Haruka’ (Fig. 4). In ‘Subaru’, IL after 2 days of NDL had no effect on the number of capitula, whereas IL after 4–12 days of NDL increased the number of capitula. The lateral flower stems of which the capitula number increased with IL appeared at a lower position with the delay in the timing of IL. IL after 4, 6, 8, and 12 days of NDL increased the number of capitula on the 1st to 10th stems, 3rd to 10th stems, 6th to 10th, and 10th stems, respectively. The number of capitula on the upper 5 flower stems did not increase in either cultivar with IL after 8–12 days of NDL. Late IL had no effect on the upper lateral flower stems of the plants. This trend was prominent in ‘Haruka’. The number of capitula on the 1st to 7th lateral flower stems increased with IL after 2–4 days of NDL, but not after 6–12 days of NDL. Thus, the IL effect on the number of capitula on each lateral flower stem varied markedly depending on the IL timing and cultivar type.

The effect of natural day length period before 12 days of interrupted lighting (IL) on the number of capitula on each lateral flower stem of summer-to-autumn-flowering small-flowered spray-type chrysanthemums. Vertical bars represent SE (n = 10–12).
‘Haruka’ showed longer lateral stems than those of the controls with IL after 4 days of NDL for the 3rd, 6th, and 9th stems, after 6 days for the 3rd and 6th stems, and after 8 days for the 6th stem (Fig. 5). In ‘Subaru’, the 3rd and 6th lateral flower stems were longer with IL after 4 and 12 days of NDL, and the 3rd, 6th, and 9th stems were longer after 6 and 8 days of NDL. The spray formation in the 2- and 4-day NDL treatments in ‘Haruka’ and the 4- to 6-day treatments in ‘Subaru’ differed substantially from that in the control, i.e., those in the treated plants became broom-like with many flower buds over the terminal capitulum (Fig. 6).

The effect of natural day length period before 12 days of interrupted lighting (IL) on the length of each lateral flower stem of summer-to-autumn-flowering small-flowered spray-type chrysanthemums. Vertical bars represent SE (n = 10–12).

The effect of natural day length (NDL) period before 12 days of interrupted lighting (IL) on spray formation of summer-to-autumn-flowering small-flowered spray-type chrysanthemums.
Average temperature from the date of first transfer to NDL (June 20) to the final end date of IL (July 14) was 24.3°C at the Okayama Prefectural Agriculture, Forestry and Fisheries Research Center. There were no morphological changes (such as flower initiation) at the shoot apex at 4 days after transfer to NDL. Eight to 20 days of IL showed quantitative effects on cut flower characteristics, although 4 days of IL had little effect (Table 2). IL increased the days to flowering, cut flower length, stem length, flower cluster length, neck length of terminal capitulum, total number of capitula per cut flower, and flower cluster width. Further, the number of capitula on each lateral flower stem (Fig. 7) and the length of each lateral flower stem (Fig. 8) increased. Plants showed a simultaneous increase in these parameters with IL period. The number of leaves and primary lateral flower stems showed no consistent trend in their response to IL.

Effects of interrupted lighting (IL) periods after 4 days of natural day length (NDL) on flowering and cut flower characteristics of summer-to-autumn-flowering small-flowered spray-type chrysanthemums.
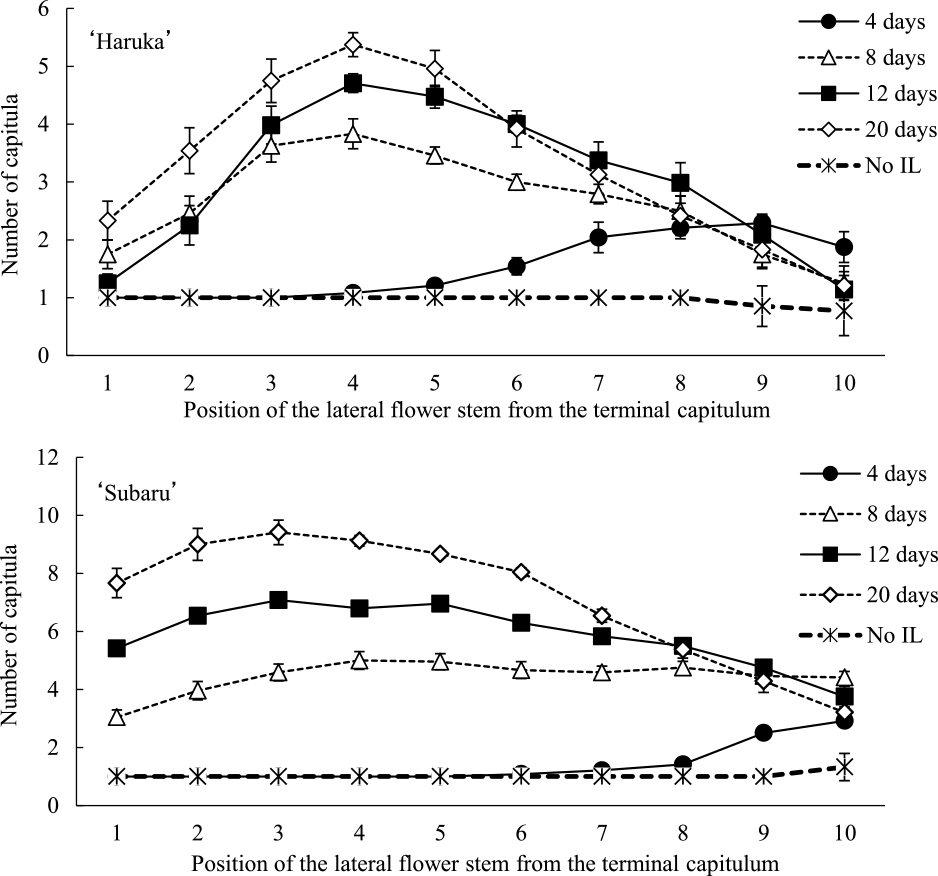
The effects of interrupted lighting (IL) periods after 4 days of natural day length on the number of capitula on each lateral flower stem of summer-to-autumn-flowering small-flowered spray-type chrysanthemums. Vertical bars represent SE (n = 23–24).

The effects of interrupted lighting (IL) periods after 4 days of natural day length on the length of each lateral flower stem of summer-to-autumn-flowering small-flowered spray-type chrysanthemums. Vertical bars represent SE (n = 23–24).
In this study, we showed that IL could substantially regulate the appearance of the cultivated SAFS chrysanthemum cultivars ‘Haruka’ and ‘Subaru’. The findings of our first experiment suggest that the NDL period before IL is important for flower development and spray formation in both cultivars. The spray formation is composed of the length of lateral flower stems and number of capitula on the upper lateral flower stem. IL affected both these parameters, and thus changed the spray formation. With IL after 2 or 4 days of NDL, ‘Haruka’ plants had longer flower clusters and more flower buds on the upper 5 lateral flower stems, leading to a broom-like spray. In ‘Subaru’ plants, these changes occurred after longer NDL periods of 4–6 days. This suggests that spray formation in SAFS chrysanthemums can be regulated by IL; however, the NDL period before IL markedly affected spray formation, as has been previously shown in autumn-flowering small-flowered spray-type chrysanthemums (Yamagata et al., 2008), and the appropriate NDL period to obtain the broom-like spray formation is different across cultivars. In pompom chrysanthemums, Post (1943) showed that long days (LD) for a period of 5–20 days after short days (SD) for 7–35 days increased the peduncle length of the upper lateral flower stems in most combinations of day length, and that 7 days of SD had the greatest effect. Consistent with our study results, their findings showed that SD period before LD could substantially influence flower shape.
The change in spray formation is likely to occur due to the effects of IL on the timing of different developmental stages. First, the timing of flower bud initiation was affected, with IL inhibiting the development of axillary flower buds near the apical flower buds. Subsequently, secondary lateral flower stems developed with the restoration of vegetative growth of the primary lateral flower stems. Next, with an increase in the length of lateral flower stems and flower clusters, more flower buds developed on the secondary lateral flower stems. Thus, spray formation changed with the increase in the number of flower buds on the upper lateral flower stems. This effect was less marked when IL was started after very little or excessive NDL. Presumably, in the former case, flower bud initiation had not yet started, whereas in the latter, flower bud development had markedly progressed for axillary bud development to be sufficiently suppressed. Thus, the timing of the beginning of IL may be very important.
In our experiment, the appropriate NDL period to obtain the broom-like spray formation was different between the 2 cultivars, i.e., 2–4 days in ‘Haruka’ and 4–6 days in ‘Subaru’ (Fig. 6). This difference may be attributed to the timing of flower initiation: flower development at 12 days after transfer to NDL in ‘Haruka’ was slightly earlier than that in ‘Subaru’; however, the difference was not significant (P = 0.061; Fig. 3). The most effective time to start IL was between the vegetative stage and dome formation on terminal buds. Thus, during commercial production, flower development needs to be uniformly controlled across many plants to maximize the profit obtained from regulating spray formation by using IL. However, under NI conditions, undesirable premature flower initiation is often observed in SAFS chrysanthemums (Mori et al., 2014). Thus, selected SAFS cultivars, which stay vegetative under non-inductive photoperiodic conditions, and show synchronous flower initiation in response to transfer from non-inductive to inductive photoperiodic conditions such as ‘Haruka’ and ‘Subaru’, should be used for stable regulation of spray formation using IL. Mori et al. (2014) suggested that ‘Seikomaki’ and ‘Seichigusa’ could be used for light culture as well as ‘Haruka’ and ‘Subaru’. Therefore, the regulation of spray formation in ‘Seikomaki’ and ‘Seichigusa’ may also be possible. However, the rate of flower development is known to be affected by environmental factors even in these SAFS chrysanthemums (Koyama and Wada, 2004; Mori et al., 2014). Therefore, monitoring the status of flower development is required to determine the effective IL timing, although microscopic observation of flower bud development may not be sufficient to decide when to start IL, because such observation may not indicate the time when flower bud initiation began. Therefore, an index needs to be developed that can be used to determine the exact time to start IL.
Our second experiment investigated the effects of IL period length. Both the cultivars developed longer flower clusters and more flower buds on the upper 5 lateral flower stems after 8–20 days of IL, transforming the spray to a broom-like one, with prominent changes noted after longer IL periods. Thus, quantitatively controlling spray formation in SAFS chrysanthemum cultivars ‘Haruka’ and ‘Subaru’ by using IL and producing various spray formations by changing the IL period may be possible. Indeed, previous studies have shown that 8–14 days of IL after 20 days of SD could increase cut flower length and the number of flower buds, with longer IL periods causing greater changes in winter-flowering small-flowered spray-type chrysanthemums (Sato, 1981). This is consistent with our findings suggesting that IL period affects the number of flower buds.
Our study results suggest that regulating spray formation in SAFS chrysanthemum cultivars ‘Haruka’ and ‘Subaru’ is possible by using IL. We found that 2–6 days of NDL followed by 8–20 days of IL was suitable; however, these periods varied according to environmental factors and responses of cultivars. Therefore, determining the suitable NDL and IL periods for each cultivar and each cropping type are necessary. Furthermore, we found that quantitatively controlling spray formation in ‘Haruka’ and ‘Subaru’ by changing the IL period may be possible. Because different customers prefer different types of spray formations, determining suitable IL periods for the different types of spray formations to meet the demands of customers is important. Moreover, because IL delayed flowering, NI should be stopped earlier in consideration of the delayed flowering period. Further studies are warranted to investigate a range of cultivation methods combined with IL. These techniques could facilitate the production of various spray formations by using the same cultivar, thereby allowing growers to meet various demands by using only a few cultivars. This would stabilize the management of cut flower production. Introduction of these techniques mean that growing areas will need an electricity supply and lighting equipment; nonetheless, this will not be a challenge for growers already using light culture.