Abstract
The flowering response to monochromatic light and flowering-related genes underlying this response need to be characterized to efficiently use light-emitting diodes for lighting culture. The flowering response to far-red light has been well studied in long-day cut flowers, but there have been few studies investigating the response to blue light. Flowering and the expression of the G. paniculata homologs of the FLOWERING LOCUS T and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 genes (GpFT, GpSOC1) were not previously promoted in Gypsophila paniculata, an important long-day cut flower, ‘Bristol Fairy’ under long-day conditions with blue light. In the present study, we found that flowering was promoted in another G. paniculata ‘Million Star’, under long-day conditions with blue light, suggesting that there is variation in G. paniculata’s flowering response to blue light. Therefore, we analyzed the expression of GpFT and GpSOC1 in the ‘Million Star’. The expression of GpFT and GpSOC1 was promoted with flowering in ‘Million Star’ under long-day conditions with blue light in contrast to ‘Bristol Fairy’. We next analyzed the G. paniculata homologs (GpFKF1, GpGI) of FLAVIN-BINDING KELCH REPEAT F-BOX 1 and GIGANTEA genes, which participate in the flowering response to blue light. GpFKF1 and GpGI amino acid sequences were well conserved; gene expression showed a diurnal rhythm with different peaks under short-day and long-day conditions, as previously observed in Arabidopsis thaliana. GpFKF1 interacted with GpGI. There were no important differences in GpFKF1 or GpGI amino acid sequences between the two cultivars. Our results suggest that variation in the flowering response to blue light is associated with GpFT and GpSOC1, rather than GpFKF1 and GpGI.
Introduction
Production of long-day cut flowers during autumn and winter is possible by long-day treatment to promote flowering. Although incandescent lamps have been used as a light source for long-day treatment, they have some disadvantages including high-energy consumption and short life. In contrast, light-emitting diodes (LEDs) are expected to reduce the environmental burden, owing to their low energy consumption and long life. Besides, because LEDs emit monochromatic light, lighting at the most effective wavelength can be used for long-day treatment. For these reasons, LEDs are receiving much attention as a new light source for long-day treatment.
The effects of monochromatic light quality on plant development should be investigated for the efficient use of LEDs in long-day treatment. Blue, red, and far-red lights are effective for plant flowering (Eskins, 1992), and their molecular mechanisms have been studied mainly in the model plant Arabidopsis thaliana (Andrés and Coupland, 2012; Shibuya and Kanayama, 2014).
FLOWERING LOCUS T (FT), which is known as florigen, with its main transcription factor CONSTANS (CO) plays a central role in the promotion of flowering in A. thaliana under long-day conditions (Corbesier et al., 2007; Wigge et al., 2005; Yoo et al., 2005). Recent studies have also shown that FT plays a key role in flowering in various horticultural plants (Nakajima et al., 2014; Xing et al., 2014). One of the blue light receptors regulating CO accumulation is CRYPTOCHROME 2 (CRY2) and the cry2 mutant shows late flowering under long-day conditions in A. thaliana (Guo et al., 1998). The other is FLAVIN-BINDING KELCH REPEAT F-BOX 1 (FKF1). FKF1 forms a complex with GIGANTEA (GI) in a blue-light-dependent manner and induces the degradation of CYCLING DOF FACTOR (CDF) 1 and its family proteins (CDFs), which are repressors of CO expression (Fornara et al., 2009; Imaizumi et al., 2003, 2005; Sawa et al., 2007). FKF1 also interacts with CO and stabilizes CO, and blue light enhances this interaction (Song et al., 2012). Furthermore, FKF1 removes CDF1, which also represses FT transcription, from the FT promoter (Song et al., 2012). FKF1 and GI thus play important roles in flowering induced by blue light. However, there are few studies on FKF1 and GI in long-day cut flowers.
Promotion of flowering by far-red or a low red:far-red ratio has been relatively well studied in terms of the flowering response of long-day cut flowers to light quality. For example, a low red:far-red ratio maintained using films that change light quality in daytime and red- or far-red-rich fluorescent lamps at night promote flowering in Matthiola incana (Yoshimura et al., 2002, 2006), and low and high red:far-red ratios promote and retard flowering under long-day conditions in Eustoma grandiflorum (Sato et al., 2009; Yamada et al., 2008a, b, 2009). Far-red alone and a low red:far-red ratio promote flowering under long-day conditions with LEDs in Gypsophila paniculata (Nishidate et al., 2012), and a low red:far-red ratio also promotes flowering in Campanula carpatica (Kristinsen, 1988). However, there are few studies on the flowering response to blue light in long-day cut flowers. Blue LEDs, in addition to far-red LEDs, represent a potential light source for lighting culture because blue light promotes flowering in the model plant A. thaliana (Eskins, 1992). Blue LEDs are also used with red LEDs in plant factories. Blue LED use has become increasingly common and widespread, and blue LEDs are inexpensive and also easier to obtain than far-red LEDs, which are not common. Using blue LEDs can thus help mitigate the costs associated with the use of LEDs to promote flowering.
For the efficient use of blue LEDs in lighting culture of long-day cut flowers, the flowering response to blue light and flowering-related genes underlying this response need to be characterized. We used Baby’s Breath (G. paniculata) in this study because it is often used in flower arrangements and is an important long-day cut flower (Kikuchi et al., 2000; Shillo and Halevy, 1982). G. paniculata is grown by lighting culture for year-round production and shows a clear flowering response to long-day treatment as a qualitative long-day plant (Hori et al., 2011; Nishidate et al., 2012). In a previous study (Hori et al., 2011), flowering and the expression of the G. paniculata homologs of the FT and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 genes (GpFT, GpSOC1) were not promoted in G. paniculata ‘Bristol Fairy’ under long-day conditions with blue light. In the present study, flowering was promoted in another G. paniculata ‘Million Star’, under long-day conditions with blue light, suggesting that there is variation in G. paniculata’s flowering response to blue light. We accordingly analyzed flowering-related genes to further investigate this variation.
Materials and Methods
Plant materials
G. paniculata ‘Million Star’ (Sumika Agrotech, Osaka, Japan) and ‘Bristol Fairy’ (Japan Agribio Company, Tokyo, Japan) were grown under short-day conditions with an 8-h photoperiod for approximately 4 weeks. Thereafter, 16-h light treatment was started following the method of Nishidate et al. (2012). Three lateral shoots of ‘Million Star’ plants with approximately 10 leaves were grown in a growth chamber at 22/18°C under natural light from 9:00 to 17:00 and under far-red or blue LEDs from 17:00 to 9:00 for observation of flowering, expression analysis of GpFT and GpSOC1, and sequencing of GpFKF1 and GpGI cDNAs. The plants were also grown in the dark from 17:00 to 9:00 as a control. Far-red and blue LEDs had peaks at 740 and 465 nm, respectively (Hori et al., 2011). LEDs of either color supplied 10–15 μmol·m−2·s−1, respectively.
For expression analysis of GpFKF1 and GpGI, ‘Bristol Fairy’ plants with approximately 14 leaves were used for long-day (16 h) treatment with incandescent light in an incubator at 22°C, and the plants were also grown under the short-day condition of an 8-h photoperiod.
Cloning of cDNA encoding GpFKF1 and GpGI from G. paniculata and comparison of their sequences between ‘Bristol Fairy’ and ‘Million Star’
Total RNA was extracted from the leaves of ‘Bristol Fairy’ plants grown under long-day conditions as described previously (Hori et al., 2011). Extraction of total RNA, purification of poly (A)+ RNA, and reverse transcription were performed by the methods of Odanaka et al. (2002) and Kato et al. (2003). For cloning cDNA encoding GpFKF1, the primers for the PCR were 5'-CCA AGR GAY GTN GCW TCN ATY GG-3' and 5'-CAY GAN AGH GTR TGN CCC CA-3'. The primer sequences were based on the conserved sequences of FKF1 (AF216523) and its homologs in an ice plant (Mesembryanthemum crystallinum, AY371291) and other species. cDNAs for GpFKF1 containing the entire coding regions were cloned by 3'- and 5'-RACE (Moriguchi et al., 2006). For cloning cDNA encoding GpGI, the primers for the first PCR were 5'-CTC ATA WGA RCT RTA ACT CC-3' and 5'-TTC CTC AGC VGT TGA TYT KC-3' (Hecht et al., 2005). The PCR products were diluted and used for the second PCR using the primers 5'-ATG RAA ATW TTY GTN GCM ACH G-3' and 5'-GCT TSA TGR CTV ACR CAN ACR G-3'. The primer sequences for the second PCR were based on the conserved sequences of GI (AAF00092) and its homologs in the soybean (ABE81212), wheat (AAQ11738), and other species. cDNAs for GpGI containing the entire coding regions were cloned by 3'- and 5'-RACE (Moriguchi et al., 2006). cDNAs for GpFKF1 and GpGI were sequenced and then amino acid sequences deduced with GENETYX version 10 software (GENETYX CO., Tokyo, Japan).
Total RNA was extracted from the leaves of G. paniculata ‘Million Star’ plants using an RNeasy Plant Mini Kit (Qiagen, Hilden, Germany) and used for reverse transcription with a QuantiTect Reverse Transcription Kit (Qiagen). The nucleotide sequences of the cDNAs of GpFKF1 and GpGI were then determined by direct sequencing of the PCR products amplified with the primers designed from the nucleotide sequences of cDNAs cloned from ‘Bristol Fairy’. Alignments were generated by GENETYX to compare the amino acid sequences of ‘Bristol Fairy’ and ‘Million Star’.
Gene expression analysis
For the expression analysis of GpFT and GpSOC1, ‘Million Star’ leaves were sampled between 16:00 and 16:30 every 2 weeks, beginning 2 weeks after the start of 16-h light treatment. Extraction of RNA and real-time PCR were performed following Hori et al. (2011). The following primer sets were designed using Primer 3 software: 5'-TGG GTA GGC AAA CTG TGT ACG-3' and 5'-GCT CCA CAT ATC CAA AGA CGA TAC-3' for GpFT1 (AB562502); 5'-CAG TGT ACC CTC CCG TGT G-3' and 5'-GGT GAT TTA AGG CGG CAA C-3' for GpFT2 (AB562503); and 5'-TAT GGC GTG ATG CCA AAG G-3' and 5'-CGC TAC TAA CGC GCT TAA TCC-3' for GpSOC1 (AB562504). The nucleotide sequences of the primer sets for GpACT are described above.
For the expression analysis of GpFKF1 and GpGI, ‘Bristol Fairy’ leaves were sampled every 3 h from zeitgeber time (ZT) 0 to ZT 21 at 8 weeks after the start of long-day treatment. Extraction of RNA, separation of genomic DNA from the RNA sample, reverse transcription, and real-time PCR were performed following Hori et al. (2011). The following primer sets were designed using Primer 3 software: 5'-GAA CCA TTT GAG GCC ATT TG-3' and 5'-TTT GTG CCT TCC TTT GCT G-3' for GpGI; 5'-AAC CGT GTT GGA GGA CAC TC-3' and 5'-AAG GAC GAC CGA ATC ACA AG-3' for GpFKF1; and 5'-GGC CGT TCT TTC ATT GTA CG-3' and 5'-TCC TGC TCG AAG TCA AGA GC-3' for GpACT (AB562505).
Yeast two-hybrid assay
Yeast two-hybrid assays were performed with the Matchmaker Gold Yeast Two-Hybrid System (Clontech, Palo Alto, CA, USA). GpFKF1 containing the light-oxygen-voltage (LOV) domain and the F-box domain (Asp57 to Pro376), AtFKF1 containing the LOV domain and the F-box domain (Gly18 to Thr333), the N-terminal region of GpGI2 (Met1 to Phe411 with 12 bp of the 5'-UTR), and the N-terminal region of AtGI (Met1 to Leu402) were amplified with the corresponding specific primer pairs (Table 1). The PCR products of GpFKF1 and AtFKF1 were cloned into the pGADT7 AD prey vector to obtain the GpFKF1AD and AtFKF1AD prey vector. The PCR products of GpGI2 and AtGI were cloned into the pGBKT7 DNA-BD bait vector to obtain the GpGI2BD and AtGIBD bait vector. These cloning steps were performed with an In-Fusion HD Cloning Kit (Clontech). A Yeastmaker Yeast Transformation System 2 (Clontech) was used for transformation of prey and bait vectors to Y2HGold competent cells (Y2HGold). The Y2HGold yeasts with prey were each plated on synthetic, defined (SD)/-Leu, SD/-Leu/X-α-Gal, and SD/-Leu/X-α-Gal/aureobasidin A (AbA) media to test self-activation of each prey vector. The Y2HGold yeasts with bait were each plated on SD/-Trp, SD/-Trp/X-α-Gal, and SD/-Trp/X-α-Gal/AbA media to test the self-activation of each bait vector. Then, each set of prey and bait vectors was co-transformed into Y2HGold.

Quadruple dropout medium (QDO; SD/-Ade/-His/-Leu/-Trp) plates and QDO/AbA plates were used to assess the interaction between prey and bait proteins. The plates were incubated for 3 days at 30°C under blue LEDs (peak at 465 nm, 15 μmol·m−2·s−1) or in the dark. The set of pGADT7-T and pGBKT7-53 was used as the positive control and the set of empty prey pGADT7 and empty bait pGBKT7 was used as the negative control.
Cloning of the promoter regions of the GpFT1 gene
The 5'-upstream region of the GpFT1 genes was obtained using the Universal GenomeWalker 2.0 Kit (Clontech). Separate aliquots of genomic DNA were digested with four blunt-end restriction enzymes (DraI, EcoRV, PvuII, and StuI) and ligated to Genome Walker adaptors. The first PCR was performed using adaptor primer 1 (AP1) and gene-specific primers (GSP1). The second PCR was performed using a nested adapter primer 2 (AP2) and inner gene-specific primers (GSP2). The amplified PCR products were examined on an agarose gel, cloned into the pUC118 vector, and sequenced. The primers used are shown in Table 2.
Statistical analyses
Multiple comparisons by the Tukey–Kramer test were performed using Ekuseru-Toukei 2010 (Social Survey Research Information, Tokyo, Japan).
Results
Effect of blue light on flowering in ‘Million Star’
Because flowering is not promoted under long-day conditions with blue light in ‘Bristol Fairy’ (Hori et al., 2011), we investigated the effects of light quality on flowering in another cultivar, ‘Million Star’, for comparison. Flower budding began at 4 and 6 weeks after the start of 16-h light treatment with far-red or blue light, respectively (Fig. 1A). Flowering began at 8 weeks after the start of 16-h light treatment with far-red or blue light and the number of flowering shoots increased until 16 weeks (Fig. 1B). The numbers of budding shoots and flowering shoots per plant were very low under short-day conditions during the treatment. Flowering in ‘Million Star’ was thus promoted by 16-h light treatment not only with far-red light but also with blue light (Fig. 1).
Gene expression analysis of GpFT and GpSOC1 in ‘Million Star’
In ‘Bristol Fairy’, flowering under long-day conditions with blue light is not promoted and the levels of GpFT1, GpFT2, and GpSOC1 mRNA are low (Hori et al., 2011). We accordingly investigated the effects of light quality on the expression of GpFT and GpSOC1 in ‘Million Star’, in which flowering was promoted under long-day conditions with blue light (Fig. 1). GpFT1, GpFT2, and GpSOC1 mRNA levels were low under short-day conditions during the treatment (Fig. 2A, B, C). The GpFT1 mRNA level was higher in advance of flower budding under long-day conditions with blue light than under short-day conditions (Fig. 2A). GpFT2 and GpSOC1 mRNA levels increased from 4 weeks to 8 weeks after the start of the treatment under long-day conditions with blue light (Fig. 2B, C). The expression of GpFT1, GpFT2, and GpSOC1 was analyzed under long-day conditions with far-red light, which promoted the flowering of ‘Million Star’, and the genes showed high expression compared to that under short-day conditions (Fig. 2A, B, C). GpFT1 showed high expression under long-day conditions with far-red light compared to that with blue light at 4 and 6 weeks after the start of the treatment.

cDNAs containing the entire coding regions of the FKF1 and GI homologs, which were named GpFKF1 and GpGI, were cloned from G. paniculata ‘Bristol Fairy’ by RT-PCR with primers based on the conserved sequences and rapid amplification of cDNA ends (RACE). Because three different sequences of the FKF1 homologs were cloned and two of them showed extremely low expression, another sequence was used in this study, GpFKF1. LOV, F-box, and Kelch-repeat domains, as well as the amino acids essential for the function of FKF1, were well conserved in G. paniculata as shown by amino acid alignment (Han et al., 2004; Sawa et al., 2007; Song et al., 2012) (Fig. 3A). GpFKF1 was 69.9% and 86.4% identical to FKF1 from A. thaliana and the FKF1 homolog of Dianthus caryophyllus. GpFKF1 fell into the same cluster as the FKF1 homologs in a phylogenetic tree of deduced amino acid sequences of FKF1 and ZEITLUPE (ZTL) homologs (Fig. 3B).

Two different sequences of the GI homolog were cloned from G. paniculata and named GpGI1 and GpGI2. The deduced amino acid sequence of GpGI1 was 98.6% identical to that of GpGI2, and 73% and 95.7% identical to those of GI from A. thaliana and the GI homolog from D. caryophyllus, respectively. The amino acids conserved in 6 sequences of GI and the GI homologs used for reference were also conserved in GpGI1 and GpGI2 (Fig. 4A), and GpGI1 and GpGI2 fell into the same cluster as GI from D. caryophyllus in a phylogenetic tree (Fig. 4B).
Gene expression and interaction analysis of GpFKF1 and GpGI
The effect of long-day treatment on the expression of GpFKF1 and GpGI was analyzed in ‘Bristol Fairy’. The primers used in real-time PCR amplified both GpGI1 and GpGI2 cDNAs. The levels of GpFKF1 and GpGI mRNA showed a clear diurnal rhythm and their peaks differed between the long-day and short-day conditions (Fig. 5). GpFKF1 and GpGI mRNA levels showed peaks around dusk under the short-day condition and in the latter half of a light period under the long-day condition.

Interaction between the N-terminal region of GpGI and the LOV and F-box domains of GpFKF1 was analyzed by yeast two-hybrid assays with cDNAs cloned from ‘Bristol Fairy’ (Fig. 6). The sequence from GpGI2 cDNA was used for the assays because the amino acid sequences of GpGI1 and GpGI2 were almost identical and there was no difference in important amino acids between them. GpFKF1 interacted with GpGI2, and GpFKF1 and GpGI2 interacted with AtGI and AtFKF1, respectively. The same results were obtained in assays in the dark and under blue light.
Comparison of the amino acid sequences of GpFKF1 and GpGI of two cultivars
We compared the amino acid sequences of GpFKF1 and GpGI of ‘Bristol Fairy’ and ‘Million Star’ because the two cultivars showed a different flowering response to long-day conditions with blue light. The amino acid sequences of GpFKF1 and GpGI2 were identical between the two cultivars, but GpGI1 had some different amino acids (Fig. 4A). The differences found in GpGI1 between the two cultivars were inferred not to be essential for function because the differing amino acids were not in conserved regions or were substituted by similar amino acids in conserved regions.
Sequence comparison of the GpFT1 promoter region between ‘Bristol Fairy’ and ‘Million Star’
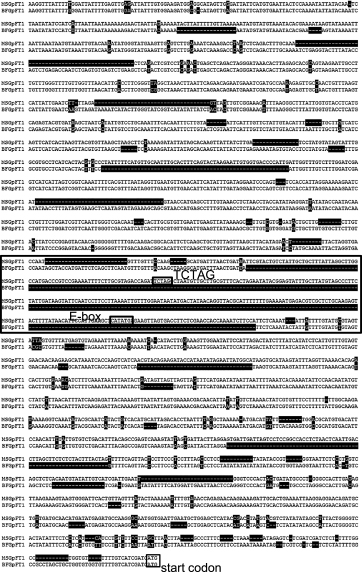
Sequences of the GpFT1 promoter region were compared between the two cultivars because the GpFT1 mRNA level increased in advance of flower budding under long-day conditions with blue light in ‘Million Star’. There were several deletions and nucleotide substitutions and, in particular, the promoter region from ‘Bristol Fairy’ contained a large deletion corresponding to −1080 bp to −1382 bp upstream of the start codon of GpFT1 in ‘Million Star’ (Fig. 7). The deleted region contained an E-box (CANNTG) and a TCTAG sequence, which are reportedly associated with the response to blue light through the CRY pathway (Liu et al., 2013; Shaikhali et al., 2012).
Discussion
In our previous study (Hori et al., 2011), flowering was not promoted under long-day conditions with blue light in G. paniculata ‘Bristol Fairy’ despite the promotion of flowering by blue light in A. thaliana. We accordingly investigated the effect of blue light on flowering in other cultivars and found that flowering of ‘Million Star’ is promoted under long-day conditions with blue light. Our results revealed that there is variation in G. paniculata’s flowering response to blue light. Although the expression of GpFT and GpSOC1 in ‘Bristol Fairy’ is induced under long-day conditions with white light, their transcripts are low under long-day conditions with blue light, which does not promote flowering (Hori et al., 2011). In contrast, in the present study, the expression of those genes was induced under long-day conditions with blue light in ‘Million Star’. In particular, GpFT1 expression was higher in advance of flower budding under long-day conditions with blue light than under short-day conditions in ‘Million Star’. Under long-day conditions with far-red light, flowering is promoted in ‘Bristol Fairy’ with the expression of GpSOC1, whereas the expression of GpFT is not induced (Hori et al., 2011). Flowering is not promoted under long-day conditions with blue light in the ft mutant of A. thaliana unlike the response in the wild type, although flowering is promoted under long-day conditions with far-red light in the ft mutant and the wild type (Hori et al., 2011). This difference in the response to light quality between the ft mutant and the wild type in A. thaliana seems to have commonalities with the difference between ‘Bristol Fairy’ and ‘Million Star’ in G. paniculata.
We analyzed the G. paniculata homologs of FKF1 and GI, which act upstream of FT in a blue light-dependent manner in A. thaliana (Sawa et al., 2007; Song et al., 2012), because these genes may play an important role in G. paniculata’s flowering response to blue light. In addition, there is a lack of information on the flowering response to blue light in long-day cut flowers and there is, to our knowledge, no report of FKF1 and GI in cut flowers. The amino acid sequences of GpFKF1, GpGI1, and GpGI2 were found to be well conserved in a sequence comparison with the proteins from A. thaliana and other species. They were classified into FKF1 and GpGI groups, respectively, and fell into the same clusters as the homologs of D. caryophyllus, which is a member of Caryophyllaceae with G. paniculata, in phylogenetic analysis. GpFKF1 was classified into the FKF1 group in the phylogenetic tree containing the groups of FKF1 and ZTL, which has the same domain as FKF1 but a different function from FKF1. In GpFKF1, the amino acid sequences were well conserved in the three important domains of LOV, F-box, and Kelch repeats, and amino acids that are essential for the reception of blue light, interaction with GI, and ubiquitin ligase activity (Han et al., 2004; Sawa et al., 2007; Song et al., 2012) were conserved..
In A. thaliana, expression patterns of FKF1 and GI show circadian rhythms, and peaks around the dusk and in the latter half of a light period, occurring several hours before the dusk, are observed under short-day and long-day conditions, respectively (Fowler et al., 1999; Imaizumi et al., 2003). The expression patterns of GpFKF1 and GpGI in G. paniculata were similar to those of FKF1 and GI in A. thaliana. These expression patterns result in the translation and accumulation of FKF1 and GI proteins under long-day conditions, and FKF1 can interact with GI during a light period for the blue light-dependent degradation of CDF1 and its related proteins, which are the transcriptional repressors of CO and FT (Imaizumi et al., 2005; Sawa et al., 2007; Song et al., 2012). There is thus a possibility that GpFKF1 and GpGI have roles in blue light response in G. paniculata.
The LOV and F-box domains of FKF1 and the N-terminal region of GI have been reported to be necessary for their interaction (Sawa et al., 2007). The LOV and F-box domains of GpFKF1 and the N-terminal region of GpGI2 were used for the interaction assays in this study. As a result, GpFKF1 interacted with GpGI2; the interaction was also observed between the proteins from G. paniculata and A. thaliana. Because FKF1 contains the LOV domain for blue light sensing, we investigated the effect of blue light on the interaction. However, interactions were observed irrespective of the presence or absence of blue light. Further study will be needed to clarify this point. In addition to the amino acid sequences and expression patterns of GpFKF1 and GpGI, their interaction, which could be important for their function, was observed in ‘Bristol Fairy’ in the present study. Given that CRY1 and CRY2 are known to be blue light receptors in addition to FKF1 (Thomas, 2006; Wang et al., 2014), it cannot be ruled out that the variation in the blue light response of flowering is associated with the CRY-mediated flowering pathway.
Variation in the flowering response to blue light was found in G. paniculata. No defects were found in GpFKF1 and GpGI of ‘Bristol Fairy’ in this study, and its possible mechanism was associated with GpFT and GpSOC1. As one candidate, the nucleotide sequences of the promoter region of GpFT1 differed between ‘Bristol Fairy’ and ‘Million Star’, and large deletions that could affect the expression were found in ‘Bristol Fairy’. Because CO is also essential for the regulation of flowering in A. thaliana, differences in the CO homolog between the two cultivars should be investigated in G. paniculata. In contrast to the situation with red and far-red light, there have been very few useful studies of the flowering response to blue light in cut flowers. Our results will thus be useful for research and technological development in the control of flowering time in cut flowers in lighting culture and plant factories.
Acknowledgment
The authors thank Mr. T. Takahashi for his help.
Literature Cited
- Andrés, F. and G. Coupland. 2012. The genetic basis of flowering responses to seasonal cues. Nature Rev. Genet. 13: 627–639.
- Corbesier, L., C. Vincent, S. Jang, F. Fornara, Q. Fan, I. Searle, A. Giakountis, S. Farrona, L. Gissot, C. Turnbull and G. Coupland. 2007. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316: 1030–1033.
- Eskins, K. 1992. Light-quality effects on Arabidopsis development. Red, blue and far-red regulation of flowering and morphology. Physiol. Plant. 86: 439–444.
- Fornara, F., K. C. S. Panigrahi, L. Gissot, N. Sauerbrunn, M. Rühl, J. A. Jarillo and G. Coupland. 2009. Arabidopsis DOF transcription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response. Dev. Cell 17: 75–86.
- Fowler, S., K. Lee, H. Onouchi, A. Samach, K. Richardson, B. Morris, G. Coupland and J. Putterill. 1999. GIGANTEA: A circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. EMBO J. 18: 4679–4688.
- Guo, H., H. Yang, T. C. Mockler and C. Lin. 1998. Regulation of flowering time by Arabidopsis photoreceptors. Science 279: 1360–1363.
- Han, L., M. Mason, E. P. Risseeuw, W. L. Crosby and D. E. Somers. 2004. Formation of an SCFZTL complex is required for proper regulation of circadian timing. Plant J. 40: 291–301.
- Hecht, V., F. Foucher, C. Ferrándiz, R. Macknight, C. Navarro, J. Morin, M. E. Vardy, N. Ellis, J. P. Beltrán, C. Rameau and J. L. Weller. 2005. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 137: 1420–1434.
- Hori, Y., K. Nishidate, M. Nishiyama, K. Kanahama and Y. Kanayama. 2011. Flowering and expression of flowering-related genes under long-day conditions with light-emitting diodes. Planta 234: 321–330.
- Imaizumi, T., T. F. Schultz, F. G. Harmon, L. A. Ho and S. A. Kay. 2005. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 309: 293–297.
- Imaizumi, T., H. G. Tran, T. E. Swartz, W. R. Briggs and S. A. Kay. 2003. FKF1 is essential for photoperiodic-specific light signaling in Arabidopsis. Nature 426: 302–306.
- Kato, K., Y. Okamura, K. Kanahama and Y. Kanayama. 2003. Nitrate independent expression of plant nitrate reductase in Lotus japonicus root nodules. J. Exp. Bot. 54: 1685–1690.
- Kikuchi, K., Y. Kanayama, Y. Wakamoto and K. Kanahama. 2000. Effect of seedling age, photoperiod and temperature on bolting and inflorescence quality in Delphinium. J. Japan. Soc. Hort. Sci. 69: 446–448.
- Kristinsen, K. 1988. Light quality regulates flower initiation, differentiation and development of Campanula carpatica Jacq. ‘Karl Forster.’ Sci. Hortic. 35: 275–283.
- Liu, Y., X. Li, K. Li, H. Liu and C. Lin. 2013. Multiple bHLH proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis. PLOS Genet. 9: e1003861.
- Moriguchi, R., K. Kanahama and Y. Kanayama. 2006. Characterization and expression analysis of the tomato telomere-binding protein LeTBP1. Plant Sci. 171: 166–174.
- Nakajima, R., S. Otagaki, K. Yamada, K. Shiratake and S. Matsumoto. 2014. Molecular cloning and expression analyses of FaFT, FaTFL, and FaAP1 genes in cultivated strawberry: their correlation to flower bud formation. Biol. Plant. 58: 641–648.
- Nishidate, K., Y. Kanayama, M. Nishiyama, T. Yamamoto, Y. Hamaguchi and K. Kanahama. 2012. Far-red light supplemented with weak red light promotes flowering of Gypsophila paniculata. J. Japan. Soc. Hort. Sci. 81: 198–203.
- Odanaka, S., A. B. Bennett and Y. Kanayama. 2002. Distinct physiological roles of fructokinase isozymes revealed by gene-specific suppression of Frk1 and Frk2 expression in tomato. Plant Physiol. 129: 1119–1126.
- Sato, T., N. Kudo, T. Moriyama, H. Ohkawa, Y. Kanayama and K. Kanahama. 2009. Acceleration of flowering of Eustoma grandiflorum in early winter by day-extension treatments with far-red rich bulbtype fluorescent lamps. Hort. Res. (Japan) 8: 327–334 (In Japanese with English abstract).
- Sawa, M., D. A. Nusinow, S. A. Kay and T. Imaizumi. 2007. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 318: 261–265.
- Shaikhali, J., J. D. Barajas-Lopéz, K. Ötvös, D. Kremnev, A. S. Garcia, V. Srivastava, G. Wingsle, L. Bako and A. Stranda. 2012. The CRYPTOCHROME1-dependent response to excess light is mediated through the transcriptional activators ZINC FINGER PROTEIN EXPRESSED IN INFLORESCENCE MERISTEM LIKE1 and ZML2 in Arabidopsis. Plant Cell 24: 3009–3025.
- Shibuya, T. and Y. Kanayama. 2014. Flowering response to blue light and its molecular mechanisms in Arabidopsis and horticultural plants. Adv. Hort. Sci. 28: 179–183.
- Shillo, R. and A. H. Halevy. 1982. Interaction of photoperiod and temperature in flowering-control of Gypsophila paniculata. Sci. Hortic. 16: 385–393.
- Song, Y. H., R. W. Smith, B. J. To, A. J. Millar and T. Imaizumi. 2012. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 336: 1045–1049.
- Thomas, B. 2006. Light signals and flowering. J. Exp. Bot. 57: 3387–3393.
- Wang, X., Q. Wang, P. Nguyen and C. Lin. 2014. Cryptochrome-mediated light responses in plants. Enzymes 35: 167–189.
- Wigge, P. A., M. C. Kim, K. E. Jaeger, W. Busch, M. Schmid, J. U. Lohmann and D. Weigel. 2005. Integration of Spatial and temporal information during floral induction in Arabidopsis. Science 309: 1056–1059.
- Xing, W., Z. Wang, X. Wang, M. Bao and G. Ning. 2014. Over-expression of an FT homolog from Prunus mume reduces juvenile phase and induces early flowering in rugosa rose. Sci. Hortic. 172: 68–72.
- Yagi, M., S. Kosugi, H. Hirakawa, A. Ohmiya, K. Tanase, T. Harada, K. Kishimoto, M. Nakayama, K. Ichimura, T. Onozaki, H. Yamaguchi, N. Sasaki, T. Miyahara, Y. Nishizaki, Y. Ozeki, N. Nakamura, T. Suzuki, Y. Tanaka, S. Sato, K. Shirasawa, S. Isobe, Y. Miyamura, A. Watanabe, S. Nakayama, Y. Kishida, M. Kohara and S. Tabata. 2014. Sequence analysis of the genome of carnation (Dianthus caryophyllus L.). DNA Res. 21: 231–241.
- Yamada, A., T. Tanigawa, T. Suyama, T. Matsuo and T. Kunitake. 2008a. Night break treatment using different light sources promotes or delays growth and flowering of Eustoma grandiflorum (Raf.) Shinn. J. Japan. Soc. Hort. Sci. 77: 69–74.
- Yamada, A., T. Tanigawa, T. Suyama, T. Matsuo and T. Kunitake. 2008b. Improvement of Eustoma grandiflorum (Raf.) Shinn. cut flower quality for early-autumn shipping with long-day treatment using light source that delay flower bud formation. J. Japan. Soc. Hort. Sci. 77: 296–303.
- Yamada, A., T. Tanigawa, T. Suyama, T. Matsuo and T. Kunitake. 2009. Red: far-red light ratio and far-red light integral promote or retard growth and flowering in Eustoma grandiflorum (Raf.) Shinn. Sci. Hortic. 120: 101–106.
- Yoo, S. K., K. S. Chung, J. Kim, J. H. Lee, S. H. Hong, S. J. Yoo, S. Y. Yoo, J. S. Lee and J. H. Ahn. 2005. CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis. Plant Physiol. 139: 770–778.
- Yoshimura, M., M. Nishiyama and K. Kanahama. 2002. Effects of red or far-red light and red/far-red ratio on the shoot growth and flowering of Matthiola incana. J. Japan. Soc. Hort. Sci. 71: 575–582 (In Japanese with English abstract).
- Yoshimura, M., A. Sasaki, T. Moriyama, Y. Shibata, K. Katsuta and K. Kanahama. 2006. Effects of various light sources for night irradiation and light intensity on the flowering of stock (Matthiola incana (L.) R. Br.) plant. Hort. Res. (Japan) 5: 297–301 (In Japanese with English abstract).