ORIGINAL ARTICLES
Seasonal Changes in Functional Component Contents in Mulberry (Morus alba L.) Leaves
2017 Volume 86 Issue 4 Pages 534-542

Details
2017 Volume 86 Issue 4 Pages 534-542
The aim of this study was to determine the best time to harvest mulberry (Morus alba L.) leaves for optimal yields of functional components. Mulberry leaves were analyzed to determine the seasonal changes in the concentrations of functional components, including flavonols, chlorogenic acid, and 1-deoxynojirimycine (DNJ). During the experimental period (May 26–October 16), the ranges of flavonols, chlorogenic acid, and DNJ contents were 1134–2230 mg/100 g dry weight (DW), 616–1014 mg/100 g DW, and 53–199 mg/100 g DW, respectively. There were high flavonols contents in mulberry leaves from late May to early July, from early to mid-August, and after late September. There were high chlorogenic acid contents from late May to early July, and after late September. From late September, the flavonols contents increased over time, and showed the highest level on October 16, the last day of the experimental period. The increased flavonols contents after late September were due to the synergistic effects of solar radiation and lower temperatures. The DNJ content showed a bell-shaped curve, peaking in mid-August, and the DNJ content was strongly correlated with total temperature. The flavonols and chlorogenic acid contents in mulberry leaves were higher after late September when the temperatures decreased and there was still high solar radiation, while the DNJ content peaked in August when temperatures were highest. The results indicated that August is the best time to harvest mulberry leaves to obtain optimal yields of beneficial functional components. If mulberry leaves are harvested twice per year, then the first harvest should be in July to August when temperatures and solar radiation hours are high, and the second harvest should be completed by the end of September.
Recently, there has been increasing interest in healthy food, and the functional benefits of edible plants have attracted much attention. The functional components of mulberry (Morus alba L.) trees, which were originally cultivated to feed silk worms, have been evaluated in various studies (Butt et al., 2008; Chauhan et al., 2015; Enkhmaa et al., 2005; Evans et al., 1985; Nakagawa, 2013; Tsuduki et al., 2013). There has been a dramatic decrease in mulberry production for sericulture, but an increase in its production for human foods such as mulberry tea and fresh green juice. Among the various functional components of mulberry leaves, flavonols have been shown to have strong antioxidant activity, as well as inhibitory activities against hypertension, arteriosclerosis, and the growth of cancer cells (Enkhmaa et al., 2005; Katsube et al., 2006; Naowaratwattana et al., 2010). Previously, we reported that quercetin 3-(6-malonyglucoside) is the major flavonol glycoside in mulberry leaves, and that it has a strong antioxidant effect (Katsube et al., 2006). We also reported that it altered blood glucose levels via a mechanism different from that of 1-deoxynojirimycin (DNJ) (Katsube et al., 2010). Our research group reported that chlorogenic acid, an abundant polyphenol in mulberry leaves, plays an important role in the antioxidant activity of leaf extracts (Katsube et al., 2009). It is also well known that DNJ, another functional component of mulberry leaves, acts as an α-glucosidase inhibitor to elevate blood glucose levels (Evans et al., 1985).
Mulberry leaves are harvested once or twice per year as a raw material to produce various foods. The mulberry plants are cultivated using technology that was established for producing silkworm feed. However, these methods may not produce plants with the highest levels of the targeted functional components, and so some modifications of the cultivation technology may be necessary. Our previous study showed the effects of cultivation conditions on the functional component contents of mulberry leaves. Our results showed that the flavonols contents of mulberry leaves differ depending on the variety, that the amount of solar radiation affects flavonols and DNJ contents, and that the amounts of applied nitrogen affect the concentrations of chlorogenic acid, flavonol, and DNJ (Sugiyama et al., 2013, 2016a, b).
It is well known that the harvest time affects the catechin contents of green tea and black tea (Kondo et al., 2007; Takayanagi et al., 1985). Consequently, we hypothesized that the harvest time may also affect the functional component concentrations in mulberry leaves. Kimura et al. (2007) and Nakanishi et al. (2011) reported that the contents of DNJ and α-glucosidase inhibitors increased in August, the hottest month of the year. In terms of polyphenol contents in mulberry leaves, Lee and Choi (2012) reported that the chlorogenic acid and flavonols contents were highest in May when leaves were harvested three times per year (in May, July, and September), although their results did not document seasonal changes in chlorogenic acid and flavonols contents. To obtain the highest yields of total functional components, it is necessary to determine the seasonal changes in flavonol, chlorogenic acid, and DNJ contents in mulberry leaves. This information can be used to identify the best harvest time to obtain the highest yields of beneficial functional components. Some compromises may be necessary, because the concentrations of the functional components may peak at different times. For example, the optimal solar radiation and nitrogen fertilization conditions for high flavonols content differed from those for high DNJ contents (Sugiyama et al., 2016a, b). In the present study, we analyzed mulberry leaves to determine the seasonal changes in the contents of functional components, including flavonols, chlorogenic acid, and DNJ, and investigated the reasons for the differences in their concentrations to identify the optimum harvest time.
Seasonal changes in the functional components in mulberry leaves were investigated at the experimental field of Shimane Agricultural Technology Center (Izumo, Shimane, Japan) in 2015. Five-year-old trees of the mulberry ‘Ichinose’ were used and were cultivated and fertilized according to established methods. Before bud break in spring, the mulberry trees were pruned at the base of the shoot, and then the re-grown shoots were used as the materials in this study. Using two shoots from each stock, the youngest open leaves with completely opened leaflets at 20 cm from the tip of the shoot and the neighboring two leaves were sampled, that is, six leaves per stock. The six leaves from each stock were grouped together for analysis. Mulberry leaves were sampled every 10 days from May 25 (when the shoots were longer than 40 cm) to October 16. As mulberry leaves are generally harvested twice a year, two experimental groups were defined: a once-harvested group and a twice-harvested group, to evaluate the effect of pruning. For the twice-harvested group, the shoots of mulberry trees were pruned at 80 cm above ground level on July 15. In the twice-harvested group, leaves were not sampled after re-emergence of buds on July 24 because the leaves were too small. A total of 30 stocks were used for experimental analysis of each treatment. Tree growth and yield were evaluated on October 26 for the once-harvested group and on both July 15 and October 26 for the twice-harvested group. Tree growth was evaluated using the longest shoot of each stock. For new shoots, the length from the base to the tip of each shoot was measured. For shoot length without leaves after defoliation, the length from the base of the shoot to the bottom of the remaining leaf was measured. Leaf length and internode length were evaluated at the uppermost one-third of the shoot, and the internode length was defined as the average of five successive nodes. Shoot weight and leaf yield were evaluated after removing all the sample shoots from each stock. Stem weight was measured after removing all the leaves from the sample shoot. The leaf yield was defined as the leaf weight, which was calculated by subtracting the stem weight without leaves from the total shoot weight with leaves. The two groups were replicated three times in the same field, and 10 stocks were evaluated for each group. To determine the growth and yield of mulberry trees, the average value of the 30 stocks (10 stocks per field segment) was calculated. At harvest, all the leaves of the longest shoot of each stock were sampled to determine the contents of functional components per stock.
Meteorological data were obtained from the Automated Meteorological Data Acquisition System located in Izumo, Shimane, Japan. Figure 1 shows the total hours of solar radiation and total temperature for 14 days before leaf sampling. The database of the Automated Meteorological Data Acquisition System was searched to obtain the average temperature between 1981 and 2010 and the hours of solar radiation between 1987 and 2010. Total temperature was based on the average temperature per day, and total temperature and total hours of solar radiation were calculated for a period of 14 days before leaf sampling. This 14-day duration was selected because one leaf on a mulberry shoot unfolds every 1.4 days in June and July. Therefore, the 14-day totals for temperature and solar radiation represented the entire growth period of the middle leaf (10th from the shoot tip) sampled at 14 days after the onset of leaf opening.

Total hours of solar radiation and total temperature for 14 days before leaf sampling in 2015 and average yearly valuesz.
z The database of the Automated Meteorological Data Acquisition System was searched to calculate the average temperature between 1981 and 2010 and the average hours of solar radiation between 1987 and 2010. The values for a period of 14 days before leaf sampling were used as standard values. Total temperature for 14 days was calculated by adding average temperature per day.
The effect of temperature on DNJ content was investigated using artificial climate rooms at our institution in 2015. Ten two-year-old nursery stocks of the mulberry ‘Hayatesakari’ were used for these analyses. Each tree was cultivated in a 25-L plastic pot and trees were pruned in summer. After pruning, the shoots were trimmed, leaving only two new shoots. Five mulberry trees were used for each temperature treatment. On October 2, five plants per group were transferred from outdoors to an artificial climate room, one set at a high temperature (25–30°C) and the other set to a low temperature (18–23°C). The photoperiod in both climate rooms was 12-h light/12-h dark. On October 16 and November 6, leaves located 20 cm from the shoot tip were sampled along with 10 μL of the latex that oozed from the leaf petiole. These samples were analyzed to determine DNJ contents.
Extraction procedureSampled leaves were dried in a convection oven (MOV-212F; Sanyo Electric, Inc., Osaka, Japan) at 60°C for 36–48 h. The dried samples were ground into a fine powder using a food mill (IFG-700G; Iwatani, Inc., Tokyo, Japan) and then used for the quantitative determination of functional components. Dry mulberry leaf powder (100 mg) was suspended in 10 mL 60% ethanol aqueous (v/v) and stirred for 3 h at 30°C in an incubator (LTI-600 SD; Tokyo Rikakikai, Inc., Tokyo, Japan). After centrifugation at 10000 × g for 5 min, the extracted solution was filtered through a 0.45-μm filter (Advantec MFS, Inc., Tokyo, Japan), and used for the quantitative analysis of chlorogenic acid, flavonols, and DNJ. The latex samples were diluted with water and then analyzed to determine DNJ contents.
Quantification of chlorogenic acid, flavonols, and DNJChlorogenic acid, flavonols, and DNJ in mulberry leaves were analyzed using an ultra-performance liquid chromatograph (Nexera UPLC System; Shimadzu Corporation, Tokyo, Japan) which consisted of a LC-30AD dual pump, a SIL-30AC auto sampler, a CTO-20AC column temperature controller, a SPD-M20A diode array detector, an RF-20Axs fluorescence detector, and a Shim-pack XR-ODS column (50 × 2.0 mm; Shimadzu).
To analyze the chlorogenic acid and flavonols contents, the column temperature was set to 40°C, and a binary gradient consisting of 89%–78% acetonitrile containing 0.1% formic acid was applied at a flow rate of 0.6 mL·min−1. The wavelengths to detect chlorogenic acid and flavonols were 280 nm and 350 nm, respectively. We quantified DNJ according to the method of Kim et al. (2003). Standard flavonols, kaempferol 3-(6-rhamnosylglucoside), quercetin 3-(6-malonylglucoside), and kaempferol 3-(6-malonylglucoside) were purified from mulberry leaves as described by Katsube et al. (2006). Rutin was purchased from Wako Chemicals Inc. (Osaka, Japan), and isoquercitrin, astragalin, and DNJ were purchased from Funakoshi, Inc. (Tokyo, Japan). The flavonol content in leaves of the mulberry ‘Ichinose’ and ‘Hayatesakari’ was the sum of six flavonol glycosides: rutin, isoquercitrin, kaempferol 3-(6-rhamnosylglucoside), quercetin 3-(6-malonylglucoside), astragalin, and kaempferol 3-(6-malonylglucoside).
Statistical analysesData were analyzed with Microsoft Excel 2013 (Microsoft Co., WA, USA) and Excel Toukei 2015 for Windows (Social Survey Research Information Co., Ltd., Tokyo, Japan). Results are expressed as the means ± standard error (SE). Data were tested by one-way analysis of variance, followed by Tukey’s test for multiple comparisons.
We evaluated the seasonal changes in flavonol, chlorogenic acid, and DNJ contents in mulberry leaves from May 26 to October 16 in 2015 (Fig. 2A–C). The flavonols contents ranged from 1134 to 2230 mg/100 g dry weight (DW), showing low levels from mid to late July and from early to mid-September, and high levels from late May to early July, from early to mid-August, and after late September (Fig. 2A). The flavonols contents continued to increase after late September and showed the highest level on October 16, the last day of the experiment.

Seasonal changes in functional component contents in mulberry leavesz. A: Flavonol, B: Chlorogenic acid, C: DNJ.
z The youngest open leaves with completely opened leaflets at 20 cm from the shoot tip and the subsequent two leaves were sampled. Data are mean ± SE (n = 30).
The chlorogenic acid content also varied, ranging from 616 to 1014 mg/100 g DW, with low levels from mid-July to early August and early September and high levels from late May to early July and after late September (Fig. 2B). The seasonal changes in chlorogenic acid and flavonols contents were similar, except in August. The flavonols contents in mulberry leaves were lowest on July 24 and highest on October 16.
The DNJ content ranged from 53 to 199 mg/100 g DW, showing a bell-shaped curve with its peak in mid-August and very low values in May and October (Fig. 2C). It increased slightly until mid-July, and then sharply increased until mid-August, before sharply decreasing after late August.
The relationships among the contents of functional components in mulberry leaves, total hours of solar radiation, and total temperature were analyzed by setting the first day of sampling as the base point (Table 1). The correlation coefficient between flavonols content and total hours of solar radiation was r = 0.59 (P < 0.01) and that between flavonols content and total temperature was r = −0.51 (P < 0.05). The multiple correlation coefficient among total hours of solar radiation, total temperature, and flavonols content was r = 0.79 (P < 0.01). The correlation coefficient between the chlorogenic acid content and total hours of solar radiation was low (r = 0.33), but that between the chlorogenic acid content and total temperature was high (r = −0.69; P < 0.01). The multiple correlation coefficient among total hours of solar radiation, total temperature, and chlorogenic acid content was r = 0.77 (P < 0.01). The DNJ content was strongly correlated with total temperature (r = 0.94; P < 0.01), but not with total hours of solar radiation (r = 0.05).

Correlation coefficients between functional component contents in mulberry leaves and total hours of solar radiation or total temperature for 14 days before samplingz.
To investigate the effect of pruning, the amounts of functional components were compared between the once-harvested and twice-harvested groups (Fig. 2A–C). The flavonols contents and the DNJ content in the twice-harvested group were about the same as those in the once-harvested group, but the chlorogenic acid content was higher in the twice-harvested group than in the once-harvested group.
Effect of high temperature on DNJ contentThe DNJ contents varied depending on total temperature in the field-grown plants (Fig. 2C). Therefore, the effect of temperature on DNJ content was investigated in detail using plants grown in artificial climate rooms. On October 2, 2015, mulberry trees in plastic pots were transferred from outdoors into the artificial climate rooms, and then the DNJ contents in leaves and latex were measured. The DNJ contents in leaves in the high-temperature room (25–30°C) were slightly higher at 14 days after transfer from outdoors, and then remained at an almost constant level. However, the DNJ contents in leaves in the low-temperature room (18–23°C) continuously decreased (Fig. 3). The DNJ contents in latex continuously increased in the high-temperature room, but remained almost constant in the low-temperature room.

Effect of temperature on DNJ content in leaves and latex of mulberryz. A: leaves, B: latex.
z DNJ content was analyzed in mulberry trees cultivated in plastic pots, which were transferred from outdoors to artificial climate rooms on October 2. Five mulberry trees were used for each temperature treatment. Leaves located at 20 cm from shoot tip were sampled. Data are expressed as mean ± SE (n = 5).
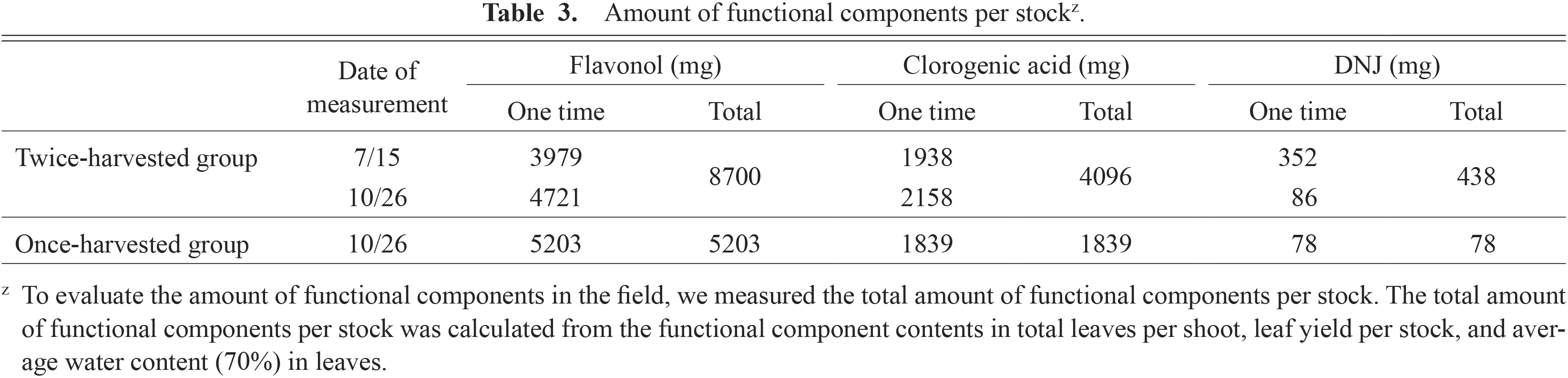
The amount of functional components per stock was evaluated based on the functional component contents in leaves and leaf yields. Mulberry leaves are harvested by plucking them from new shoots. When there are two harvests per year, the first harvest is generally in July, and then the re-grown shoots are harvested in September. Table 2 shows mulberry tree growth and leaf yield at harvest time. The leaf yield of the once-harvested group was higher than that of the twice-harvested group in October, although leaf yield of the once-harvested group was less than half the sum of the twice-harvested group. The shoot length without leaves after defoliation in the once-harvested group was approximately three times longer than that in the twice-harvested group.

Mulberry tree growth and leaf yield at harvest.
Previously, we showed that the functional component contents of mulberry leaves depends on the leaf’s position on the shoot. The levels of flavonols and DNJ contents were higher in the upper leaves and lower in the lower leaves, and the upper–lower leaf difference was greater for DNJ contents than flavonols (Sugiyama et al., 2016a). To determine the amounts of functional components per stock, we calculated the functional component contents for all of the leaves on one shoot (Fig. 4A–C). Then, the total amount of functional components per stock was calculated based on the functional component contents in total leaves, leaf yield, and water content (Table 3). The flavonols content per shoot in the twice-harvested group was highest on October 26 (Fig. 4A). The chlorogenic acid content per shoot in the twice-harvested group was highest on October 26, and that in the twice-harvested group on July 15 was same as that in the once-harvested group on October 26 (Fig. 4B). Based on these results, the total amounts of flavonols and chlorogenic acid per stock were 1.7-fold and 2.2-fold greater, respectively, in the twice-harvested group than in the once-harvested group (Table 3). The lowest DNJ content per shoot was in the once-harvested group (Fig. 4C). Interestingly, the total amount of DNJ per stock was 438 mg/stock in the twice-harvested group, but only 78 mg/stock in the once-harvested group (Table 3).

Functional component contents in mulberry leaves per shoot at harvestz. A: Flavonols, B: Chlorogenic acid, C: DNJ.
z Because the functional component contents of mulberry leaves differ depending on the position of the leaf, all the leaves of each shoot were used to determine its functional component contents. All the leaves on the longest shoot of each stock were sampled to evaluate the functional component contents in bulk. Different letters indicate statistically significant differences (P < 0.01; Tukey’s test).
Amount of functional components per stockz.
We analyzed mulberry leaves to determine the seasonal changes in the concentrations of functional components, including flavonols, chlorogenic acid, and DNJ, to determine the optimal harvest time to obtain the maximum yield of functional components.
The functional component contents in mulberry leaves varied widely throughout the experimental period. There were high levels of flavonols and chlorogenic acid (polyphenols) after late September. The flavonols content markedly increased from mid-September, and peaked in October. This result differed from those of previous reports, which stated that polyphenol contents in leaves were highest in spring and early summer. For example, Kawakami et al. (2010) reported that the polyphenols content in persimmon leaves peaked in June, and then decreased gradually. Takahashi and Okiura (2013) reported that the rutin content in fig leaves was highest in May and June. Midorikawa et al. (2010) reported that the concentrations of caffeic acid derivatives in Ilex latifolia were higher in spring than in autumn. In contrast, Sonehara and Izumi (1991) reported that the polyphenols contents in persimmon leaves increased from June, and peaked in October. Lee and Choi (2012) reported that the chlorogenic acid and flavonols contents in mulberry leaves were highest in May when there were three harvests per year (in May, July, and September). However, our results showed that the flavonols and chlorogenic acid contents were highest on October 16, consistent with the findings of Sonehara and Izumi (1991).
The reasons for the differences between our results and those of previous studies may have been caused by differences in the functional components examined, the methods used, and the target species. For example, Kawakami et al. (2010) measured water-soluble polyphenol in persimmon leaves using the Folin–Denis method; thus, the components they examined and the method used were different from ours. Furthermore, the growth shapes of persimmon trees and mulberry trees are very different. That is, persimmon trees stop growing branches and new shoots extend in the lateral direction. Therefore, the direction that light hits persimmon leaves is different from that hitting mulberry leaves.
Our experimental method was closer to that of Takahashi and Okiura (2013), who measured rutin in the upper leaves of fig trees using high-performance liquid chromatography. Furthermore, the time before new growth ends and the pattern of new shoot growth in fig trees are similar to those in mulberry trees. Nevertheless, the time when rutin content peaked in Takahashi and Okiura’s study was different from our results. However, we did find that flavonols contents in May and June were as high as those in August, which is similar to their findings. Although the hours of solar radiation in the Takahashi and Okiura’s study are unknown, an abnormal amount of solar radiation hours may be one reason for the differences between our results and theirs. In our study carried out in 2015, the pattern of total hours of solar radiation from May to mid-June and early- to mid-August was similar to that of normal years (Fig. 1). However, from mid-late September to early October in 2015, the total hours of solar radiation were higher than average. Thus, the relatively high solar radiation may partially explain such variance.
Our results showed that the flavonols contents were positively correlated with the total hours of solar radiation and negatively correlated with total temperature (Table 1). In our previous report, we found a positive correlation between flavonols contents and total hours of solar radiation by investigating the functional component contents of mulberry leaves every 10 days in common fields from May 28 to July 28, 2009 and May 24 to July 27, 2010 (Sugiyama et al., 2016a). The correlation coefficients between chlorogenic acid contents and total hours of solar radiation and total temperature in 2009 were r = 0.67 (P = 0.18) and r = −0.57 (P = 0.25) and those in 2010 were r = 0.40 (P = 0.45) and r = −0.30 (P = 0.58), respectively. The multiple correlation coefficients for chlorogenic acid contents were r = 0.81 (P = 0.19) in 2009 and r = 0.47 (P = 0.60) in 2010. The correlation coefficients between flavonols contents and total solar radiation and total temperature were r = 0.94 (P = 0.01) and r = −0.33 (P = 0.32) in 2009 and r = 0.83 (P = 0.01) and r = −0.49 (P = 0.08) in 2010, respectively. The multiple correlation coefficients for flavonols were r = 0.96 (P = 0.02) in 2009 and r = 0.93 (P = 0.02) in 2010 (data not shown). Although the chlorogenic acid contents in that study did not show a significant correlation with either total solar radiation or total temperature, the flavonols contents did have a positive correlation with total hours of solar radiation. Furthermore, the multiple correlation coefficients between flavonols and total hours of solar radiation and total temperature were greater than those of the single correlation coefficients, indicating a similar trend to the results of the current study. High solar radiation and lower temperatures were the main factors contributing to the high flavonols content in October 2015. In the present study, no new leaves opened starting in early September because the mulberry trees had stopped growing. Consequently, the leaves sampled after late September had received more solar radiation than had those sampled in mid-September. This increase in the amount of solar radiation reaching the leaves sampled after late September is probably one of the reasons for their increased flavonols content.
Previous studies have focused on the relationship between flavonoid concentrations in grape berry skins and environmental conditions. Spayd et al. (2002) reported that temperature did not affect the flavonols concentration in grape berry skins. Mori et al. (2005) reported that anthocyanin contents decreased under high night temperatures, but flavonols contents did not. Although the relationship between flavonols content and temperature is not so clear, Azuma et al. (2012) reported that the flavonols contents in grape skins increased with low temperature and light in an in vitro experiment. Løvdal et al. (2010) reported that rutin contents in tomato leaves increased with low temperature in nitrogen-limited conditions in a growth chamber. Our results showed similar trends to those reported by Azuma et al. (2012) and Løvdal et al. (2010) (Fig. 2A). The average temperature from May to October 2015, when harvesting of mulberry leaves is possible, ranged from 15°C to 29°C (accumulated temperature for 14 days before sampling ranged from 234°C to 380°C). In our outdoor experiment, the accumulated temperature was negatively correlated with flavonols contents. As the primary factor was solar radiation, temperature was considered to be the second factor affecting flavonols contents.
The multiple correlation among total solar radiation hours, total temperature, and flavonols content was higher than the respective single correlations (Table 1), suggesting that solar radiation (positive correlation) and temperature (negative correlation) have synergistic effects on flavonols contents. That is, the increase in polyphenol contents after late September is due to the synergistic effect of solar radiation and decreasing temperatures, and the low flavonols contents in August are due to high solar radiation and the inhibitory effect of high temperatures. The reason for the lower flavonols contents in June than in October is likely the lack of a synergistic effect between solar radiation and temperature, although there was abundant solar radiation in June.
The seasonal changes in DNJ contents were strongly correlated with temperature. Correlation between the DNJ contents and total hours of solar radiation was not noted in the common culture field study data from May 28 to July 28, 2009 and May 24 to July 27 in 2010 (Sugiyama et al., 2016a). However, the correlation coefficients with total temperature in their study were high at r = 0.92 (P = 0.02) in 2009 and r = 0.80 (P = 0.04) in 2010, suggesting a similar trend to the present study (data not shown). Kimura et al. (2007) and Nakanishi et al. (2011) reported that the DNJ contents in leaves peaked in August, and were affected by temperature. In our study, the DNJ contents in mulberry leaves increased slightly from late May to mid-July, peaked in August, and then decreased sharply after late August. Konno (2011) found that most DNJ is in the latex, and that only small amounts of latex are produced in new shoots because of its immature laticifers. This may explain why there was only a slight increase in DNJ contents in mulberry leaves between May and June.
We further investigated the effect of temperature on DNJ contents using plants cultivated in growth chambers. The DNJ contents in leaves under high temperature (25–30°C) did not change, but that in leaves under low temperature (18–23°C) decreased (Fig. 3). The DNJ content in latex did not change under low temperature, but increased under high temperature. Thus, high temperature appeared to accelerate DNJ synthesis in latex. Therefore, to obtain leaves with high DNJ contents, leaves should be harvested while the shoots of mulberry trees are actively growing when they contain abundant latex.
Mulberry leaves are harvested once or twice per year as raw materials to produce edible products. The amount of total functional components per unit leaf area depends on the total leaf yield and the concentrations of functional components in leaves. Therefore, we evaluated the total amount of functional components per stock. The leaf yield of the once-harvested group was half that of the twice-harvested group. The new shoot length and the shoot length without leaves after defoliation of the once-harvested group were approximately three and 1.8 times longer than those of the twice-harvested group (October 26), respectively (Table 2). This was probably because the leaves located at a lower position on the shoot received a smaller amount of solar radiation, leading to an elongated portion of the shoot where leaves were lost. This phenomenon was considered to be one of the causes for the markedly smaller leaf yield in the once-harvested group than in the twice-harvested group. The flavonols contents per shoot was higher in the once-harvested group than in the twice-harvested group on the first harvest day (July 15), but the total amount of functional components per stock was higher in the twice-harvested group than in the once-harvested group (Table 3). These results confirmed that both leaf yield and the amount of functional components were higher with two harvests per season than with one harvest per season. This trend was strongest for DNJ, presumably because of the synergetic effect of low DNJ levels in leaves at lower temperature and the low leaf yield after a single harvest. Chlorogenic acid contents in mulberry leaves increased owing to trimming of shoots in the twice-harvested group (Fig. 2). These results indicate that harvesting twice is better than harvesting once a year, not only for the total functional component amount per unit area, but also for the functional component content in leaves.
Based on the results of the current study, hours of solar radiation and temperature appear to be valid indicators for determining the best harvest time to obtain the highest amounts of each targeted functional component. Although there was a wide deviation in total hours of solar radiation between the year of the current study (2015) and average yearly values, the trends in total hours of solar radiation and total temperature were similar (Fig. 1). Therefore, the harvest timing to obtain optimal functional component contents in 2015 should have been almost the same as that of normal years. Furthermore, to improve the accuracy of the estimated optimal harvest timing for obtaining high functional component contents, it is important to take into account yearly trends in solar radiation and temperature. Our results showed that with one harvest per year, the flavonols and chlorogenic acid contents in mulberry leaves were high after late September when the temperatures were lower, but there was still abundant solar radiation, and the DNJ levels were highest in August when the temperatures were high. Taking all of the functional components into account, August is a suitable time for harvesting because it represents a compromise among the contents of functional compounds. With two harvests per year, the first harvest should be in early summer (not later than early August) when the temperatures are high and solar radiation is abundant. The second harvest should be completed by the end of September, considering that shoot extension that affects the yield of mulberry leaves.