Abstract
Flower vase life of cut ornamental flowers, including carnations, is important in determining their quality and consumer preference for cut flowers. Therefore, we repeatedly crossed and selected promising offspring with long vase life for seven generations, from 1992 to 2008, in order to improve the vase life of carnation (Dianthus caryophyllus L.) flowers using conventional cross-breeding techniques. We investigated flower vase life, ethylene production at natural senescence, ethylene production after ethylene treatment (autocatalytic ethylene biosynthesis), response time to ethylene treatment (ethylene sensitivity), and flower diameter in six carnation cultivars used for initial breeding materials and a total of 123 selected lines from the first to seventh generations. Our results indicate that there was a large genetic variability in the five traits. Significant negative correlations were observed between flower vase life and ethylene production at natural senescence (r = −0.88**), between flower vase life and ethylene production after ethylene treatment (r = −0.90**), and between flower vase life and flower diameter (r = −0.92**). However, there was no correlation between flower vase life and ethylene sensitivity. Thus, the long vase life of selected carnation lines was strongly associated with a decrease in their ethylene production. Our results indicate that variation in flower vase life is not due to differences in ethylene sensitivity, but due to differences in ethylene production. Although flower size is an important floricultural trait for commercial production, crossing and selection for flower vase life resulted in a considerable reduction in flower diameter. The course of mean flower vase life over generations showed that selection was effectively made in the first to sixth generations. Repeatability for the flower vase life was roughly constant (0.3 to 0.4) in the first to sixth generations, but decreased to 0.12 in the seventh generation, suggesting a decrease in genetic variation in the seventh generation.
Introduction
The carnation (Dianthus caryophyllus L.) is one of the most commercially important floricultural crops in Japan and around the world. Flower vase life of cut ornamental flowers, including carnations, is one of the most important breeding targets determining their quality and ability to satisfy consumer preferences, thus stimulating repeated purchases. Therefore, we started a breeding research program in 1992 to improve the vase life of carnation flowers using conventional cross-breeding techniques (Onozaki et al., 2001).
Heritability is one of the most fundamental concepts in quantitative genetics (Falconer, 1960). Broad-sense heritability (H2) is defined as the ratio of genetic variance to total variance (phenotypic variance), that is H2 = VG/{VG + (VE/r)}, where VG, VE, and r are the genotypic variance, environmental variance, and number of replicates, respectively (Goncalves et al., 2013; Van der Meulen-Muisers et al., 1999). It is effective to estimate the H2 to determine whether the breeder has made, or will be able to make, an effective selection (Nishio et al., 2014). Selecting for characteristics with high broad-sense heritability will lead to faster and greater gains in the offspring than selecting for traits with low heritability (Browning et al., 1994). H2 takes values in the range from 0 to 1. If H2 is zero, all observed differences in target trait are due to environmental factors, any selection is meaningless, and the breeder cannot select superior genotypes. If H2 is 1, all observed differences among phenotypic values are due to genetic differences. Therefore, breeders should concentrate on generational differences of heritability in target traits (Nishio et al., 2014).
Breeding for postharvest performance in ornamentals is challenging, since many different deteriorative processes determine vase life (Van Geest et al., 2017). As flower vase life is strongly influenced by environmental factors, it is important to estimate the heritability which reflects all the genetic contributions to a population’s phenotypic variance. The breeders usually make several crosses, and primarily select offspring individuals with no replicates, yielded from the crosses, based on their performance in terms of vase life, and make crosses among the selected offspring. The performance in each offspring individual consists of a genotypic effect and the sum of environmental effects specific to the individual and flowers evaluated. The environmental effect specific to an individual can be estimated only by using replicates of an offspring individual with the same genotypic value. It is too costly and laborious for breeders to make selection for multiple individuals within offspring after propagating offspring in each generation, resulting in an estimate of broad-sense heritability. In contrast, the environmental effect specific to flowers evaluated can be easily estimated because multiple flowers are usually evaluated for each offspring individual.
Falconer (1960) divided VE into VEg (environmental variance between individuals) and VEs (environmental variance within individual), and showed repeatability, (VG + VEg)/VP, rather than heritability. In the present study, we estimate the repeatability instead of the broad-sense heritability, and the change in genetic variance over generations assuming unchanged VEg and VEs with the same culture conditions over generations.
We repeatedly crossed and selected promising offspring with long vase life for seven generations, from 1992 to 2008, in order to improve the vase life of carnation flowers (Onozaki et al., 2001, 2006b, 2011). In previous papers, we described the development of many carnation lines with increased vase life as a result of reduced ethylene biosynthesis achieved by crossing and selection for seven generations. Two cultivars, ‘Miracle Rouge’ and ‘Miracle Symphony’, with a genetically determined long vase life of 17.7 to 20.7 days under standard conditions (23°C and 70% RH under a 12-h photoperiod), were developed in 2005 (Onozaki et al., 2006a). Their flowers produce a very small amount of ethylene during senescence (Onozaki et al., 2006a), and their long vase life is correlated with the low expression of 1-aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase genes (DC-ACS1, DC-ACS2, and DC-ACO1) in the gynoecium and petals (Tanase et al., 2008). Furthermore, we developed two ultra-long-vase-life lines, 532-6 and 806-46b, with a flower vase life of over 27 days under standard conditions (23°C and 70% RH under a 12-h photoperiod) (Onozaki et al., 2011, 2015). The ultra-long vase life of line 532-6 may be controlled by DcCPIn and DcCP1, which are senescence-related genes in carnations (Tanase et al., 2015).
However, characteristic alterations by repeated crossing and selection for long vase life, in other words, variation in traits with long vase life, were poorly studied. Therefore, we investigated the relationship between ethylene production at natural senescence, ethylene production after ethylene treatment, response time to ethylene treatment, flower diameter, and flower vase life using a total of 123 selected lines spanning seven generations.
Materials and Methods
Crossing and selection
We chose six commercial standard carnation (D. caryophyllus) cultivars with large differences in vase life as initial breeding materials: four Mediterranean-type cultivars (‘Pallas’, ‘Sandrosa’, ‘Candy’, and ‘Tanga’) and two American Sim-type cultivars (‘White Sim’ and ‘Scania’) (Onozaki et al., 2001). In spring 1992, crosses were made among these cultivars. We then crossed and selected promising offspring with long vase life for seven generations, from 1992 to 2008, in order to improve the vase life of carnation flowers. The populations that we used in the present study were the same offspring populations over generations as reported previously (Onozaki et al., 2001, 2006b, 2011). Here we use the term “first generation”, “second generation”, “third generation”, “fourth generation”, “fifth generation”, “sixth generation”, and “seventh generation” in place of “parental generation”, “first generation”, “second generation”, “third generation”, “fourth generation”, “fifth generation”, and “sixth generation” described in the previous papers. In previous studies, we described the detailed breeding process (Onozaki et al., 2001, 2006b, 2011).
Six parental cultivars, 9 lines selected from the first generation, 15 lines selected from the second generation, 20 lines selected from the third generation, 17 lines selected from the fourth generation, 29 lines selected from the fifth generation, 19 lines selected from the sixth generation, and 14 lines selected from the seventh generation were used for five traits evaluation (Table 1).
Measurement of flower diameter and vase life evaluation
Carnation cultivars and lines were grown in a greenhouse following standard production methods for carnations (Onozaki et al., 2006a) and harvested at the flower-opening stage (i.e., when the outer petals became orientated at right angles to the stem). Stems of the freshly harvested flowers were trimmed to 50 cm, and the two or three lowest pairs of leaves were removed. After the adjustment, the diameter of each harvested flower was measured with a ruler. The flowers were then placed in 4-L jars containing ca. 1 L of distilled water. The water was replaced every 3 or 4 days.
The vase life of each flower was determined by the number of days from harvesting until the petals showed in-rolling, browning, loss of surface turgor, or other morphological changes detrimental to their ornamental value. Flowers were evaluated daily in an inspection room, which was kept at 23°C, 70% RH, and 12-h photoperiod (08:00 to 20:00 h) provided by cool fluorescent lamps (10 μmol·m−2·s−1 irradiance).
In the seedling trials in the first year of each generation, all harvested flowers were used for vase life evaluation and individual flower vase life data was used for estimates of repeatability.
Regarding the clonal line test in the second year, as a general rule, 10 flowers were harvested for evaluation in the selected line of each generation, and the mean vase life in each line was determined for secondary-selection.
Measurement of ethylene production at natural senescence and after ethylene treatment
Ethylene measurements at natural senescence were carried out when senescence symptoms first appeared. Flowers were harvested at the stage of flower opening (i.e., when the outer petals became orientated at right angles to the stem) and their stems were trimmed to 5 cm and placed individually in a test tube containing distilled water. Flowers were kept under standard conditions (23°C, 70% RH, and 12-h photoperiod) until petals showed in-rolling, browning, or loss of surface turgor and ornamental value. When senescence was first observed, individual flowers were weighed (fresh weight), and then put in a glass jar and kept at 23°C. After incubation for 1 or 2 h, a 0.5-mL sample of the headspace gas was withdrawn and injected into a gas chromatograph (model GC-7A or GC-13A; Shimadzu, Kyoto, Japan) equipped with an alumina column and a flame ionization detector to determine ethylene concentration.
Measurements of ethylene production after ethylene treatment (autocatalytic ethylene biosynthesis) were carried out using flowers on day 0 treated with ethylene at 2 μL·L−1 for 16 h at 23 ± 1°C in a 50-L chamber. After ethylene treatment, flowers were left for 8 h at 23°C and 70% RH and exposed to fresh air. The flowers were then individually enclosed in a glass jar and held at 23°C. After 1-h incubation, each gas sample was analysed for ethylene concentration as described above.
Video evaluation of ethylene sensitivity
Ethylene sensitivity was measured using a time-lapse video recording system at 10 μL·L−1 ethylene concentration and 23 ± 1°C in a 50-L acryl chamber, as described previously (Onozaki et al., 2004). On day 0, the stems of freshly harvested flowers were trimmed to 20 cm and kept individually in 100-mL Erlenmeyer flasks containing distilled water. The relative humidity in the chamber was kept between 40% and 60% by including about 100 g of silica gel. The time to onset of petal in-rolling, referred to as the ‘response time’ hereafter, was determined by fast-forwarding the image data files.
Statistical analyses
We performed analysis of variance (ANOVA) for individual flower vase life of offspring in each generation using the following model:
|
X
i
j
=
μ
+
(
G
+
E
g
)
i
+
E
s
i
j
|
where
Xij is the observed value of the
jth flower of the
ith offspring (genotype),
μ is the overall mean, (
G+
Eg)
i is the genetic and environmental between-individual effect of the
ith offspring (genotype), and
Esij is the environmental within-individual effect of the
jth flower of the
ith offspring (genotype).
The mean was correlated with standard deviation for the vase life (days) in offspring; therefore, we used a log-transformed value of the vase life days for ANOVA. The heterogeneity of variances in offspring was not significant by Cochran’s test for the log-transformed data of offspring chosen with a given constant number (four or six) of flowers evaluated for vase life in each generation, indicating that the ANOVA model is applicable. ANOVA provided the genetic plus and environmental between-individual variance component (VG + VEg) and the environmental within-individual variance component (VEs).
Repeatability (R) for the flower vase life was estimated as R = (VG + VEg)/(VG + VEg + VEs). As Falconer (1960) showed, accuracy is measured by the “(VG + VEg)/{VG + VEg + (VEs/n)}” value as the gain in accuracy, where n is the number of repeating measurements. “n” should be a constant, but, n was not constant depending on the offspring in each generation in the present study. Here, to monitor the change in the gain in accuracy over generations approximately, we used the average number of flowers evaluated for vase life in the offspring as “n” in each generation.
Results
Crossing and selection
The mean flower vase life of individual seedlings in each generation increased slowly and steadily as generations progressed (Fig. 1). The mean vase life was 7.4 days for the first generation derived from crossing six parental cultivars; in contrast, after six cycles of crossing and selection, this was improved to 15.9 days, resulting in a net increase of 8.5 days (Fig. 1). The effect of crossing and selection did not stay constant, but varied according to generations. The biggest genetic improvement for vase life was observed between the fourth and fifth generations. The population mean for vase life between them increased by 4.2 days.
Changes in five traits by repeated selection and crossing for vase life
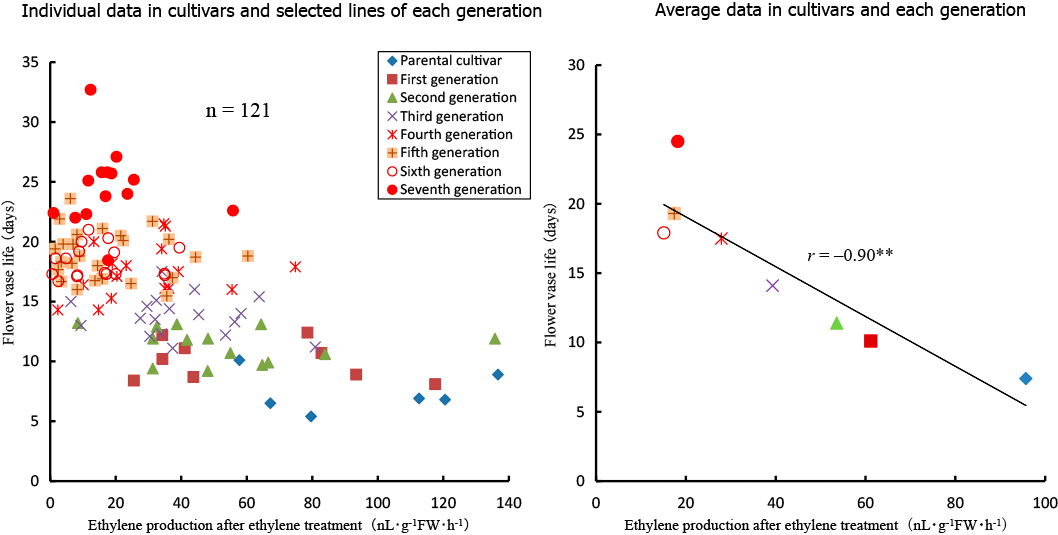
Six cultivars and our selected lines showed great variation in their flower vase life, ethylene production at natural senescence, ethylene production after ethylene treatment, response time to ethylene treatment, and flower diameter (Figs. 2, 3, 4, and 5).
Ethylene production at natural senescence markedly decreased from the first to seventh generations (Fig. 2). In particular, the selected lines in the fourth to seventh generations produced a very small amount of ethylene without exception. Average data of each generation showed a significant negative correlation (r = −0.88**) between ethylene production at natural senescence and flower vase life.
Ethylene production after ethylene treatment showed the same tendency of ethylene production at natural senescence (Fig. 3). A significant negative correlation (r = −0.90**) was observed between ethylene production after ethylene treatment and flower vase life. Ethylene production after ethylene treatment of selected lines decreased gradually from the first to fifth generation. The average ethylene production after ethylene treatment of the fifth, sixth, and seventh generations was approximately the same (17.4, 15.1, and 18.2 nL·g−1FW·h−1, respectively).
On the other hand, there was no correlation between response time to ethylene treatment and flower vase life (Fig. 4). All cultivars and almost all lines showed response times from 6 to 16 h, which were classified as sensitive or moderately sensitive based on our criteria for the degree of ethylene sensitivity (Onozaki et al., 2008). Only one selected third-generation line (64-54) and two selected sixth-generation lines (203-42S and 204-41S) showed ethylene resistance, with a response time of more than 16 h (Fig. 4).
Although flower size is an important floricultural trait for commercial production, crossing and selection for flower vase life resulted in a considerable reduction in flower diameter. Although six parental cultivars had large size flowers (6.0–7.5 cm), the flower diameter of selected lines with long vase life tended to decrease gradually from the first to seventh generations (Fig. 5). Mean flower diameter in the first generation was 6.4 cm, and that in the seventh generation was 5.6 cm (Table 2). A significant negative correlation (r = −0.92**) was observed between flower diameter and flower vase life (Fig. 5).
Changes in repeatability over generations for flower vase life
Repeatability was roughly constant (0.3 to 0.4) in the first to sixth generation; however, it markedly decreased to 0.12 in the seventh generation (Table 3). The effect of between-offspring was not significant only in the seventh generation in agreement with the above. These results suggest a marked decrease in the genetic variance in the seventh generation, assuming unchanged environmental variation over generations due to unchanged culture and flower vase evaluation conditions over generations.

The number of average flowers per offspring individual gradually decreased over generations, from around three in the first to third generations, to around 1.5 in the sixth to seventh generations (Table 3), suggesting a decrease in vigor over generations, which is generally observed in heterozygous crops with a decrease in heterozygosity after repeated selection for genetically additive controlled traits. The selection was based on the average flower vase life per offspring, so the decrease in the number of flowers evaluated per offspring led to the decrease in efficiency of selection. Consequently, the gain in accuracy, which is measured by “(VG + VEg)/{VG + VEg + (VEs/n)}” value (Falconer, 1960), was roughly constant (around 0.6) from the first to fifth generations, decreased to 0.4 in the sixth generation, and markedly decreased to 0.19 in the seventh generation. The decrease in the seventh generation was estimated to be caused by both the decrease in genetic variance and the number of flowers evaluated per offspring.
Overall, these results indicate that effective selection was achieved from the first to sixth generations, which had considerable genetic variation, irrespective of the large environmental variance for the flower vase life.
Discussion
We conducted a long-term carnation breeding study of crossing and selection for seven generations based on flower vase life from 1992 to 2008. In this process, we selected lines with a long vase life in each generation for use as breeding materials for the next generation. Plants were not selected for ethylene production or sensitivity, but on the vase life of flowers. We selected a total of 123 lines (including 2 cultivars, ‘Miracle Rouge’ and ‘Miracle Symphony’) with improved vase life by conventional breeding techniques (Table 1). The crossing and selection resulted in dramatic alterations in the flower vase life of each generation and particularly in ethylene production of many selected lines (Onozaki et al., 2001, 2006b, 2011).
In this study, we evaluated 6 cultivars and 123 selected lines for five traits. There was a wide genetic variability in flower vase life, ethylene production at natural senescence (Fig. 2), ethylene production after ethylene treatment (autocatalytic ethylene biosynthesis) (Fig. 3), response time to ethylene treatment (ethylene sensitivity) (Fig. 4), and flower diameter (Fig. 5). Significant negative correlations were observed between flower vase life and ethylene production at natural senescence, between flower vase life and autocatalytic ethylene biosynthesis, and between flower vase life and flower diameter (Figs. 2, 3, and 5).
On the other hand, there was no relationship between ethylene sensitivity and flower vase life. In previous studies, we selected a total of 13 ethylene-resistant lines (Onozaki et al., 2008, 2011) by a video evaluation method (Onozaki et al., 2004). However, none of lines had an extremely long vase life in comparison with our two long-vase-life cultivars (‘Miracle Rouge’ and ‘Miracle Symphony’) or other selected lines with extremely low ethylene production, such as line 532-6 (Onozaki et al., 2008, 2011). Flowers of line 532-6 had the longest vase life (32.7 days) among a total of 129 cultivars and lines despite having high ethylene sensitivity with a response time of 6.6 h (Fig. 4).
Furthermore, we succeeded in developing line 806-46b with both ultra-long-vase-life and ethylene resistance by conventional cross-breeding techniques (Onozaki et al., 2015), but genes conferring an ultra-long vase life seemed to correlate poorly with ethylene sensitivity (Onozaki et al., 2015). These results strongly imply that crossing and selection based on only flower vase life is ineffective for breeding ethylene-resistant lines.
In another previous paper, we investigated longevity, ethylene production, and ethylene sensitivity of flowers among 42 pot carnation cultivars (Onozaki et al., 2009). Although significant negative correlations were observed between flower longevity and ethylene production at natural senescence, and between flower longevity and autocatalytic ethylene production, there was no correlation between flower longevity and ethylene sensitivity (Onozaki et al., 2009). Our results here and in previous studies indicate that variation in flower vase life is not due to differences in ethylene sensitivity, but to differences in ethylene production. Similar results were reported by Ebrahimzadeh et al. (2011) using ten Mediterranean carnation cultivars.
Our investigation revealed a significant negative correlation between flower diameter and flower vase life (Fig. 5). As flower diameter is an important characteristic in cut carnations, its extreme reduction is undesirable for commercial cultivars, especially the standard type (one flower per stem). To develop commercial cultivars with both long vase life and large flowers, we need to carry out selection and crossing with consideration of flower diameter.
However, as this significant negative correlation between them might potentially relate to inbreeding depression, further studies are needed to clarify the relationship between flower diameter and flower vase life. Carnation cultivars are developed to be highly heterozygous so as to avoid the effects of inbreeding depression. Sato et al. (2000) reported that inbreeding depression of carnation appears in the third selfed generation (S3), so it is almost impossible to produce S4 seeds.
Another way to overcome this problem is to breed spray-type cultivars (multi-flowers per stem) with a long vase life. In spray-type cultivars, a relatively small flower size is acceptable for commercial cultivars. From this view point, we have conducted joint research on carnation breeding with the Aichi Agricultural Research Center since 2006, and developed ‘Kane Ainou 1-go’, a new spray-type carnation cultivar with a long vase life, using our selected fifth-generation line 108-44 for breeding material, in 2015 (Hotta et al., 2016).
Estimates for broad-sense heritability of flower vase life have been previously reported in several cut ornamental flowers. Broad-sense heritability estimates were found to be 0.36 to 0.46 (Harding et al., 1981) and 0.28 (Wernett et al., 1996) in Gerbera × hybrida, and 0.79 in Lilium (Van der Meulen-Muisers et al., 1999).
In our study, the gain in accuracy measured by the “(VG + VEg)/{VG + VEg + (VEs/n)}” value for vase life of carnations in the first to fifth generations was around 0.6, higher than the heritability of Gerbera × hybrida (Harding et al., 1981; Wernett et al., 1996), which was estimated as VG/{VG + (VE/n)}, possibly due to a considerable VEg in the present study. The course of mean flower vase life over generations is shown in Figure 1 and these estimates showed that selection was effectively made from the first to sixth generations.
Moreover, these findings raise the possibility that the decrease in genetic variance for vase life in later generations was related with a rapid decrease in their genetic capacity to produce ethylene, which had a large effect on the vase life of carnations (Figs. 2 and 3). In particular, ethylene production at natural senescence in the sixth and seventh generations was extremely low (average ethylene production levels at natural senescence were 1.1 and 0.9 nL·g−1FW·h−1, respectively, as shown in Table 2) and there was little difference in that among 17 and 14 selected lines in the sixth and seventh generations (Fig. 2).
In conclusion, we demonstrated a close correlation between flower vase life and ethylene production in our nearly two decades-long breeding research on cut carnations. On the other hands, no significant correlation was found between flower vase life and ethylene sensitivity. Repeatability was roughly constant (0.3 to 0.4) from the first to sixth generation; however, it markedly decreased to 0.12 in the seventh generation, suggesting a marked decrease in the genetic variance in the seventh generation. The course of mean flower vase life over generations was from the first generation with 7.4 days to the seventh generation with 15.9 days (Fig. 1); these estimates showed that selection was effectively made from the first to sixth generations, which had considerable genetic variation, irrespective of a large environmental variance for flower vase life.
Literature Cited
- Browning, S. J., T. P. Riordan, R. K. Johnson and J. Johnson-Cicalese. 1994. Heritability estimates of turf-type characteristics in buffalograss. HortScience 29: 204–205.
- Ebrahimzadeh, A., S. Jimenez-Becker, S. Manzano-Medina, M. Jamilena-Quesada and M. T. Lao-Arenas. 2011. Evaluation of ethylene production by ten Mediterranean carnation cultivars and their response to ethylene exposure. Span. J. Agric. Res. 9: 524–530.
- Falconer, D. S. 1960. Introduction to quantitative genetics. Oliver and Boyd, London.
- Goncalves, E., I. Carrasquinho, A. St. Aubyn and A. Martins. 2013. Broad-sense heritability in mixed models for grapevine initial selection trials. Euphytica 189: 379–391.
- Harding, J., T. Byrne and R. L. Nelson. 1981. Heritability of cut-flower vase longevity in gerbera. Euphytica 30: 653–657.
- Hotta, M., H. Hattori, T. Hirano, T. Kume, Y. Okumura, K. Inubushi, Y. Inayoshi, M. Ninura, J. Matsuno, T. Onozaki, M. Yagi, H. Yamaguchi and N. Yamaguchi. 2016. Breeding and characteristics of spray-type carnation ‘Kaneainou 1 go’ with a long vase life. Res. Bull. Aichi Agric. Res. Ctr. 48: 63–71 (In Japanese with English abstract).
- Nishio, S., M. Yamada, N. Takada, H. Kato, N. Onoue, Y. Sawamura and T. Saito. 2014. Environmental variance and broad-sense heritability of nut traits in Japanese chestnut breeding. HortScience 49: 696–700.
- Onozaki, T., H. Ikeda and M. Shibata. 2004. Video evaluation of ethylene sensitivity after anthesis in carnation (Dianthus caryophyllus L.) flowers. Sci. Hortic. 99: 187–197.
- Onozaki, T., H. Ikeda, M. Shibata, N. Tanikawa, M. Yagi, T. Yamaguchi and M. Amano. 2006a. Breeding and characteristics of carnation Norin No. 1 ‘Miracle Rouge’ and No. 2 ‘Miracle Symphony’ with long vase life. Bull. Natl. Inst. Flor. Sci. 5: 1–16 (In Japanese with English abstract).
- Onozaki, T., H. Ikeda and T. Yamaguchi. 2001. Genetic improvement of vase life of carnation flowers by crossing and selection. Sci. Hortic. 87: 107–120.
- Onozaki, T., N. Tanikawa, M. Yagi, H. Ikeda, K. Sumitomo and M. Shibata. 2006b. Breeding of carnations (Dianthus caryophyllus L.) for long vase life and rapid decrease in ethylene sensitivity of flowers after anthesis. J. Japan. Soc. Hort. Sci. 75: 256–263.
- Onozaki, T., M. Yagi and M. Shibata. 2008. Selection of ethylene-resistant carnations (Dianthus caryophyllus L.) by video recording system and their response to ethylene. Sci. Hortic. 116: 205–212.
- Onozaki, T., M. Yagi and K. Tanase. 2009. Genetic variation in the longevity, ethylene production and ethylene sensitivity of flowers among pot carnation cultivars. Hort. Res. (Japan) 8: 399–405 (In Japanese with English abstract).
- Onozaki, T., M. Yagi and K. Tanase. 2015. Selection of carnation line 806-46b with both ultra-long vase life and ethylene resistance. Hort. J. 84: 58–68.
- Onozaki, T., M. Yagi, K. Tanase and M. Shibata. 2011. Crossings and selections for six generations based on flower vase life create lines with ethylene resistance or ultra-long vase life in carnations (Dianthus caryophyllus L.). J. Japan. Soc. Hort. Sci. 80: 486–498.
- Sato, S., N. Katoh, H. Yoshida, S. Iwai and M. Hagimori. 2000. Production of doubled haploid plants of carnation (Dianthus caryophyllus L.) by pseudofertilized ovule culture. Sci. Hortic. 83: 301–310.
- Tanase, K., T. Onozaki, S. Satoh, M. Shibata and K. Ichimura. 2008. Differential expression levels of ethylene biosynthetic pathway genes during senescence of long-lived carnation cultivars. Postharvest Biol. Technol. 47: 210–217.
- Tanase, K., S. Otsu, S. Satoh and T. Onozaki. 2015. Expression levels of ethylene biosynthetic genes and senescence-related genes in carnation (Dianthus caryophyllus L.) with ultra-long-life flowers. Sci. Hortic. 183: 31–38.
- Van der Meulen-Muisers, J. J. M., J. C. van Oeveren, J. Jansen and J. M. van Tuyl. 1999. Genetic analysis of postharvest flower longevity in Asiatic hybrid lilies. Euphytica 107: 149–157.
- Van Geest, G., A. Post, P. Arens, R. G. F. Visser and U. van Meeteren. 2017. Breeding for postharvest performance in chrysanthemum by selection against storage-induced degreening of disk florets. Postharvest Biol. Technol. 124: 45–53.
- Wernett, H. C., G. J. Wilfret, T. J. Sheehan, P. M. Lyrene, F. G. Martin, T. L. White, G. L. Powell and C. J. Wilcox. 1996. Postharvest longevity of cut-flower Gerbera. 2. Heritability of vase life. J. Amer. Soc. Hort. Sci. 121: 222–224.