Abstract
Uniform temperature throughout a greenhouse is recommended, as the present climate control method and many other studies have shown that the temperature gradient decreases vertically and horizontally in a greenhouse. However, recent research revealed that roots, fruits, flowers, and shoot-tips are more sensitive to temperature changes than leaves and stems, indicating that uniform temperature control may not be necessary. In addition, energy-saving techniques that do not lead to yield loss are desirable to reduce energy costs and ensure sustainable greenhouse production. In this paper, we review current studies on local temperature control methods in greenhouse vegetable production, primarily focusing on the tomato, and compare them with novel climate-control techniques. Roots, fruits, shoot-tips, and flowers are sensitive to temperature changes, showing negative symptoms under extreme temperature conditions. Therefore, the temperature of these plant organs should be controlled locally. Root zone temperature control enhances root growth and its associated physiological activities, promoting uptake of water and mineral nutrients. This subsequently leads to enhanced growth of shoots. Fruit temperature control may not be effective for tomato plants, but it promotes fruit growth and fruit sugar accumulation in melons and watermelons. Shoot-tip temperature control promotes the differentiation of leaf and flower buds. Flower temperature control enhances pollen viability and promotes fruit set. The combination of shoot-tip and flower heating enables low energy consumption compared with conventional heating, without loss of yield. Local temperature control techniques (except roots) have been studied in recent years; however, there is a distinct lack of research on the physiological mechanisms and practical approaches to develop a better local temperature control system. Thus, further studies are required on this area in the future.
Introduction
In greenhouse vegetable production, temperature can be controlled easily compared with production in open fields. Using greenhouses with thermal screen and heating systems is important in securing winter vegetable production. Additionally, in recent years, greenhouses have been using cooling systems for summer production, which enable year-round greenhouse production. These techniques are effective for maximizing employee utility and ensuring the best possible productivity in greenhouses because high temperatures in summer often interrupt vegetable production in some regions. Since temperature control techniques are costly, it is important to research ways in which costs can be minimized and the effects of temperature control can be maximized. Reducing energy expenditure simultaneously reduces carbon dioxide emissions and contributes to sustainable vegetable production.
The study of temperature control in greenhouses started in the 1940s (Went, 1943, 1944). After that, more studies followed, and the optimum temperature for plant growth and the temperatures at which high and low temperature injury occur were also demonstrated. Tomato plants (Solanum lycopersicum L.), for example, grow well at a daily mean temperature of 15–25°C, but are damaged under suboptimal temperature conditions, differing according to cultivar and other environmental conditions such as solar radiation (Table 1). Flowers (Peet et al., 1997; Sato et al., 2000, 2002) are the most temperature-sensitive organs in tomato plants, followed by roots (Gosselin and Trudel, 1983a, b), shoot-tips (Grimstad and Frimanslund, 1993), and developing fruits (Adams and Valdés, 2002; Adams et al., 2001; Bertin, 2005), which are also relatively sensitive. On the other hand, severe injuries to the leaves and stems rarely occur earlier than those in the other organs.

Horizontal and vertical temperature differences in greenhouses should be reduced and many temperature control methods have been tested to achieve a uniform temperature by changing the position of the heating pipes or by using a circulator (Fernandez and Bailey, 1994; Jeff and Jones, 1973; Kuroyanagi, 2016; Teitel and Tanny, 1998). However, having a temperature is not absolutely essential for adequate plant growth, as roots and flowers (the most temperature-sensitive organs) only occupy a small space in the greenhouse. Therefore, local temperature control techniques based on the physiological temperature responses of each plant organ, are important to ensure adequate plant growth. These are novel temperature control techniques, by which temperature-sensitive organs are controlled aggressively, whereas non-temperature-sensitive organs do not need to be controlled aggressively. Since these techniques do not require temperature control uniformly throughout the entire greenhouse, reduced energy expenditure can be expected. In addition, less temperature injury will occur as these techniques ensure that temperature-sensitive organs are maintained at an optimal temperature.
Heating is used as temperature control in winter, thereby increasing energy costs due to greater fuel use, which is highly detrimental to farmers. The application of chilling-tolerant cultivars (van der Ploeg et al., 2007), solar heat collection (Fang et al., 2015; Joudi and Farhan, 2014), heat-pumps (Tong et al., 2012; Zhou et al., 2016), thermal screens (Giacomelli and Roberts, 1993; Kawashima, 2015), water thermal screen systems (Ogura and Mukai, 1988), and split-night temperature (Gent et al., 1979; Toki, 1970) have been proposed as methods for saving energy. Local heating is also an efficient energy saving technique. Although many studies have been conducted on root-zone heating, there have been few and only relatively recent studies regarding the effects of local heating on other organs. Root-zone temperature can be easily controlled and can be distinguished from above-ground temperature, whereas local temperature control on above-ground organs is difficult.
Cooling is used as temperature control in the summer, using fogging (Handarto et al., 2006, 2007) and fan-and-pad (Al-Mulla et al., 2018; Lopez et al., 2010) systems. These systems operate with low energy costs as they use heat from water vaporization, but they have more difficulty to control temperature and humidity adequately. Although heat-pumps are used when accurate temperature control is needed, their use is not recommended during the day due to the huge energy costs required. Local cooling techniques could balance between accurate temperature control and low energy costs; however, little information is available about these techniques compared to local heating techniques. Even so, studies on root-zone cooling methods have been reported and some of them are in commercial use. On the other hand, few studies on local cooling, except on the root-zone, have been reported. However, these techniques will play a more important role in the future, as high temperature injury is predicted to occur more frequently in low latitude areas due to the impact of global warming.
In this review, we discuss current studies on local temperature control methods in greenhouse vegetable production, primarily focused on the tomato. We also compare them with novel climate control techniques.
Local Temperature Control of Roots
Root-zone temperature conditions differ from those above-ground and the study of plant reactions to root-zone temperature has a long history (Kramer, 1934). Because root-zone temperatures can be easily distinguished from above-ground temperatures, local temperature control of the root-zone has also been studied for a long time. The oldest studies focused on mineral absorption and vegetative growth in tomato plants under various root-zone temperature conditions in greenhouses (Davis and Lingle, 1961; Lingle and Davis, 1959). Since then, many studies on root-zone temperature control have been reported, with most of these focusing on maximizing root activity and shoot growth under optimal air temperature (Fujishige et al., 1991; Gosselin and Trudel, 1982, 1983a, b, 1984, 1985; Hurewitz and Janes, 1983; Ikeda and Osawa, 1988; Kabu and Toop, 1970; Tindall et al., 1990; Yelle et al., 1987). However, few studies have investigated root-zone temperature control to save energy or to alleviate temperature injury under suboptimal air temperature conditions. When the root-zone temperature is kept at an optimal level under suboptimal air temperature conditions, the balance of plant growth regulators changes, causing enhanced root growth and activities, accelerated water and mineral nutrient uptake, enhanced shoot growth and activity (e.g., photosynthesis), and finally increased yield (Fig. 1).
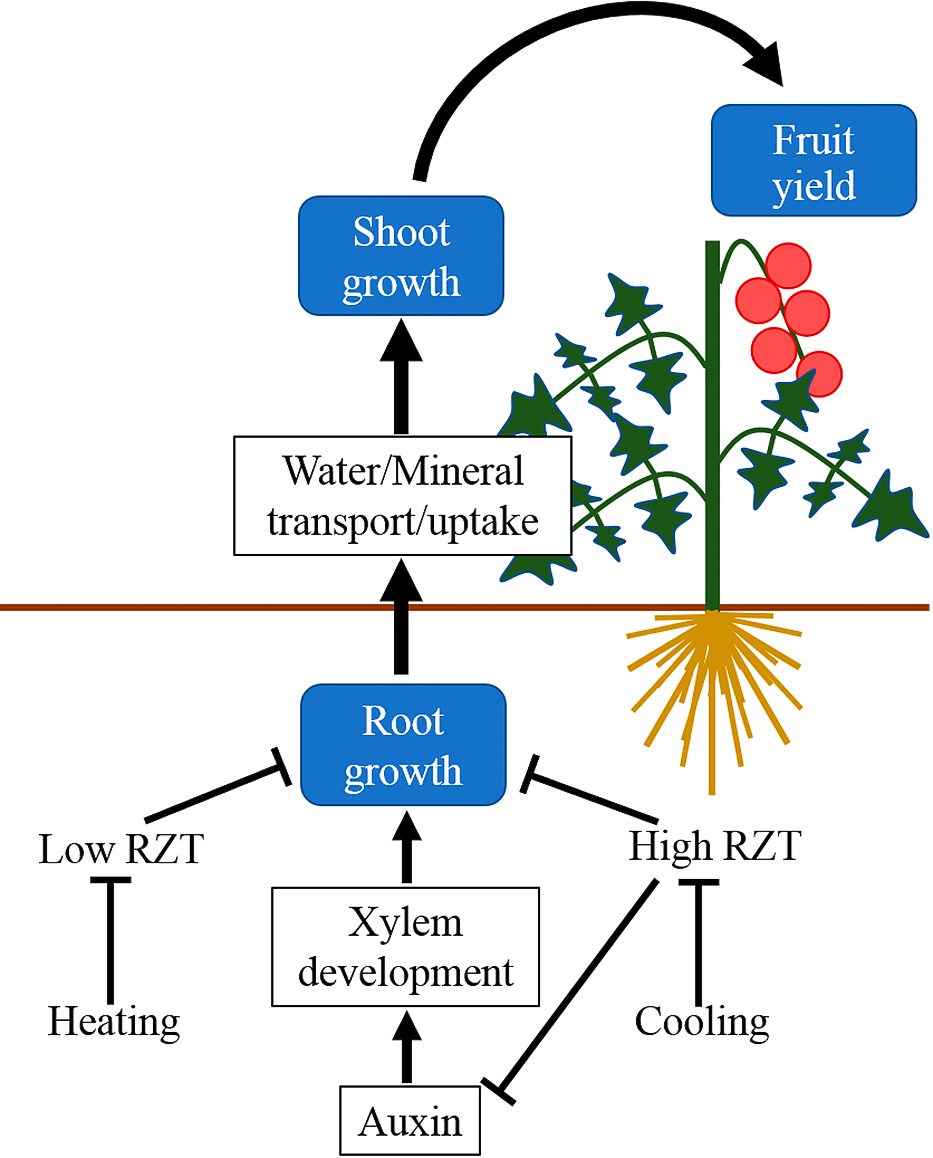
An optimum root-zone temperature increases the distribution of assimilates to the roots and promotes root growth (Shishido and Kumakura, 1994). In tomato plants, root growth was promoted earlier than shoot growth when the root-zone was cooled from 33.7°C to 24.6°C under high air temperature conditions in summer (30.8°C; average daily mean temperature) (Kawasaki et al., 2013). A similar effect was observed when the root-zone was heated from 5.8°C to 16.6°C under low air temperature conditions in winter (5.9°C; average daily minimum night temperature) (Kawasaki et al., 2014). In cucumber plants (Cucumis sativus L.), root weight was increased when soil temperature 15 cm below the ground surface was cooled from 26.9°C to 20.6°C under an average maximum air temperature of 39.2°C (Moon et al., 2007b).
Subsequently, uptake of water and mineral nutrients are accelerated with high root activity. In tomato plants, root-zone cooling from 33°C to 17°C at an air temperature of 35/22°C (day/night) increased water and nitrogen absorption, as well as enhancing xylem exudation (Nakano, 2007). Similar effects were also reported by root-zone heating and indicated that mineral nutrient transport to shoots was accelerated by xylem vessels (Kawasaki et al., 2014). These effects resulted in an increased mineral nutrient content in shoots (Gent and Ma, 2000; Klock et al., 1997; Nkansah and Ito, 1995b; Papadopoulos and Tiessen, 1987). In cucumber plants, heating the root-zone to 19°C from 13°C increased xylem exudation at 20/12°C (day/night) air temperature (Wang et al., 2016).
Shoot growth is promoted by enhanced shoot activities. In tomato plants, photosynthesis, stomatal conductance, and shoot weight were maximized by cooling the root-zone to 25°C at an air temperature of 40/23°C (day/night) (Nkansah and Ito, 1994, 1995a). Some similar studies have been reported (Kawasaki et al., 2013; Klock et al., 1997; Sasaki and Itagi, 1989). Around 20°C of root-zone heating increased leaf area, plant height, and relative growth rate (RGR) at low air temperature (16.2°C of daily mean temperature) (Kawasaki et al., 2014). In cucumber and tomato plants, the photosynthesis rate, stomatal conductance and shoot RGR increased with root-zone heating from 13°C to 19°C at an air temperature of 20/12°C (Wang et al., 2016). In the sweet pepper (Capsicum annuum L.), an increase in stomatal conductance and a decrease in leaf water potential were also observed, while the leaf area and dry weight of the whole plant increased, when the root-zone was cooled from 40°C to 20°C at an average daily maximum air temperature of 37.0°C (Dodd et al., 2000). In the lettuce (Lactuca sativa L.), root-zone cooling increased photosynthetic activities (He et al., 2001) and shoot dry weight (Ilahi et al., 2017).
These effects all ultimately lead to an increased yield. Chong and Ito (1982) reported that the total and marketable fruit yield of tomatoes were maximized when the root-zone temperature was kept at 20°C under a maximum air temperature exceeding 35°C; this had the added effect of reducing the occurrence of blossom-end-rot and abnormal fruits. However, some studies did not observe an increase in fruit yield, indicating that the effect of root-zone cooling on yield also depends on above-ground conditions (Gent and Ma, 1998; Papadopoulos and Tiessen, 1987). Root-zone heating at low air temperatures increased fruit yield and quality (Hurd and Graves, 1985; Jones et al., 1978; Kawasaki et al., 2014; Morgan and O’haire, 1978; Orchard, 1980). Specifically, Hurd and Graves (1985) and Jones et al. (1978) investigated energy consumption and benefits of root-zone heating, respectively, and indicated the commercial advantage of root-zone heating at low air temperatures. For the cucumber, root-zone cooling at high air temperature increased fruit number and weight (Moon et al., 2007b). For the sweet pepper, root-zone cooling in the warm season increased calcium content in fruits and reduced the incidence of blossom-end-rot (Benoit and Ceustermans, 2001). For the strawberry (Fragaria × ananassa Duch.), root-zone heating at low air temperature accelerated flower bud differentiation and flowering of axillary flower clusters, as well as increasing commercial fruit yield (Kim et al., 2009).
Furthermore, some reports showed that enhancement of plant growth and physiological activities by root-zone temperature control is mediated by plant growth regulators. Under optimum air temperature conditions, cytokinin (Ali et al., 1996; Kafkafi, 2001; Moon et al., 2007a), gibberellin (Ali et al., 1996; Moon et al., 2007a), and abscisic acid (Moon et al., 2007a) were affected in tomato and cucumber plants. Root-zone cooling at high air temperature caused auxin to mediate root growth in tomato plants (Kawasaki et al., 2013). However, auxin did not affect root growth with root-zone heating at low air temperature, indicating that the mechanism of root growth differs between root-zone heating and cooling (Kawasaki et al., 2014).
Root-zone temperature control techniques are generally used to keep roots at an optimum temperature and promote plant growth. On the other hand, it has been reported that excessive root-zone cooling increased fruit quality because of water stress on roots. Fujimura et al. (2012) chilled the root-zone of tomato plants from 20°C to 12°C in winter and reported that the dry weight and fruit sugar content increased, whereas the dry weight of vegetative organs decreased, especially in the leaves and roots.
Local Temperature Control of Fruits
Temperature has a large impact on fruit growth and ripening. Higher temperature promotes the fruit growth rate and shortens the number of days between flowering and harvesting in a suitable temperature range (Adams et al., 2001; Bertin, 2005). Fruit growth and ripening are affected by the temperature of the fruit itself, rather than the whole plant temperature (Kitano et al., 1998). Dorais et al. (2001) suggested a hypothesis that sink strength and dry matter distribution of fruit tend to increase with fruit temperature. Therefore, higher fruit production can be expected by heating fruits locally in the cold (winter) season.
Fanwoua et al. (2012) enclosed trusses of tomatoes into cuvettes with heating from approximately 22/18°C (day/night) to 27/23°C and reported that heating the fruit shortened the fruit growth period and reduced the final fruit size due to a reduction in the final pericarp cell volume. Gautier et al. (2005) tested fruit heating using 45°C water in flexible heating pipes circulating near growing cherry tomato fruits, and reported that the fresh weight, dry matter, and some secondary metabolites of the fruit were reduced. In these reports, the effectiveness of fruit heating of tomatoes was not shown. Fruit heating of tomatoes inhibits fruit production, contrary to expectations. On the other hand, the effectiveness of fruit heating was reported in the melon (Cucumis melo L.) and watermelon (Citurullus lanatus Matsum. et Nakai), in which sucrose accumulation was promoted in fruits (Kano, 2006; Kano et al., 2008, 2012; Matsumoto et al., 2012). The effects of fruit heating may therefore differ among crops.
In contrast to some reports on the effect of local heating of fruits, no information about local cooling on fruits was found. Because some effects of cooling fruits are thought to be prolonging the number of days between flowering and harvesting and increasing fruit size, a certain effectiveness is expected; however, dew drops on fruit that occur with cooling may contribute to the emergence of diseases and cuticle cracking.
Local Temperature Control of Shoot-tips and Flowers
Temperature control of shoot-tips had an impact on the differentiation of leaves and flower buds. Savvides et al. (2013) showed that the shoot-tip temperature deviated from air temperatures under moderate environments and measured a deviation range between −2.6°C and 3.8°C in tomatoes and between −4.1°C and 3.0°C in cucumbers, indicating that shoot-tip temperature in cucumbers was lower than that in tomatoes because of higher transpiration. Furthermore, these authors showed that the leaf differentiation rate of cucumbers derived only from the shoot-tip temperature, and not other temperatures (Savvides et al., 2016). Kawasaki et al. (2010) revealed that local heating of tomato shoot-tips from a 9.6°C to a 13.0°C surface temperature using electrical heaters in the winter season accelerated flower truss appearance (Table 2). Temperature of flower buds influences the fruit set ratio and directly impacts fruit yield. Kawasaki et al. (2010) heated flower trusses along with shoot-tips and showed that a higher flower temperature enhanced pollen viability, promoted fruit set and increased fruit yield.

Fine-tuning local temperature control of shoot-tips and flowers is difficult because shoot-tips and flowers in many vegetable plants grown in greenhouses are generally located near the plant canopy, and these positions are elevated with elongation growth. Instead of pinpoint control, therefore, vertical temperature gradients can offer practical local temperature control of shoot-tips and flowers, meaning that the upper parts containing shoot-tips and flowers are kept at an optimum temperature and the lower parts are not actively controlled. Kempkes et al. (2000) developed a temperature distribution model for greenhouse tomato production based on physical transport processes, including radiation, convection, and latent heat transfer, and simulated the effect of heating pipe positions. These authors also showed that the temperature and transpiration of the upper leaves, including shoot-tips and flowers, was increased when heating pipes were arranged close to the canopy, although air temperature increased only slightly, indicating that the use of heating pipes enables local heating, especially shoot-tip and flower heating. Kawasaki et al. (2011) tested the local heating of tomato shoot-tips and flowers using an air heater and plastic ducts hung near the plant canopy (Fig. 2), and demonstrated that the surface temperatures of shoot-tips and flowers were increased locally, whereas lower parts were maintained at low temperatures compared with a conventional duct arrangement laid on the ground (Fig. 3). In addition, the local heating reduced fuel consumption by 26% (Fig. 4), with a similar or higher yield (Table 3). Qian et al. (2015) also showed that growth and yield of tomato plants were not changed when the lower parts were maintained at temperatures lower than the optimum when the upper parts were maintained at optimum temperatures (25–15°C). This result indicates that the temperature of the lower parts of the plant has little effect on growth and yield, and is similar to that of local heating of shoot-tips and flowers (Kawasaki et al., 2011).

Local cooling of shoot-tips and flowers under high air temperature is effective, as well as local heating under low air temperature. Kawasaki and Ahn (2015) reported that local cooling at night, using a heat pump that blows cold air on tomato shoot-tips and flowers, reduced their surface temperature approximately from 27°C to 25°C under a 30°C daily mean air temperature, leading to an increased fruit yield and marketable fruit ratio. Although local cooling has proven to be effective in increasing fruit production, its energy costs have not been reported. Therefore, further research on the economic benefits of local cooling is required.
For the strawberry, local temperature control has also been studied. Strawberries differ from the other fruit and vegetables, as they have shoot-tips (crowns) located near the ground surface, and their position does not move with growth. Crown temperature can be controlled easily by a system similar to that used for root-zone temperature control. Sato and Kitajima (2010) heated crowns using electrically heated wire to 21°C under an 8°C minimum daily temperature and observed that the differentiation of leaves and flower buds was accelerated, and that petiole elongation and flowering were promoted. Hidaka et al. (2017) cooled strawberry crowns to 20°C using a plastic tube filled with chilled water under high air temperature conditions (controlled day/night temperatures of 30/27°C) and showed acceleration of differentiation and flowering on first inflorescence, as well as an increased early marketable yield. Dan et al. (2015) demonstrated crown cooling in summer and heating in winter in a strawberry forcing culture and showed that marketable fruit yield increased by 9–15% when the crown temperature was kept around 20°C throughout the culture compared to the control condition.
Although the contribution of energy saving with local temperature control of shoot-tips and flowers has not yet been fully clarified, it will likely reduce energy consumption while resulting in the same yield compared to conventional methods where shoot-tip and flower temperatures are kept the same and the other parts are not actively temperature-controlled. Moreover, this technique can increase fruit yield with less energy consumption when the temperature is maintained nearer the optimal level than that of conventional techniques.
Local Temperature Control of Other Parts
Almost all studies on local temperature control of plant organs are described above; however, few reports are available on leaf and stem temperature control, as they are relatively insensitive to temperature changes compared to other organs (Table 1). On the other hand, basal stem heating ensured similar or greater effects on flower differentiation and fruit yield in the eggplant (Solanum merongena kouki) when its temperature was kept at 25°C and the set point of the heater was lowered to 10°C from 12°C (Moriyama and Oku, 2012; Moriyama et al., 2011). These authors also showed that fuel consumption was reduced by approximately 30% using basal stem heating with a 2°C lower heater setting. Furthermore, basal stem heating promoted the growth of lateral branches and differed from root-zone heating, which promoted primary stem and root growth (Moriyama et al., 2012). This difference could be due to a plant growth regulator; however, further studies are required to confirm this hypothesis.
Conclusion and Perspectives
Temperature management in greenhouses has primarily used uniform temperature conditions throughout the greenhouse. However, recent studies on the temperature reactions of each plant organ revealed that crops showed high productivity even when temperature conditions were not uniform. On the basis of these findings, local temperature control techniques have been developed to create accurate ununiform temperature gradients. They can contribute to reducing energy expenditure and the associated environmental impact; therefore, greenhouse vegetable production using these techniques would be more beneficial than that using conventional temperature control techniques. On the other hand, there is little information on the exact cost reduction and the resulting lower environmental footprint. Furthermore, physiological mechanisms in plants benefiting from local temperature control are not well known, for example, gene expression and balance of plant growth regulators. More research on these areas is required in the future.
The combination of some local temperature control techniques could lead to higher productivity than that of a single technique; however, a combination technique would be more difficult to control and would increase the initial cost of the system. Therefore, more studies are necessary to develop the best technique for local temperature control that can be applied in greenhouse production.
Literature Cited
- Adams, S. R. and V. M. Valdés. 2002. The effect of periods of high temperature and manipulating fruit load on the pattern of tomato yields. J. Hort. Sci. Biotech. 77: 461–466.
- Adams, S. R., K. E. Cockshull and C. R. J. Cave. 2001. Effect of temperature on the growth and development of tomato fruits. Ann. Bot. 88: 869–877.
- Al-Mulla, Y. A., M. Al-Balushi, H. Al-Busaidi, A. Al-Madhouri, C. Kittas and N. Katsoulas. 2018. Analysis of microclimate and cucumber fruit yield in a screenhouse and an evaporatively cooled greenhouse in a semi-arid location. Transactions of the ASABE. 61: 619–629.
- Ali, I. A., U. Kafkafi, I. Yamaguchi, Y. Sugimoto and S. Inanaga. 1996. Effects of low root temperature on sap flow rate, soluble carbohydrates, nitrate contents and on cytokinin and gibberellin levels in root xylem exudate of sand-grown tomato. J. Plant Nutri. 19: 619–634.
- Benoit, F. and N. Ceustermans. 2001. Impact of root cooling on blossom end rot in soilless paprika. Acta Hortic. 548: 319–323.
- Bertin, N. 2005. Analysis of the tomato fruit growth response to temperature and plant fruit load in relation to cell division, cell expansion and DNA endo endoreduplication. Ann. Bot. 95: 439–447.
- Charles, W. B. and R. E. Harris. 1972. Tomato fruit-set at high and low temperatures. Can. J. Plant Sci. 52: 497–506.
- Chong, P. C. and T. Ito. 1982. Growth, fruit yield and nutrient absorption of tomato plant as influenced by solution temperature in nutrient film technique. J. Japan. Soc. Hort. Sci. 51: 44–50.
- Dan, K., W. Sugeno, S. Nakahara, N. Goto, Y. Iwasaki, I. Takano, M. Okimura, K. Hidaka, T. Takayama and H. Imamura. 2015. Experiment on the crown-temperature control technique in forcing culture of strawberries in Miyagi. Bull. NARO Kyushu Okinawa Agric. Res. Cent. 64: 1–11 (In Japanese with English abstract).
- Davis, R. M. and J. C. Lingle. 1961. Basis of shoot response to root temperature in tomato. Plant Physiol. 36: 153–162.
- Dodd, I. C., J. He, C. G. N. Turnbull, S. K. Lee and C. Critchley. 2000. The influence of supra-optimal root-zone temperatures on growth and stomatal conductance in Capsicum annuum L. J. Exp. Bot. 51: 239–248.
- Dorais, M., A. P. Papadopoulos and A. Gosselin. 2001. Greenhouse tomato fruit quality. Hort. Rev. 26: 239–319.
- Fang, H., Q. Yang, Y. Zhang, Q. Sum, W. Lu, Y. Tong and H. Liang. 2015. Performance of a solar heat collection and release system for improving night temperature in a Chinese solar greenhouse. Appl. Eng. Agric. 31: 283–289.
- Fanwoua, J., P. de Visser, E. Heuvelink, G. Angenent, X. Yin, L. Marcelis and P. Struik. 2012. Response of cell division and cell expansion to local fruit heating in tomato fruit. J. Amer. Soc. Hort. Sci. 137: 294–301.
- Fernandez, J. E. and B. J. Bailey. 1994. The influence of fans on environmental conditions in greenhouse. J. Agric. Eng. Res. 58: 201–210.
- Fujimura, S., K. Suzuki, M. Nagao and M. Okada. 2012. Acclimation to root chilling increases sugar concentrations in tomato (Solanum lycopersicum L.) fruits. Sci. Hortic. 147: 34–41.
- Fujishige, N., T. Sugiyama and R. Ogata. 1991. Effect of root temperature on the flower formation and fruit yield of tomatoes. J. Japan. Soc. Hort. Sci. 60: 97–103 (In Japanese with English abstract).
- Gautier, H., A. Rocci, D. Grasselly, M. Buret and M. Causse. 2005. Effect of adding heating pipes on the temperature and the physical and chemical traits of tomato fruits. Acta Hortic. 691: 59–66.
- Gent, M. P. N. and Y-Z. Ma. 1998. Diurnal temperature variation of the root and shoot affects yield of greenhouse tomato. HortScience 33: 47–51.
- Gent, M. P. N. and Y-Z. Ma. 2000. Mineral nutrition of tomato under diurnal temperature variation of root and shoot. Crop Sci. 40: 1629–1636.
- Gent, M. P. N., J. H. Thone and D. E. Aylor. 1979. Split-night temperatures in a greenhouse: the effects on the physiology and growth on plants. Conn. Agr. Exp. Sta. Bull. 781: 1–15.
- Giacomelli, G. A. and W. J. Roberts. 1993. Greenhouse covering systems. HortTechnology 3: 50–58.
- Gosselin, A. and M. J. Trudel. 1982. Influence de la temperature du substrat sur la croissance, le developpement et le contenu en elements mineraux de plants de tomate (cv. Vendor). Can. J. Plant Sci. 62: 751–757 (In French with English abstract).
- Gosselin, A. and M. J. Trudel. 1983a. Interactions between air and root temperatures on greenhouse tomato: I. growth, development, and yield. J. Amer. Soc. Hort. Sci. 108: 901–905.
- Gosselin, A. and M. J. Trudel. 1983b. Interactions between air and root temperatures on greenhouse tomato: II. mineral composition of plants. J. Amer. Soc. Hort. Sci. 108: 905–909.
- Gosselin, A. and M. J. Trudel. 1984. Interactions between root-zone temperature and light levels on growth, development and photosynthesis of Lycopersicon esculentum Mill. cultivar ‘Vendor’. Sci. Hortic. 23: 313–321.
- Gosselin, A. and M. J. Trudel. 1985. Influence of root-zone temperature on growth, development and yield of cucumber plants cv. Toska. Plant and Soil 85: 327–336.
- Gosselin, A. and M. J. Trudel. 1986. Root-zone temperature effects on pepper. J. Amer. Soc. Hort. Sci. 111: 220–224.
- Grimstad, S. O. and E. Frimanslund. 1993. Effect of different day and night temperature regimes on greenhouse cucumber young plant production, flower bud formation and early yield. Sci. Hortic. 53: 191–204.
- Handarto, M. Hayashi, E. Goto and T. Kozai. 2007. Experimental verification of control logic for operation of a fog cooling system for a naturally ventilated greenhouse. Environ. Control Biol. 45: 47–58.
- Handarto, M. Hayashi, K. Ohyama, H. Toida, E. Goto and T. Kozai. 2006. Developing control logic for a high-pressure fog cooling system operation for a naturally ventilated greenhouse. Environ. Control Biol. 44: 1–9.
- He, J., S. K. Lee and I. C. Dodd. 2001. Limitations to photosynthesis of lettuce grown under tropical conditions: alleviation by root-zone cooling. J. Exp. Bot. 52: 1323–1330.
- Heuvelink, E. and M. Dorais. 2005. Crop growth and yield. p. 85–144. In: E. Heuvelink (ed.). TOMATOES. CABI publishing. Wallingford.
- Hidaka, K., K. Dan, H. Imamura and T. Takayama. 2017. Crown-cooling treatment induces earlier flower bud differentiation of strawberry under high air temperatures. Environ. Control Biol. 55: 21–27.
- Hurd, R. G. and C. J. Graves. 1985. Some effects of air and root temperatures on the yield and quality of glasshouse tomatoes. J. Hort. Sci. 60: 359–371.
- Hurewitz, J. and H. W. Janes. 1983. Effect of altering the root-zone temperature on growth, translocation, carbon exchange rate, and leaf starch accumulation in the tomato. Plant Physiol. 73: 46–50.
- Ikeda, H. and T. Osawa. 1988. Effects of NO3/NH4 ratios and temperature of the nutrient solution on growth, yields, and blossom-end rot incidence of tomato. J Japan. Soc. Hort. Sci. 57: 62–69.
- Ilahi, W. F. F., D. Ahmad and M. C. Husain. 2017. Effects of root zone cooling on butterhead lettuce grown in tropical conditions in a coir-perlite mixture. Hortic. Environ. Biotechnol. 58: 1–4.
- Jeff, A. E. and D. A. G. Jones. 1973. The effect of heating pipe position on the cropping of early tomatoes. Exp. Hort. 25: 127–132.
- Jones, D. A. G., I. Sandwell and C. J. W. Talent. 1978. The effect of soil temperature when associated with low air temperatures on the cropping of early tomatoes. Acta Hortic. 76: 167–171.
- Joudi, K. A. and A. A. Farhan. 2014. Greenhouse heating by solar air heaters on the roof. Renew. Energy 72: 406–414.
- Kabu, K. L. and E. W. Toop. 1970. Influence of soil temperature and potassium fertilization on magnesium content of tomato plants. Can. J. Plant Sci. 50: 740–742.
- Kafkafi, U. 2001. Root zone parameters controlling plant growth in soilless culture. Acta Hortic. 554: 27–38.
- Kano, Y. 2006. Effect of heating fruit on cell size and sugar accumulation in melon fruit (Cucumis melo L.). HortScience 41: 1431–1434.
- Kano, Y., Y. Ikeshita, Y. Kanamori and N. Fukuoka. 2008. Effects of night-heating of fruit on cell size and sucrose accumulation in the outer portion of watermelon (Citrullus lanatus Matsum. et Nakai). HortScience 43: 792–794.
- Kano, Y., J. Matsumoto, Y. Aoki and T. Madachi. 2012. Effects of heating bearing shoot near fruit on cell size, sucrose metabolizing enzymes and sugar accumulation in watermelon fruit. J. Japan. Soc. Hort. Sci. 81: 171–176.
- Kawasaki, Y. and D. H. Ahn. 2015. Local cooling at night around flowering trusses improved yield of tomato grown under hot conditions. J. SHITA 27: 137–143 (In Japanese with English abstract).
- Kawasaki, Y., S. Matsuo, Y. Kanayama and K. Kanahama. 2014. Effect of root-zone heating on root growth and activity, nutrient uptake, and fruit yield of tomato at low air temperatures. J. Japan. Soc. Hort. Sci. 83: 295–301.
- Kawasaki, Y., S. Matsuo, K. Suzuki, Y. Kanayama and K. Kanahama. 2013. Root-zone cooling at high air temperatures enhances physiological activities and internal structures of roots in young tomato plants. J. Japan. Soc. Hort. Sci. 82: 322–327.
- Kawasaki, Y., K. Suzuki, K. Yasuba, H. Kawashima, H. Sasaki and M. Takaichi. 2010. Effect of local heating around the tomato shoot apex and flower clusters on plant surface temperature and characteristics related to fruit yield. Hort. Res. (Japan) 9: 345–350 (In Japanese with English abstract).
- Kawasaki, Y., K. Suzuki, K. Yasuba and M. Takaichi. 2011. Effect of local air heating by a hanging duct near the tomato shoot apex and flower clusters on vertical temperature distribution, fruit yield and fuel consumption. Hort. Res. (Japan) 10: 395–400 (In Japanese with English abstract).
- Kawashima, H. 2015. Development of a new energy-saving pipe-framed greenhouse. JARQ 49: 235–243.
- Kempkes, F. L. K., N. J. Van de Braak and J. C. Bakker. 2000. Effect of heating system position on vertical distribution of crop temperature and transpiration in greenhouse tomatoes. J. Agric. Eng. Res. 75: 57–64.
- Kim, Y. S., M. Endo, Y. Kiriiwa, L. Chen and A. Nukaya. 2009. Effects of root zone heating during daytime on the flowering, growth and yield of strawberry ‘Akihime’ grown in substrate culture. Hort. Res. (Japan) 8: 193–199 (In Japanese with English abstract).
- Kitano, M., T. Araki and H. Eguchi. 1998. Environmental effects on dynamics of fruit growth and photoassimilate translocation in tomato plants. I. Effects of irradiation and day/night air temperature. Environ. Control in Biol. 36: 159–167 (In Japanese with English abstract).
- Klock, K. A., H. G. Taber and W. R. Graves. 1997. Root respiration and phosphorus nutrition of tomato plants grown at a 36°C root-zone temperature. J. Amer. Soc. Hort. Sci. 122: 175–178.
- Kramer, P. J. 1934. Effects of soil temperature on the absorption of water by plants. Science 79: 371–372.
- Kuroyanagi, T. 2016. Current usage of air circulators in greenhouses in Japan. JARQ 50: 7–12.
- Lingle, J. C. and R. M. Davis. 1959. The influence of soil temperature and phosphorus fertilization on the growth and mineral absorption of tomato seedlings. J. Amer. Soc. Hort. Sci. 73: 312–322.
- Lopez, A., D. L. Valera, F. D. Molina-Aiz and A. Peña. 2010. Experimental evaluation by sonic anemometry of airflow in a mediterranean greenhouse equipped with a pad-fan cooling system. Transactions of the ASABE. 53: 945–957.
- Matsumoto, J., Y. Kano, T. Madachi and Y. Aoki. 2012. Heating bearing shoots near fruits promotes sugar accumulation in melon fruit. Sci. Hortic. 133: 18–22.
- Moon, J. H., H. O. Boo and I. O. Jang. 2007a. Effect of root-zone temperature on water relations and hormone contents in cucumber. Hort. Environ. Biotechnol. 48: 257–264.
- Moon, J. H., Y. K. Kang and H. D. Suh. 2007b. Effect of root-zone cooling on the growth and yield of cucumber at supraoptimal air temperature. Acta Hortic. 761: 271–274.
- Morgan, J. V. and R. O’Haire. 1978. heated hydroponic solutions as an energy saving technique. Acta Hortic. 76: 173–180.
- Moriyama, T. and K. Oku. 2012. Development of basal stem heating system with plastic tunnel and branch duct in forcing culture of eggplant. Hort. Res. (Japan) 11: 531–536 (In Japanese with English abstract).
- Moriyama, T., H. Fushihara and K. Oku. 2011. Effects of basal stem heating on fruit set, yield and fruit quality in forcing culture of eggplant. Hort. Res. (Japan) 10: 545–550 (In Japanese with English abstract).
- Moriyama, T., H. Fushihara and K. Oku. 2012. Effects of heating parts and time on eggplant fruit shape and physiology in localized heating culture. Hort. Res. (Japan) 11: 337–341 (In Japanese with English abstract).
- NaanDanJain irrigation Ltd. 2012. Tomato cultivation in open fields and greenhouses. <http://www.naandanjain.com/uploads/catalogerfiles/tomato-2/Tomato_eng_booklet_190812final%20.pdf>.
- Nakano, Y. 2007. Response of tomato roots systems to environmental stress under soilless culture. JARQ 41: 7–15.
- Nkansah, G. O. and T. Ito. 1994. Comparative studies on growth and development of heat-tolerant and non heat-tolerant tomato plants grown at different root-zone temperatures. J. Japan. Soc. Hort. Sci. 62: 775–780.
- Nkansah, G. O. and T. Ito. 1995a. Effect of air and root-zone temperatures on physiological characteristics and yield of heat-tolerant and non heat-tolerant tomato cultivars. J. Japan. Soc. Hort. Sci. 64: 315–320.
- Nkansah, G. O. and T. Ito. 1995b. Comparison of mineral absorption and nutrient composition of heat-tolerant and non heat-tolerant tomato plants at different root-zone temperatures. J. Hort. Sci. 70: 453–460.
- Ogura, Y. and T. Mukai. 1988. Thermal insulation characteristics of a curved roof greenhouse with an internal curtain for spraying water. Nogyo Shisetsu 18 (3): 58–62 (In Japanese with English abstract).
- Orchard, B. 1980. Solution heating for the tomato crop. Acta Hortic. 98: 19–28.
- Papadopoulos, A. P. and H. Tiessen. 1987. Root and air temperature effects on the elemental composition of tomato. J. Amer. Soc. Hort. Sci. 112: 988–993.
- Peet, M. M., D. H. Willits and R. Gardner. 1997. Response of ovule development and post-pollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. J. Exp. Bot. 48: 101–111.
- Qian, T., J. A. Dieleman, A. Elings, A. de Gelder and L. F. M. Marcelis. 2015. Response of tomato crop growth and development to a vertical temperature gradient in a semi-closed greenhouse. J. Hort. Sci. Biotech. 90: 578–584.
- Sasaki, K. and T. Itagi. 1989. Effect of root cooling treatment of nutrient solution on growth and yield of tomato, cucumber and melon. Environ. Control in Biol. 27: 89–95.
- Sato, K. and N. Kitajima. 2010. Local heating temperature effects on the growth and yield of strawberries in high-bench culture. Bull. Fukuoka Agric. Res. Cent. 29: 27–32 (In Japanese with English abstract).
- Sato, S., M. Kamiyama, T. Iwata, N. Makita, H. Furukawa and H. Ikeda. 2006. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processed in male reproductive development. Ann. Bot. 97: 731–738.
- Sato, S., M. M. Peet and J. F. Thomas. 2000. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 23: 719–726.
- Sato, S., M. M. Peet and J. F. Thomas. 2002. Determining critical pre- and post-anthesis periods and physiological processed in Lycopersicon esculentum Mill. exposed to moderately elevated temperatures. J. Exp. Bot. 53: 1187–1195.
- Savvides, A., J. A. Dieleman, W. van Ieperen and L. F. M. Marcelis. 2016. A unique approach to demonstrating that apical bud temperature specifically determines leaf initiation rate in the dicot Cucumis sativus. Planta 243: 1071–1079.
- Savvides, A., W. van Ieperen, J. A. Dieleman and L. F. M. Marcelis. 2013. Meristem temperature substantially deviates from air temperature even in moderate environment: is the magnitude of this deviation species-specific? Plant Cell Environ. 36: 1950–1960.
- Shishido, Y. and H. Kumakura. 1994. Effects of root temperature on photosynthesis, transpiration, translocation and distribution of 14C-photoassimilates and root respiration in tomato. J. Japan. Soc. Hort. Sci. 63: 81–89 (In Japanese with English abstract).
- Teitel, M. and J. Tanny. 1998. Radiative heat transfer from heating tubes in a greenhouse. J. Agric. Eng. Res. 69: 185–188.
- Tindall, J. A., H. A. Mills and D. E. Radcliffe. 1990. The effect of root zone temperature on nutrient uptake of tomato. J. Plant Nutri. 13: 939–956.
- Toki, T. 1970. Physiological studies on optimal environment in growing of the vegetable crops under the glass- and plastic-houses. 1.) Temperature analysis for growing cucumber. Bull. Chiba Agric. Exp. Stn. 11: 62–72 (In Japanese with English summary).
- Tomos, M. L. 1963. Temperature inhibition of carotene synthesis in tomato. Bot. Gaz. 124: 180–185.
- Tong, Y., T. Kozai, N. Nishioka and K. Ohyama. 2012. Reductions in energy consumption and CO2 emissions for greenhouses heated with heat pumps. Appl. Eng. Agric. 28: 401–406.
- van der Ploeg, A., M. van der Meer and E. Heuvelink. 2007. Breeding for a more energy efficient greenhouse tomato: past and future perspectives. Euphytica 158: 129–138.
- Wang, X., W. Zhang, Y. Miao and L. Gao. 2016. Root-zone warming differently benefits mature and newly unfolded leaves of Cucumis sativus L. seedlings under sub-optimal temperature stress. PLoS ONE 11: e0155298. DOI: 10.1371/journal.pone.0155298.
- Went, F. W. 1943. Plant growth under controlled conditions. I. The air-conditioned greenhouses at the California Institute of Technology. Amer. J. Bot. 30: 157–163.
- Went, F. W. 1944. Plant growth under controlled conditions. II. Thermoperiodicity in growth and fruiting of the tomato. Amer. J. Bot. 31: 135–150.
- Yelle, S., A. Gosselin and M. J. Trudel. 1987. Effect of atmospheric CO2 concentration and root-zone temperature on growth, mineral nutrition, and nitrate reductase activity of greenhouse tomato. J. Amer. Soc. Hort. Sci. 112: 1036–1040.
- Zhou, S., Y. Zhang, Q. Yang, R. Cheng, H. Fang, X. Ke, W. Lu and B. Zhou. 2016. Performance of active heat storage-release unit assisted with a heat pump in a new type of Chinese solar greenhouse. Appl. Eng. Agric. 32: 641–650.



