ORIGINAL ARTICLES
Breeding for Long Vase Life in Dahlia (Dahlia variabilis) Cut Flowers
2019 Volume 88 Issue 4 Pages 521-534

Details
2019 Volume 88 Issue 4 Pages 521-534
Flower vase life is one of the most important traits for ornamental plants. The vase life of cut dahlia (Dahlia variabilis) flowers is very short, and genetic improvement of this trait is desirable. We started a breeding research program in 2014 to improve the vase life of dahlia flowers using conventional cross-breeding techniques. We found large significant differences in flower vase life among 24 dahlia cultivars: Nine cultivars had long vase life (e.g., ‘Syukuhai’, ‘Rinka’, and ‘Micchan’); eight had normal vase life (e.g., ‘Kamakura’, ‘Agitate’, and ‘Benifusya’); and seven had short vase life (e.g., ‘Gin-Ei’, ‘Port Light Pair Beauty’, and ‘Yumesuiren’). We used 22 cultivars as initial breeding materials, repeatedly crossed them, and selected promising offspring with long vase life for three generations from 2014 to 2018. Two cycles of selection and crossing led to a 1.7-day increase in vase life (population mean) from the first to the third generation, clearly showing that this approach can extend the vase life of dahlia flowers. The mean vase life of ‘Kamakura’, a leading white dahlia cultivar in Japan, was 5.0–6.2 days in distilled water, 6.0–6.8 days in an isothiazolinic antibacterial agent CMIT/MIT solution (5-chloro-2-methyl-4-isothiazolin-3-one and 2-methyl-4-isothiazolin-3-one) and 6.0–7.6 days in a GLA solution (10 g·L−1 glucose, 0.5 ml·L−1 CMIT/MIT, and 50 mg·L−1 aluminum sulfate), whereas in six finally selected lines it was 6.2–12.0 days in distilled water, 6.6–10.2 days in CMIT/MIT solution, and 9.4–13.6 days in GLA solution (1.4–2.1 times that in ‘Kamakura’). In particular, the selected second-generation line 606-46 showed a stably longer vase life than ‘Kamakura’. ‘Micchan’, which has a long vase life, was a common progenitor used for breeding of parental lines in cross combinations with long vase life in the second generation and all cross combinations in the third generation. The final six selected lines with long vase life were all progeny of ‘Micchan’. Our results strongly suggest that ‘Micchan’ has genes related to long flower vase life, and that the trait is heritable.
The vase life of cut ornamental flowers determines their quality and ability to satisfy consumer preferences, thereby stimulating repeat purchasing. It is therefore one of the most important breeding targets (Onozaki, 2018a). Flower distributors want to reduce the deterioration in flower quality during the long transportation time from producer to consumer (Shibuya, 2018). Therefore, the ability to control petal aging and flower senescence is of great interest to breeders (Zuker et al., 1998).
The main breeding targets for ornamentals used to be visual qualities such as appearance, flower color, type, size, and stem length (Boxriker et al., 2017). Vase life is a highly complex quantitative trait that involves multiple genes with additive effects (Boxriker et al., 2017, 2018), and few attempts to improve flower longevity were made before the 1990s (Harding et al., 1981; Van Eijk and Eikelboom, 1976). Breeding for extended vase life is challenging because vase life is affected by many different deterioration processes (Van Geest et al., 2017). The assessment of vase life is labor-intensive (Boxriker et al., 2018) because each genotype must be grown and harvested, and cut flowers must be evaluated every day for senescence symptoms. However, the longevity of cut flowers has now become an important quality factor because short-lived flowers have limited consumer appeal and therefore limited marketability. Breeding to improve vase life has been carried out in snapdragon (Weber et al., 2005), chrysanthemum (Van Geest et al., 2017), gerbera (Wernett et al., 1996a, b), carnation (Onozaki, 2018a; Onozaki et al., 2001, 2006a, b, 2011, 2015, 2018), Asiatic hybrid lily (van der Meulen-Muisers et al., 1999), and rose (Carvalho et al., 2015; Fanourakis et al., 2012).
The Institute of Vegetable and Floriculture Science (NIVFS, NARO, Japan) has bred carnations (Dianthus caryophyllus L.) for long vase life since 1992 (Onozaki, 2018a). We crossed and selected promising offspring with long vase life for seven generations from 1992 to 2008. The mean vase life was improved from 7.4 days in the first generation to 15.9 days in the 7th generation through conventional crossing techniques. Three developed cultivars, ‘Miracle Rouge’, ‘Miracle Symphony’ (Onozaki et al., 2006a), and ‘Kane Ainou 1-go’ (Hotta et al., 2016), with a genetically determined long vase life, and two lines, 532-6 and 806-46b, with an ultra-long vase life (>27 days) under standard conditions, commonly produce only trace amounts of ethylene during natural senescence and show neither petal inrolling nor rapid wilting. Our nearly two-decade-long breeding and research program revealed a close correlation between vase life and ethylene production in cut carnations (Onozaki et al., 2018).
Dahlia (Dahlia variabilis; Asteraceae), an important bulb crop sold as cut flowers, garden ornamentals and potted plants in many countries, has become a popular cut flower in Japan in recent years (Onozaki, 2018b). Dahlias are not surveyed in the nationwide Statistics of Agriculture, Forestry and Fisheries in Japan, but the number of cut dahlia flowers handled in the Tokyo metropolitan market increased from 1,024,124 in 2002 to 4,733,901 in 2018 (a 362% increase; http://www.shijou-tokei.metro.tokyo.jp/). Demand for cut dahlia flowers has been increasing year by year. Dahlias show huge variation in flower traits such as color, shape, and size, owing to their high polyploidy (Okumura and Fujino, 1989; Wegner and Debener, 2008), likely autoallooctoploidy (2n = 8x = 64) (Gatt et al., 1998). However, the vase life of cut dahlias without chemical treatment is usually only about four days at room temperature, with some differences among cultivars (Ichimura et al., 2011). This very short vase life has curtailed the expansion of demand for cut dahlia flowers (Shimizu-Yumoto and Ichimura, 2013). Although continuous exposure to ethylene (2 or 10 μL·L−1) significantly accelerated petal abscission in cut flowers of ‘Kokucho’, a leading dark-red dahlia cultivar in Japan, silver thiosulfate complex, an inhibitor of ethylene action, did not extend its vase life (Shimizu-Yumoto and Ichimura, 2013). Dole et al. (2009) reported that the vase life of the dahlia ‘Karma Thalia’ was unaffected by exogenous ethylene or anti-ethylene treatments (silver thiosulfate complex or 1-methylcyclopropene). These reports suggest that ethylene is not a very important factor in inducing flower senescence in dahlia.
Three Japanese dahlia breeders informed us that long vase life of dahlia flowers may be related to a lack of stem cavities, i.e., a smaller stem diameter (personal communications: Koji Washizawa and Yusaku Konishi, September 2014; Yoshiki Amano, July 2015). Therefore, we investigated the relationship between stem diameter and flower vase life in the first and second generations.
Tsujimoto et al. (2016b) showed genetic variation in the vase life of cut flowers of 27 dahlia cultivars and reported that several commercial dahlia cultivars (e.g. ‘Rinka’, ‘Syukuhai’, ‘Moon Waltz’, ‘Benifusya’, ‘Micchan’, and ‘Akebono-Temari’ in winter; ‘Rinka’, ‘Syukuhai’, ‘Micchan’, ‘Akebono-Temari’, ‘Pink Sapphire’, and ‘Moon Waltz’ in summer) have extended vase life. This genetic variation should make it possible to breed lines with a genetically determined long vase life. In 2014, we started a breeding research program to improve the vase life of dahlia flowers using conventional cross-breeding techniques that had proved to be very effective in carnation breeding (Onozaki, 2018a). The aim of this program is to produce commercially successful dahlia cultivars with a long vase life. Here, we report the results of crossing and selection over three generations to improve the vase life of dahlia.
The vase life of 24 commercial dahlia cultivars used as cut flowers (Table 1) was evaluated. Bulbs were purchased in April 2014 from Akita International Dahlia Park (Akita, Japan); Takii & Co., Ltd. (Kyoto, Japan); Kokkaen & Co., Ltd. (Osaka, Japan); or Fukukaen Nursery & Bulb Co., Ltd. (Nagoya, Japan). Plants were grown in an open field of NIVFS, Tsukuba, Japan, from May to November. Commercial fertilizer (Cyclo-Di-Urea (CDU) S222; Zen-Noh, Tokyo, Japan) was applied at N:P2O5:K2O = 20:20:20 kg/10a just before planting each year. Two bulbs per cultivar were planted 40 cm apart on 13 May 2014 and 12 May 2015. Liquid compound fertilizer (Yoeki-Dokou No. 1, N:P2O5:K2O = 15:8:17; OAT Agrio Co., Ltd., Tokyo, Japan, 1:1000 dilution) was applied once a week from June to October in both years. The shoots of each plant were pinched to two or three nodes on 17 June 2014 and 16 June 2015, and the plants were grown following standard production methods in the open field for seasonal flowering (Yamagata, 2018).

Vase life (days) of cut flowers of dahlia cultivars in distilled water (DW), antibacterial agent (CMIT/MIT), or GLA solution.
Vase life of dahlia cultivars was evaluated from late July to October in 2014 and from September to early November in 2015. As an antibacterial agent, we used a CMIT/MIT (Kathon CG, formerly Legend MK; Rohm and Haas Japan K.K., Tokyo, Japan) solution containing 11.3 g·L−1 5-chloro-2-methyl-4-isothiazolin-3-one (CMIT) and 3.9 g·L−1 2-methyl-4-isothiazolin-3-one (MIT) as active ingredients. Flowers were harvested when the first row of petals from the outside had opened at right angles to the stem. The flowers were then cut to a stem length of 40 cm and all leaves except the top ones were removed. Two or three cut flowers were placed in 300- or 500-mL conical beakers containing distilled water (DW) or 0.5 mL·L−1 CMIT/MIT solution or GLA solution (10 g·L−1 glucose, 0.5 mL·L−1 CMIT/MIT [Legend MK], 50 mg·L−1 aluminum sulfate; Ichimura et al., 2006). As a result, we used three different solutions (DW, CMIT/MIT, and GLA) to evaluate 24 cultivars.
Flowers were maintained at 23°C, 70% relative humidity, under a 12-h photoperiod provided by cool fluorescent lamps (10 μmol·m−2·s−1 irradiance), and were evaluated daily. The solutions were not exchanged, but if the level decreased to less than two-thirds of the initial level, more was added.
Flower senescence was classified visually as wilting, wilting with browning, browning, or petal abscission (Fig. 1). The vase life of each flower was determined as the number of days from harvest until one-third of all of petals showed one of the four patterns.

Morphology of flowers with four senescence patterns: (A) wilting, (B) wilting with browning, (C) browning, (D) petal abscission.
Finally, the grand mean (total average of DW, CMIT/MIT, and GLA) was calculated and assigned an index as follows: 1 (long vase life), ≥ 7 days; 2 (normal vase life), ≥ 6 to < 7 days; 3 (short vase life) < 6 days.
Crossing and selectionAs initial breeding materials, we chose 22 of the 24 cultivars (‘Gin-Ei’ and ‘Black Cat’ were excluded; see Table 1) with large differences in vase life. In autumn 2014, crosses among these cultivars were made in a vinyl house and open field (Table 2) using the standard crossing method for dahlias (Konishi, 2009; Onozaki, 2018b). All ray florets were removed from the whole flower and bagged with water-resistant paper bags. After the disk florets had opened, the bags were removed and the flowers were pollinated by hand. After pollination, they were bagged again to prevent pollination by insects. About 45 days later (just before the first frost in mid-November), the crossed flowers were cut from the plants and seeds were collected.

Cross combinations and results of crosses for the first generation.
We then crossed and selected promising offspring with long vase life for three generations from 2014 to 2018. On 23 April 2015 and 14 May 2015, the 1,109 seeds obtained were sown and 439 plants germinated by 29 June 2015; 419 of these were grown in the open field. Plants that did not flower until 18 September 2015 (the last day of evaluation) were discarded, and the remaining 314 plants were considered as the first generation (Table 2). In October 2015, 64 plants with the longest mean vase life (> 5.1 days) were selected and used as the next crosses. From these crosses, 571 seeds were obtained. These were sown on 4 April 2016, and 355 plants germinated by 12 June 2016; 349 of these were grown in the open field. Plants that did not flower until 16 September 2016 (the last day of evaluation) were discarded, and the remaining 308 plants were considered as the second generation (Table 3). In October 2016, 73 plants with the longest mean vase life (> 6.0 days) were selected and used for the next crosses. From these crosses, 764 seeds were obtained and were sown on 3 April 2017 or 29 March 2018 and 253 plants in total germinated by 25 May 2017 or 4 June 2018; 247 of these were grown in the open field. Plants that did not flower until 15 September 2017 or 10 September 2018 (the last days of evaluation) were discarded, and the remaining 155 plants were considered as the third generation (Table 4). In October 2017 and 2018, a total of 58 plants with the longest mean vase life (> 6.1 or > 6.2 days, respectively) were selected.

Cross combinations and results of crosses for the second generation.
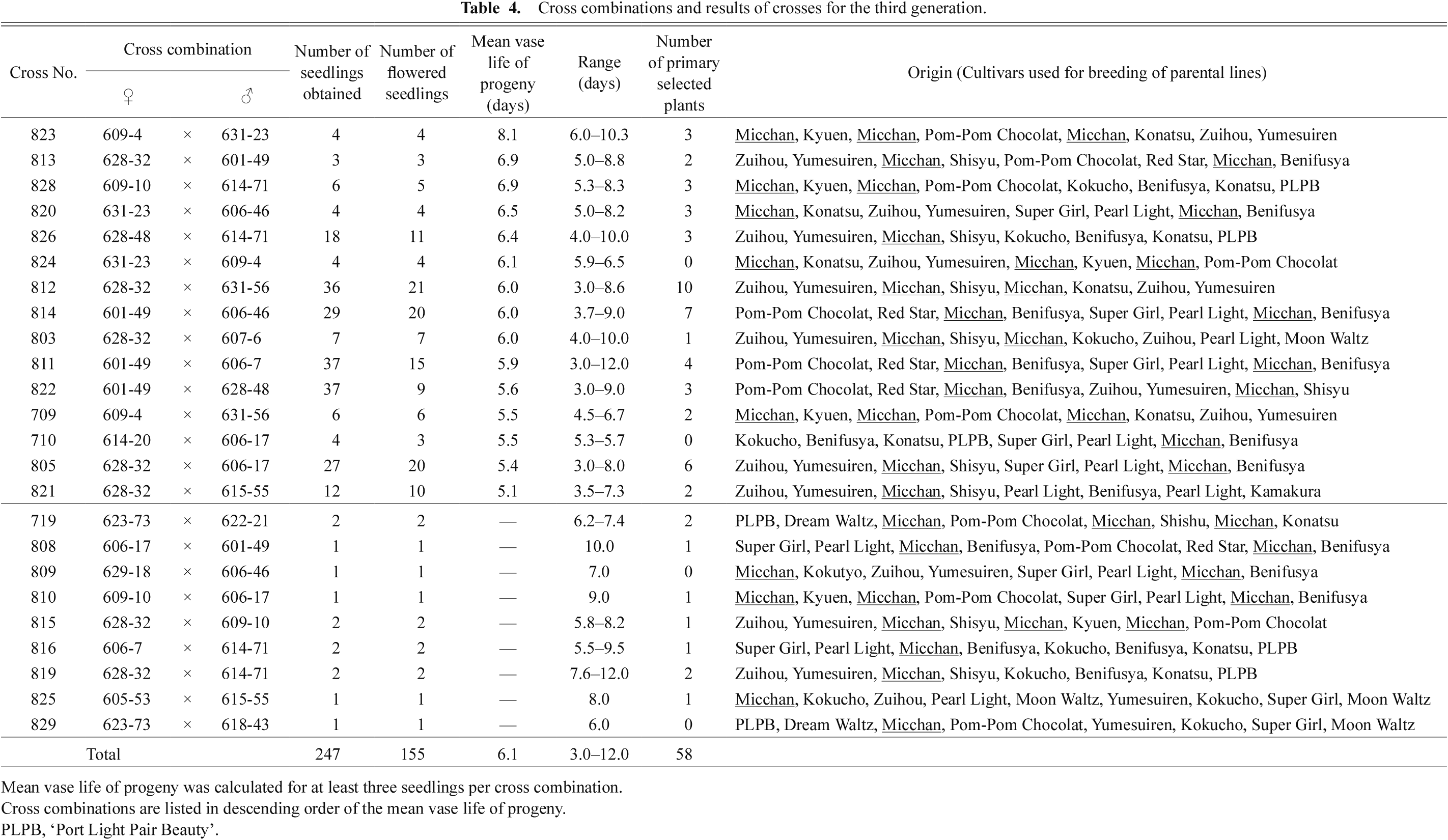
Cross combinations and results of crosses for the third generation.
The initially selected first- and second-generation lines were propagated vegetatively, and 11 lines out of 64 in 2016 and 10 lines out of 73 in 2017 were further selected. We examined the vase life of the cut flowers of the initially selected lines in detail in two different seasons and using three different cultivation styles, that is, a winter–spring season in a greenhouse heated above 12°C with daylength extension to 14.5 h, and/or a summer–autumn season in an open field or a vinyl house (no heating, natural daylength). Lines with a vase life of 10 days or more in GLA solution in at least one cultivation method were further selected. The second selection was based mainly on the vase life but also partly on other important traits (e.g., flowers are fully double with no open center at any time, high productivity, not extremely late flowering). Trial production of the lines selected in the second round was conducted to release cultivars at three sites (NIVFS, Akita Agricultural Experiment Station, and Nara Agricultural Research Center) from 2017 to 2019 and at the Kochi Agricultural Research Center from 2018 to 2019, and six lines were finally selected on the basis of their vase life and other important traits.
Vase life evaluation in seedlings and selected linesIn the seedling trials (the first year of each generation), all harvested flowers were evaluated for vase life in the CMIT/MIT solution. The average numbers for vase life surveyed per flowered seedling in the first, second, and third generations were 3.0, 3.4, and 3.3, respectively. The number of days from planting to flowering was recorded, and the diameters of the stem base and stem neck of the first harvested cut flower were measured in the first and second generations with a Vernier caliper.
In the 2017–18 winter–spring season, rooted cuttings of four control cultivars (‘Kamakura’, ‘Kokucho’, ‘Micchan’, and ‘Port Light Pair Beauty’) and six finally selected lines (Table 5) were prepared and planted in 21-cm-diameter pots with 4 L of standard culture soil (Royal Culture Soil; Tachikawa Heiwa Nouen Co., Ltd., Tochigi, Japan) on 31 October 2017. Plants were grown until April in a greenhouse at NIVFS heated above 12°C, with artificial lighting (daylength extension to 14.5 h) using incandescent light bulbs (K-RD110V75W/D; Panasonic Co., Ltd., Osaka, Japan) from 05:00 to 07:00 and from 16:00 to 19:30. Vase life was evaluated from January to April 2018.

Flower vase life of dahlia cultivars and selected first- and second-generation lines in distilled water, antibacterial agent (CMIT/MIT) or GLA solution under standard conditions (23°C, 12-h photoperiod, 70% relative humidity).
In summer and autumn 2018, four control cultivars (‘Kamakura’, ‘Kokucho’, ‘Micchan’, and ‘Port Light Pair Beauty’) and six finally selected lines (Table 5) were grown in the open field at NIVFS as described before, or in a vinyl house with a shade net (cool white, 45%–50% shade; Dio Chemicals, Ltd., Tokyo, Japan) under 50%–55% sunlight (natural daylength, no heating). Rooted cuttings of each cultivar or line were planted 40 cm apart in the open field on 17 May 2018 or in 24 cm-diameter pots with 5 L of the standard culture soil described above in the vinyl house on 24 April 2018. The shoots from each plant were pinched to 2 or 3 nodes on 18 June 2018, and the plants were grown until November following standard production methods (Yamagata, 2018). Vase life was evaluated from September to October 2018.
Statistical analysesThe results shown in Tables 1 and 5 were analyzed with BellCurve for Excel software (Social Survey Research Information Co., Ltd., Tokyo, Japan) using Tukey’s test (P < 0.05).
We found large significant differences in flower vase life among 24 dahlia cultivars (Table 1): ‘Syukuhai’, ‘Rinka’, ‘Micchan’, ‘Super Girl’, ‘Mizou Noir’, ‘Jessy Rita’, ‘Yukitsubaki’, ‘Kokucho’, and ‘Pearl Light’ had long vase life (Index 1); ‘Kyuen’, ‘Moon Waltz’, ‘Kamakura’, ‘Pom-Pom Chocolat’, ‘Agitate’, ‘Benifusya’, ‘Konatsu’, and ‘Black Cat’ had normal vase life (Index 2); and ‘Zuihou’, ‘Dream Waltz’, ‘Shishu’, ‘Red Star’, ‘Gin-ei’, ‘Port Light Pair Beauty’, and ‘Yumesuiren’ had short vase life (Index 3). The responses of the cultivars to the CMIT/MIT and GLA solutions varied (Table 1). Although ‘Yumesuiren’ had the shortest vase life in DW (2.8–3.3 days) in both years, it showed the greatest response to CMIT/MIT (ratio of CMIT/MIT to DW = 1.5). The effect of GLA treatment was strong in ‘Agitate’ (ratio of GLA to DW = 1.9) and in ‘Super Girl’ (1.8).
Crossing and selectionIn all three generations, continuous normal frequency distributions of vase life were observed (Fig. 2). The mean vase life of individual seedlings increased slowly and steadily as generations progressed. Although the frequency of flowers with superior vase life (≥ 8 days) was 1.0% in the first generation and 4.2% in the second generation, it rose to 19.4% in the third generation. The proportion of flowers with inferior vase life (< 4 days) decreased from 29.3% in the first generation to 6.5% in the third generation. The mean vase life was 4.4 days in the first generation, but after two cycles of crossing and selection it increased to 6.1 days (a net total increase of 1.7 days; Fig. 2). The increase was 0.8 days between the first and second generations and 0.9 days between the second and third generations. Therefore, the effect of crossing and selection between generations remained almost constant.

Frequency distributions of flower vase life in the three generations of dahlia breeding populations. Vertical bar represents the mean.
In the first generation, when flowers lost ornamental value, 94.5% showed wilting (Fig. 3). The proportion of flowers with browning (browning plus wilting with browning) increased from 5.4% in the first generation to 19.3% in the third generation. Only 0.1%, 0.9%, and 0% of individual flowers showed petal abscission in the first, second, and third generations, respectively.

Frequency distribution of senescence patterns in the three generations of dahlia breeding populations.
No correlation between vase life and stem base diameter was found (Fig. 4B). However, there was a weak but significant positive correlation between vase life and the number of days from planting to flowering or stem neck diameter (Fig. 4A, C).

Relationships between flower vase life and (A) number of days from planting to flowering, (B) stem base diameter, and (C) stem neck diameter.
In the first generation, the difference between the cross combination with the highest mean vase life (cross no. 533; ‘Kokucho’ × ‘Red Star’; 5.2 days) and that with the lowest mean vase life (cross no. 543; ‘Agitate’ × ‘Yumesuiren’; 3.5 days) was 1.7 days (Table 2). In the second generation, the difference between the cross combination with the highest mean vase life (cross no. 608; 505-47 × 578-4; 7.1 days) and that with the lowest mean vase life (cross no. 624; 544-42 × 541-55; 4.1 days) was 3.0 days (Table 3). In the third generation, the difference between the cross combination with the highest mean vase life (cross no. 823; 609-4 × 631-23; 8.1 days) and that with the lowest mean vase life (cross no. 821; 628-32 × 615-55; 5.1 days) was 3.0 days (Table 4). Tracing back to the origin of each cross combination in the third generation revealed that ‘Micchan’ was a common breeding material in all cross combinations (Table 4).
Vase life of cultivars and selected linesWe examined the vase life of the cut flowers of four control cultivars and six finally selected lines (Fig. 5) in detail in two different seasons and using three different cultivation styles, and found a wide range of variation (Table 5). The mean vase life of ‘Kamakura’, a leading white dahlia cultivar in Japan, was 5.0–6.2 days in DW, 6.0–6.8 days in CMIT/MIT, and 6.0–7.6 days in GLA, whereas in the six finally selected lines it was 6.2–12.0 days in DW, 6.6–10.2 days in CMIT/MIT, and 9.4–13.6 days in GLA (1.4–2.1 times that in ‘Kamakura’). In particular, line 606-46 showed a stably and significantly longer vase life than ‘Kamakura’, except in DW in the vinyl house and CMIT/MIT in the open field (Table 5). The outer petals of the control cultivar ‘Port Light Pair Beauty’ began to wilt after four days (Fig. 6B) and completely lost ornamental value after six days (Fig. 6C), whereas lines 512-2 and 606-46 showed no wilting until eight days (Fig. 6D).

Dahlia cultivar ‘Micchan’ and finally selected first- and second-generation lines with long vase life.

Vase life of two selected lines, 512-2 and 606-46, and the control cultivar ‘Port Light Pair Beauty’ in distilled water. Flowers were photographed at (A) 0, (B) 4, (C) 6, and (D) eight days after harvest (experimental period: 24 April to 2 May 2018). Left: selected first-generation line 512-2; center: ‘Port Light Pair Beauty’; right: selected second-generation line 606-46. The flowers were kept at 23°C, 70% relative humidity, under a 12-h photoperiod.
Since dahlia have long been grown in home gardens rather than commercially, there are no reports of dahlia breeding aimed at improving useful traits such as flower longevity or disease resistance. Here, we report the first attempt to extend the vase life of dahlia flowers using cross-breeding techniques. Two cycles of selection and crossing led to a 1.7-day increase (Fig. 2). The results clearly show that the vase life of dahlia flowers can be extended by crossing and selection.
The seed germination percentage in dahlia crosses was relatively low. For example, the germination rate of the first generation at 67 days after sowing (23 April to 29 June 2015) was 39.3%. The seeds from crosses tended to germinate slowly and gradually, with some seeds germinating two months after sowing. Vivar-Evans et al. (2006) reported that seeds of wild Dahlia coccinea, one of the progenitors of cultivated dahlia in this genus (Okumura and Fujino, 1989), showed physiological dormancy. Dormancy inherited from a wild Dahlia species may be responsible for the low germination rate. In dahlia breeding, it is considered necessary to select an appropriate germination period to take into account seed dormancy.
Although we examined the relationship between vase life and stem base diameter in the seedlings of the first and second generations, according to the recommendations of three Japanese dahlia breeders (see Introduction), we found no correlation between them (Fig. 4B). Harvested flowers were evaluated in CMIT/MIT solution, but not in DW because accidental stem rot was likely to occur in summer. In DW, this correlation may be observed because of a decrease in water absorption due to vascular occlusion. Breeding of dahlias with no hollows or small hollows in the stems to further improve the vase life is our future objective.
Naka et al. (2015) investigated seasonal variations in the vase life in three dahlia cultivars (‘Kamakura’, ‘Kokucho’, and ‘Shukuhai’). The vase life in DW decreased from July to October and increased from December to February, whereas there was little seasonal variation in vase life in GLA solution and it was almost constant throughout the year. In our study, to develop cultivars with a long vase life in summer, when the vase life of dahlia declines, seedlings were selected from July to mid-September. After the initially selected lines were vegetatively propagated, the vase life was examined in detail in two different seasons and using three different cultivation styles (winter–spring in a greenhouse and/or summer–autumn in an open field or vinyl house). This procedure is reliable for selecting lines with a genetically long vase life.
Senescence patterns observed when the flowers lost ornamental value changed gradually with generations (Fig. 3). After two cycles of crossing and selection, the rate of browning senescence patterns (browning plus wilting with browning) increased to 19.3%. In carnation, the relationship between senescence patterns and vase life has been well characterized: petal inrolling at the onset of wilting is a well-known characteristic of ethylene-dependent senescence of normal carnation flowers (Iwazaki et al., 2004; Otsu et al., 2007; Satoh, 2011). In contrast, desiccation and browning of petals are characteristics of ethylene-independent senescence in carnation cultivars with low ethylene production. In gerbera breeding for long vase life, changes in the proportion of senescence patterns (bending, folding, and wilting) were also observed (Wernett et al., 1996a). Before breeding, gerbera flowers senesced due to bending rather than wilting. After only one cycle of selection and crossing, the proportion shifted dramatically from bending to wilting (Wernett et al., 1996a). We plan to continue studying the relationship between the senescence patterns and vase life using future generations in dahlias.
Dahlia flowers come in various flower types, such as straight cactus, incurved cactus, water lily, and formal decorative (Okumura and Fujino, 1989). The breeding materials shown in Table 1 had five different flower types. However, the finally selected lines had only two types, formal decorative and semi-cactus (Fig. 5; Table 5). The flower type of 58 lines initially selected in the third generation was almost exclusively formal decorative (data not shown). Therefore, a loss of flower type diversity was caused by selection and crossing for long vase life. Further studies are needed to clarify the relationship between flower type and flower vase life.
As mentioned in the Introduction, Tsujimoto et al. (2016b) reported that ‘Rinka’, ‘Syukuhai’, ‘Moon Waltz’, ‘Benifusya’, ‘Micchan’, ‘Akebono-Temari’, and ‘Pink Sapphire’ have very long vase life. Azuma et al. (2019) reported that the vase life of outer florets in DW was longer in ‘Kokucho’, ‘Micchan’, and ‘Moon Waltz’ than in seven other cultivars examined. The vase life of cut dahlia flowers is affected by bacterial proliferation in the vase solution and carbohydrate level in petals (Azuma et al., 2019). Furthermore, we found that the CMIT/MIT treatment extended the vase life of three cultivars, all of which had relatively high numbers of bacteria in their vase solutions (Azuma et al., 2019). Ichimura et al. (2011) showed that continuous treatment with GLA effectively extended the vase life of ‘Kokucho’ dahlia flowers. Therefore, we used three different solutions (DW, CMIT/MIT, and GLA) over two years (2014 and 2015), and found that ‘Syukuhai’, ‘Rinka’, and ‘Micchan’ were the top three Index 1 cultivars (Table 1), in agreement with the result of Tsujimoto et al. (2016b), although ‘Moon Waltz’ and ‘Benifusya’ were classified as Index 2 (normal vase life), unlike in Tsujimoto et al. (2016b).
Among 24 cultivars, the effect of GLA was highest in ‘Agitate’ and ‘Super Girl’ (Table 1). In a previous study, co-treatment with glucose and CMIT/MIT significantly increased the fresh weight of ‘Agitate’ flowers because the carbohydrate content in their petals was particularly low (Azuma et al., 2019). These results indicate that low levels of carbohydrates in ‘Agitate’ may have contributed to the strong effect of GLA on vase life.
We found a close relationship between the cross combination and flower vase life. In the first generation, the difference between the cross combination with the highest mean vase life and that with the lowest mean vase life was 1.7 days (Table 2). However, there was no relationship between the vase life of the parent cultivar and that of the progeny. In the second generation, the difference between the cross combination with the highest mean vase life and that with the lowest mean vase life was 3.0 days (Table 3). ‘Dream Waltz’, ‘Port Light Pair Beauty’, and ‘Yumesuiren’ (all Index 3; Table 1) were used to breed parental lines of four cross combinations with the shortest vase life (crosses no. 618, 621, 625, and 624; Table 3). In the third generation, ‘Micchan’ (Index 1; Table 1) was used to breed parental lines of all cross combinations. The six finally selected lines with long vase life (Table 5) were all progeny of ‘Micchan’.
For ‘Syukuhai’ and ‘Rinka’ (top two Index 1 cultivars; Table 1), 55 seeds were obtained in eight of the 11 cross combinations in which these two cultivars were used as seed parents or pollen parents (data not shown). All obtained seeds were sown, but the germination rate was low. Only six seedlings were evaluated for vase life: three from cross no. 558 (‘Pom-Pom Chocolat’ × ‘Rinka’), two from cross no. 503 (‘Yukitsubaki’ × ‘Rinka’), and one from cross no. 553 (‘Jessy Rita’ × ‘Syukuhai’). The mean vase life of these six seedlings was 3.0–4.0 days, and none of them were selected initially (Table 2).
In each generation, we selected lines with long vase life for use as breeding materials for the next generation. Plants were not initially selected for any other characteristics. Our results strongly suggest that ‘Micchan’ has genes related to long flower vase life and that the trait is heritable. ‘Micchan’, a pink ball-type cultivar (Fig. 5) bred by the famous Japanese dahlia breeder Koji Washizawa in 2005, had the largest market share in 2017 at the Ota Floriculture Auction Co., Ltd., the biggest flower market in Japan (Onozaki, 2018b). One of the major factors that allowed this cultivar to dominate the market seems to be its long vase life.
In Japanese morning glory (Ipomoea nil (L.) Roth), a NAC (NAM/ATAF1,2/CUC2) transcription factor, EPHEMERAL1 (EPH1), is a key regulator of ethylene-independent petal senescence (Shibuya, 2018; Shibuya et al., 2014). As dahlia flowers also show ethylene-independent petal senescence, an EPH1 homolog may be involved in petal aging in dahlia. Although there was no correlation between vase life and stem base diameter (Fig. 4), Tsujimoto et al. (2016a) reported a highly significant positive correlation between cell density in the second nodes of the stem cross-section and flower vase life. We plan to measure cell densities in our selected lines with long vase life to clarify the relationship between these traits.
In conclusion, using conventional cross-breeding techniques, we extended the flower vase life of dahlia flowers and developed six lines with genetically determined long vase life. We are currently evaluating other important characteristics in these lines at four sites in Japan (NIVFS, Akita Agricultural Experiment Station, Nara Agricultural Research Center, and Kochi Agricultural Research Center) with the aim of releasing them as high-value-added pilot cultivars in the near future.
We sincerely thank Dr. M. Mato, Ms. A. Yamagata (Akita Agricultural Experiment Station), Dr. T. Naka, Mr. T. Nakajima, Mr. K. Inda (Nara Agricultural Research Center), Mr. A. Kataoka, and Mr. S. Yamashita (Kochi Agricultural Research Center) for their contributions to the trial production of selected lines. We also thank the members of the Tsukuba Technical Support Center, Fujimoto-Owashi Operations Unit, NARO, for their technical support in field and greenhouse management.