Abstract
In the present study, we evaluated the effects of intermittent low temperature storage (ILTS) following cold imbibition of Eustoma seeds as a low-cost method. The objective of this study was to investigate the effects of the total number of cold storage days and the lengths of ILTS cycles for seedlings on the subsequent growth and flowering of three cultivars with different susceptibilities to rosetting: ‘Nancy’, ‘Exe Lavender’, and ‘Bolero White’. Before sowing, the seeds were imbibed with water at 10°C under a dark condition for 35 days. A cycle of 6D/6D (6 days in a refrigerator kept at 10°C in the dark/6 days in a greenhouse kept at a high ambient temperature) was repeated 0, 1, 2, 3, 4, and 5 times to attain 0, 6, 12, 18, 24, and 30 days of cold storage (DCS), respectively. Using a cycle of 6D/6D at least one time (6 DCS) increased the bolting and flowering percentages and accelerated the bolting date by 6–10 days and the flowering date by 4–9 days in ‘Nancy’, a high-rosetting cultivar. The percentage of harvesting was above 80% only in seedlings subjected to 24 and 30 DCS in ILTS. In addition, under equal 30 DCS of ILTS, cycles of 3D/3D × 10 times, 6D/6D × 5 times, and 15D/15D × 2 times were investigated by exposing the seedlings to a high ambient temperature for 3, 6, and 15 days and then transferring them to a refrigerator (10°C) for 3, 6, and 15 days, respectively. ILTS cycles of 3D/3D and 6D/6D resulted in earlier bolting and flowering dates than cycles of 15D/15D. The cycle of 15D/15D resulted in a lower percentage of marketable harvesting and an increased number of nodes to the first flower. These results suggest that applying ILTS with a cycle of 6D/6D at least four times can be an efficient method to promote the bolting and flowering of Eustoma without applying air conditioning to the entire greenhouse.
Introduction
Eustoma (Eustoma grandiflorum L.) is widely grown for the production of potted flowers and cut flowers throughout the world. In Japan, the demand for Eustoma cut flowers is especially high during the New Year holidays. In order to produce cut flowers for this season, the plants need to be sown in late July to early August and transplanted in early September. In Eustoma, rosetting easily occurs in young seedlings following exposure to high temperatures (Harbaugh et al., 1992; Li et al., 2002; Ohkawa et al., 1991). To prevent rosetting, cold imbibition of Eustoma seeds has been widely used recently (Fukushima et al., 2017; Kageyama et al., 1990; Tanigawa et al., 1999).
Although imbibing seeds under low temperatures has become a very common practice, increasing summer temperatures caused by global warming is unfavorable for the growth of seedlings of various high-rosetting Eustoma cultivars in southwestern Japan. For this reason, even during the cultivation of cold-imbibed seeds, it is necessary to cool the temperatures of greenhouses (25°C/15°C, day/night) using air conditioners in the summer to harvest qualified cut flowers up to the end of the year. However, the installation and energy consumption to maintain air conditioning systems throughout the period of seedlings growth require are expensive. Therefore, the development of alternative cost-effective procedures to ensure stable flowering and good seedling growth is needed.
Recently, intermittent low temperature storage (ILTS), in which plants are repeatedly transferred between a refrigerator (low temperature) and ambient conditions (high temperature), was reported as a useful and low-cost procedure for flower induction of June-bearing strawberries (Yoshida et al., 2012). In this study, it was found that in plants subjected to ILTS, the flowering was earlier and more uniform compared to strawberry plants grown under a continuous dark cold storage condition. The supply of assimilates due to outdoor photosynthesis during ILTS may be an important factor. Furthermore, the flowering of potted plants was found to be accelerated by applying ILTS in Begonia × hiemalis ‘Netja’ (Nakajima and Goto, 2018). During ILTS in these plants, effects of the application of low temperatures, high temperature avoidance, and short-day have been reported (Nakajima and Goto, 2018; Yoshida et al., 2012). In Eustoma, previous studies reported that the plants need a low temperature environment after sowing or planting to enhance the rate of stem elongation and bolting speed (Pergola, 1992; Takeda, 1995). With the aim of producing good quality seedlings in the summer and producing cut flowers for the end of the year, ILTS could be an efficient method for growing cold-imbibed Eustoma seeds. However, application of ILTS has not yet been examined in Eustoma. In the present study, we investigated the effects of the total number of cold storage days and the lengths of ILTS cycles during the growing period of Eustoma seedlings on the subsequent bolting and flowering of cultivars with different susceptibilities to rosetting.
Materials and Methods
General
ILTS was conducted from sowing (July) to transplanting (September). Eustoma seeds were imbibed with distilled water at 10°C under a dark condition for 35 days and then sown in cell trays (288-cell size) before beginning the ILTS procedure. ILTS was performed with a xD/xD cycle × repeated time in which the seedlings were rotationally transported between a refrigerator (10°C) and a greenhouse at an ambient temperature of a set number of days (x days). The cycle was repeated several times to complete the total number of cold storage days for each experiment (Fig. 1). During the treatment time in the greenhouse (ambient day length and high ambient temperature), seeds were placed under mist irrigation to germinate until the cotyledons expanded completely. Then, the seedlings were supplied daily with OAT A solution (N, 17.7 mM; P, 1.7 mM; K, 7.8 mM; Ca, 4.2 mM; Mg, 1.86 mM; OAT Agrio Co., Ltd., Japan) at 1/6 strength and by sub-irrigation until transplanting while in the greenhouse. When moved to a refrigerator, the seedlings were stored at 10°C without lighting or irrigation. When ILTS was completed, six seedlings were transplanted to a planter (64 cm × 22 cm × 18 cm, soil capacity 15.0 L) as one replication and there were three replications per treatment. The planters of transplanted seedlings were placed in a greenhouse kept at 15°C minimum for 16 h of day length (natural day length with additional lighting by an incandescent lamp from 4:00 to 9:00 and from 16:00 to 20:00). Ten days after transplanting, 1.5 liters of 1/2 strength OAT A solution was applied to each planter once a week. The air temperature in the greenhouse was recorded every 5 min using a datalogger (Ondotori TR-71Ui; T&D Corporation, Japan).

The bolting date and bolting node were recorded when the length of an internode exceeded 5 mm. Rosetting was determined when the plant did not bolt 60 days after transplanting. The dates of flowering (when the first flower opened) was recorded for all plants except the rosetted ones. Flowering shoots were harvested when the fourth flower opened and cut flower traits were investigated such as fresh weight and length, the number of nodes to the first flower, and the total numbers of visible flower buds and flowers. Cut flowers with more than a total of six flowers including flower buds were used to calculate the marketable harvesting percentage. The mean values of each replication were subjected to one-way ANOVA and means were compared using Tukey’s HSD test (Excel-toukei 2010; Social Survey Research Information Co., Ltd., Japan).
Experiment 1. Effect of total duration of low temperature storage applied intermittently
In this experiment, a mild-rosetting cultivar, ‘Exe Lavender’ (Kaneko Seeds Co., Ltd., Japan), and a high-rosetting cultivar, ‘Nancy’ (Fukukaen Nursery & Bulb Co., Ltd., Japan), were examined in 2018. In our preliminary experiments, the rate of rosetting was 44.4% in ‘Exe Lavender’ and 92.9% in ‘Nancy’ under a natural high temperature season (unpublished data). After the end of cold imbibition, seeds of both cultivars were sown and placed in a greenhouse for three days. After that, the seeds were placed in a refrigerator for six days, and then moved to a greenhouse for six days (cycle of 6D/6D). The cycle of 6D/6D was repeated 0, 1, 2, 3, 4, and 5 times to attain 0, 6, 12, 18, 24, and 30 days of cold storage (DCS), respectively. The Eustoma cold-imbibed seeds were sown in 2018 on July 3, 9, 15, 21, 27, and August 2 depending on the duration of low temperature storage (Fig. 1). The duration of high ambient temperature exposure was the same for all the treatments at 33 days. Seeds of the control (without any low temperature treatment) were also examined and sown on August 2. All seedlings were transplanted on September 4.
Experiment 2. Effect of intermittent low temperature storage cycles
A low-rosetting cultivar ‘Bolero White’ (Miyoshi Co. Ltd., Japan) and the high-rosetting cultivar ‘Nancy’ were examined. In the preliminary experiment, the rate of rosetting was 7.1% in ‘Bolero White’ under a natural high temperature season (unpublished data). Before ILTS, the seeds of both cultivars were imbibed in water at 10°C on May 27, 2013, until sowing. After sowing, the seeds were placed in a greenhouse for 3, 6, and 15 days and then placed in a refrigerator for 3, 6, and 15 days, respectively. ILTS cycles × repeated time of 3D/3D × 10 times, 6D/6D × 5 times, and 15D/15D × 2 times were investigated (Fig. 1). The total exposure to low temperature and high-temperature conditions were designed to be the same in all treatments of 30 days and 33 days, respectively. 0D seeds were imbibed at 10°C on June 26, 2013, and sown on July 31, but not subjected to ILTS. All seedlings were transplanted on September 2.
Results
Experiment 1. Effect of total duration of low temperature storage applied intermittently
During the seedling growth in 2018 (July–September), the daily mean temperature ranged from 24.3°C to 34.1°C; it was mostly higher than 30°C in the period July 13–August 14 (except for August 7–8) and August 21–30. The average daily minimum temperature, daily mean temperature, and daily maximum temperature were 25.0°C, 30.7°C, and 38.3°C, respectively (Fig. 2).

Treatments with ILTS that started three days after sowing the cold-imbibed seeds, accelerated bolting by 3–7 days in ‘Exe Lavender’ and by 6–10 days in ‘Nancy’ compared to no ILTS treatment (0 DCS) (Table 1). In a mild-rosetting cultivar, ‘Exe Lavender’, 88.9% of control plants started rosetting whereas most of cold treatment plants resulted in bolting. Although there was no significant difference in the bolting date or flowering date, ILTS treatments only affected the flowering percentage. No ILTS treatment using only cold imbibition of seeds (0 DCS) improved the induction rate to 94.4% bolting, but induced only 50.0% of the total plants to flower by December 20 (Table 1). Most ILTS treatments resulted in 100% bolting (excepted for the 12 DCS treatment) and over 70% flowering. Twenty-four and 30 DCS resulted in a significantly higher percentage of flowering compared with 0, 6, and 12 DCS (Table 1). Moreover, plants grown in 24 and 30 DCS started to flower and reached a half of the total flowering rate earlier than those grown using shorter lengths of cold storage (Fig. 3). In the high-rosetting cultivar, ‘Nancy’, all control plants formed rosettes. Although the 0 DCS treatment and other durations of ILTS induced 94.4% and 100% bolting, respectively, the bolting date of the 0 DCS treatment was 7–10 days later than that of plants treated more than 12 days (Table 1). The percentage of flowering was greatest and the number of days to flowering was lowest in 24 DCS and 30 DCS (Table 1). Plants grown in 24 and 30 DCS achieved 83% flowering on December 7, while those grown in 6, 12, and 18 DCS showed a flowering rate lower than 60% on the same day (Fig. 4); however, there was no significant difference in flowering in the 12–30 DCS treatments.


The duration of low temperature storage affected the percentage of harvesting until December 31 in both ‘Exe Lavender’ and ‘Nancy’ (Table 2). By increasing the low temperature days to 30 days, the percentage of harvesting increased markedly to over 80%. However, there was no significant difference in cut flower length, number of flowers or visible flower buds in treatments for ‘Exe Lavender’. The number of nodes to the first flower was largest in the 0 DCS treatment (10.3). In ‘Nancy’, the highest percentage of harvesting and the longest cut flower length were both with 30 DCS. There was no significant difference in the number of nodes to the first flower, cut flower weight, or number of flowers and visible flower buds among ILTS treatments of this cultivar.
Experiment 2. Effects of intermittent low temperature storage cycles
During the seedling growth in 2013 (July–September), the daily mean temperature ranged from 20.1°C to 32.1°C; it was mostly higher than 25°C and decreased to 23°C in early September. The average daily minimum temperature, daily mean temperature and daily maximum temperature were 24.3°C, 28.9°C, and 35.5°C, respectively (Fig. 5).

In a low-rosetting cultivar, ‘Bolero White’, 100% bolting and flowering were achieved in all treatments, but the bolting date and flowering date for ILTS cycles of 3D/3D and 6D/6D were earlier than those of 0D and 15D/15D (Table 3). In the high-rosetting cultivar ‘Nancy’, 100% bolting was also observed in all treatments and 100% flowering was observed in all plants treated with ILTS, whereas plants grown without ILTS resulted in only 66.7% flowering (Table 3; Fig. 6). Seedlings grown under ILTS cycles of 6D/6D and 3D/3D generally bolted and produced flowers in a similar manner. Their bolting and flowering were earlier than those of the 0D treatment by 11 days and 23–25 days, respectively. Flowering in 15D/15D was delayed by 13–14 days compared with 6D/6D and 3D/3D (Table 3; Fig. 6).

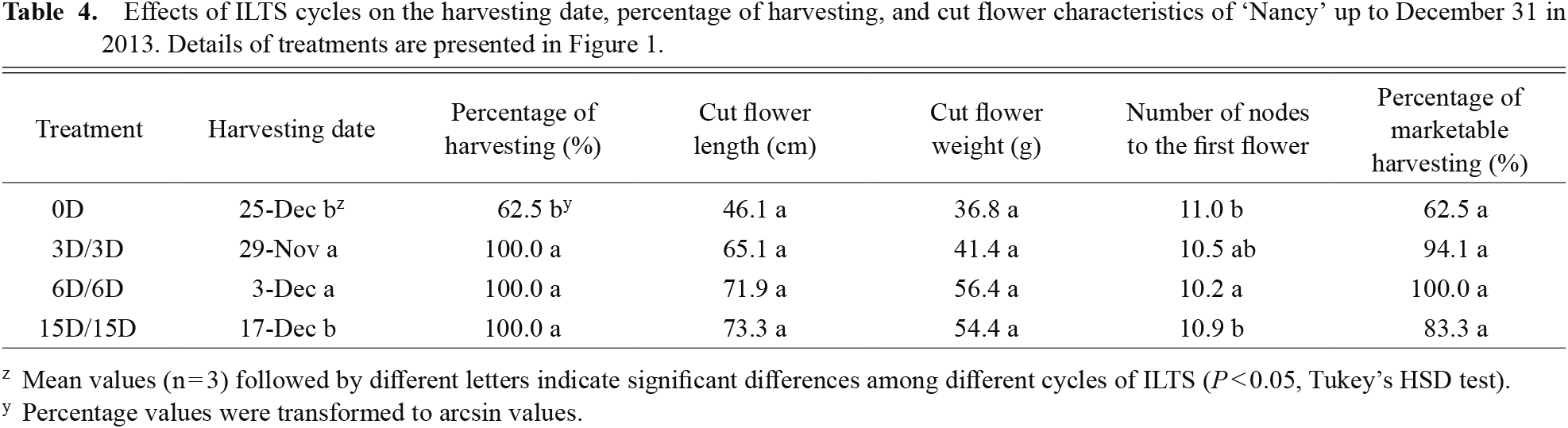
Table 4 shows the effects of ILTS cycles on cut flower traits of ‘Nancy’ up to December 31. Only 62.5% of seedlings without ILTS produced cut flowers, whereas all seedlings treated with ILTS produced cut flowers up to December 31. However, only the cycle of 6D/6D achieved 100% marketable harvesting. There was no significant difference in the fresh weight or length of cut flowers among treatments. The number of nodes to the first flower in ILTS cycles of 6D/6D was smaller than that in the 15D/15D cycle in ‘Nancy’. There was no significant difference in these values among treatments in ‘Bolero White’ (data not shown).
Discussion
To produce Eustoma cut flowers for the New Year holidays in the warm southwestern area in Japan, it is necessary to successfully grow seedlings in the summer. Cold imbibition of the seeds of Eustoma can prevent rosetting, but does not induce flowering sufficiently and cut flower harvesting may be limited up to the end of the year if the greenhouse temperature is not actively cooled while raising the seedlings in the summer. Accordingly, in this study, we investigated whether flowering, cut flower harvesting, and cut flower quality could be improved using intermittent low temperature storage (ILTS) in several cultivars with different susceptibilities to rosetting, all of which had their seeds pre-treated with cold imbibition.
In the experimental design, the total number of cold storage days and the lengths of ILTS cycles were determined as follows. In traditional commercial cultivation, Eustoma rosetted seedlings are placed in a refrigerator at 10°C for 30–35 days to break rosetting (Ninomiya et al., 1997). During this time, irrigation is performed once a week. In the ILTS procedure, irrigation is not applied while the seedlings are stored at low temperatures under a dark condition. Therefore, in Experiment 1, we decided to store seedlings at low temperatures intermittently every six days, up to 30 days.
As the minimum temperature during the seedling stage was over 25°C (Fig. 5), plant growth and flowering suppression were likely to occur. Seedlings not stored at low temperatures (control) rarely bolted in ‘Exe Lavender’ and ‘Nancy’ because young Eustoma seedlings, especially just after germination, are very sensitive to high temperatures and rosetting is easily induced (Harbaugh et al., 1992; Ohkawa et al., 1991). Most seedlings bolted when seeds were subjected to 10°C imbibition for 35 days before sowing. However, it was found that only half of the plants grown from cold-imbibed seeds without ILTS treatment flowered up to December 20. Similarly, the effect of high temperature on reducing bolting can be avoided by other low temperature treatments such as artificial night cooling, low ambient temperatures, or planting in highlands; however, flowering was not induced in all plants that bolted (Ohkawa et al., 1996). Therefore, we confirmed that applying only cold imbibition to Eustoma seeds was insufficient to stimulate flowering after stem elongation when growing them at high ambient temperatures.
The total number of cold storage days depended on the number of cycles of 6D/6D in ILTS. The seedlings treated with ILTS after cold imbibition of the seeds resulted in early bolting and flowering and a higher percentage of flowering in both ‘Exe Lavender’ and ‘Nancy’. Nevertheless, it was found that flowering was affected by the duration of low temperature storage applied intermittently after the cold imbibition of the seeds; a longer duration of low temperature exposure (more than 18 days) increased the percentage of flowering. The induced flowering was maximum (100%) in both 24 and 30 DCS. In both ‘Exe Lavender’ and ‘Nancy’, the first flower buds of 24 and 30 DCS opened earlier than those of 6, 12, and 18 DCS (Figs. 3 and 4). Thus, length of low temperature storage has a great impact on bolting and flowering timing of Eustoma.
When the high-rosetting cultivar ‘Nancy’ was exposed to low temperatures for 30 days, the percentage of cut flowers reached 100% before December 31 and the longest fresh cut flower was achieved (Table 2). The percentage of flowering was significantly lower in 6, 12, and 18 DCS than 30 DCS. Thus, it is possible that the effect of low temperature exposure after sowing can be mitigated by the duration of high ambient temperature exposure. A similar result was found in a previous study where a shortened duration of cold imbibition (from five weeks to one week) reduced the bolting rate of Eustoma sown under a high ambient temperature (Tanigawa et al., 2002). In order to control the flowering effectively, as well as harvesting high-rosetting Eustoma cultivars, cold exposure with ILTS should be 24 days or longer. This will enable growers to produce Eustoma cut flowers in the winter without applying air conditioning to the entire greenhouse in the summer; therefore, applying ILTS can be considered as a low-cost method.
Furthermore, we examined the lengths of ILTS cycles when the total number of cold storage days was 30 days. No significant difference was observed between 3D/3D and 6D/6D in bolting and flowering. In this experiment, the percentages of flowering and harvesting were 100% in all the plants treated with ILTS. If the total low temperature duration of ILTS is the same, there may be no difference in the effects when the treatment cycle is changed between three days to six days. As such, it is interesting that the effectiveness of continuous low temperature exposure on Eustoma seedlings could be divided into brief durations while the total amount of low temperature exposure was the same. This finding was consistent with the report of Nakajima and Goto (2018) who reported that the day of blooming after ILTS cycles of 2D/2D × 8 times was similar to that of cycles of 4D/4D × 4 times in Begonia × hiemalis ‘Netja’. Continuous low temperature exposure was also effective in inducing flowering in rosetting cultivars of Eustoma after transplanting at four weeks (Ohkawa et al., 1994; Pergola, 1992); however, no report has examined the potential of applying interrupted low temperature storage for growing seedlings. This is the first report to show that bolting and flowering of Eustoma can be achieved successfully under constant interruption of low temperature exposure.
Even though the duration of low temperature storage was applied for the same 30 DCS, the cycle of 15D/15D was less effective for Eustoma compared to cycles of 3D/3D and 6D/6D. Seedlings grown under ILTS with 15D/15D showed significantly later bolting, flowering, and harvesting than with cycles of 3D/3D and 6D/6D, and there was a lower percentage of marketable harvest in ‘Nancy’. The starting time of low temperature exposure for seedlings was too late in 15D/15D and 15 days of ambient high temperature may have negatively affected development (Fig. 1). Murata et al. (1990) reported that flowering of Eustoma was delayed when night air cooling was applied two weeks after sowing compared to that applied immediately after sowing. In another report, the percentage of flowering in Eustoma ‘GCREC-Blue’ decreased when the young seedlings were exposed to high temperature (Harbaugh, 1995). These results suggest that the low temperature effect obtained during seed cold-imbibition may be reduced by high temperature exposure immediately after sowing. The start time of ILTS in the cycle of 15D/15D should be investigated to see if it affects the bolting and flowering ability of Eustoma.
This report suggests that the negative effects of high ambient temperatures can be reduced by applying ILTS after sowing of cold-imbibed seeds instead of using air conditioners to cool down the temperatures of greenhouses. The duration of low temperature exposure should be 24 days or longer using 6D/6D of ILTS at 10°C to improve the flowering rate and cut flower yield at the end of the year.
Literature Cited
- Fukushima, K., S. Kajihara, S. Ishikura, N. Katsutani and T. Goto. 2017. Effect of exposing imbibed seeds to low temperatures on the growth and characteristics of Eustoma grandiflorum. Hort. Res. (Japan) 16: 177–184 (In Japanese with English abstract).
- Harbaugh, B. K. 1995. Flowering of Eustoma grandiflorum (Raf.) Shinn. cultivars influenced by photoperiod and temperature. HortScience 30: 1375–1377.
- Harbaugh, B. K., M. S. Roh, R. H. Lawson and B. Pemberton. 1992. Rosetting of lisianthus cultivars exposed to high temperature. HortScience 27: 885–887.
- Kageyama, Y., Y. Fukushima and K. Konishi. 1990. Effects of raising seedling at cool temperature and chilling treatment of seed on rosette formation of Eustoma. J. Japan. Soc. Hort. Sci. 59 (Suppl. 1): 496–497 (In Japanese).
- Li, J., Y. Notsu, I. M. Ogawa, H. Ohno and K. Ohkawa. 2002. Rosetting characteristics-based classification of Eustoma grandiflorum (Raf.) Shinn. cultivars sown on different dates. Environ. Control Biol. 40: 229–237.
- Murata, T., Y. Iwakiri and H. Takahashi. 1990. Effects of cooling treatment in night with seedling of Eustoma russellianum on growth and flowering. Kyushu Agricultural Research 53: 202 (In Japanese).
- Nakajima, T. and T. Goto. 2018. Effect of intermittent low temperature storage on flowering and growth acceleration of Begonia × hiemalis ‘Netja.’ Hort. Res. (Japan) 17: 73–78 (In Japanese with English abstract).
- Ninomiya, C., K. Takano and A. Azuma. 1997. Effect of duration of the temperature treatment on growth and development in greenhouse controled at low night temperature after planting. Bull. Kochi Agric. Res. Cent. 6: 31–36 (In Japanese with English abstract).
- Ohkawa, K., A. Kano, K. Kanematsu and M. Korenaga. 1991. Effects of air temperature and time on rosette formation in seedlings of Eustoma grandiflorum. Sci. Agric. 48: 171–176.
- Ohkawa, K., S. Yamaguchi, M. Miyoshi and A. Yamazaki. 1996. Forcing of rosetted Eustoma grandifforum seedlings by low temperature treatments. Environ. Control Biol. 34: 45–52.
- Ohkawa, K., T. Yoshizumi, M. Korenaga and K. Kanematsu. 1994. Reversal of heat-induced rosetting in Eustoma grandiflorum with low temperatures. HortScience 29: 165–166.
- Pergola, G. 1992. The need for vernalization in Eustoma russellianum. Sci. Hortic. 51: 123–127.
- Takeda, T. 1995. Effects of low temperature treatments on bolting and flowering of rosetted Eustoma Seedlings. J. Japan. Soc. Hort. Sci. 64: 356–366 (In Japanese with English abstract).
- Tanigawa, T., Y. Kobayashi and T. Kunitake. 1999. Low temperature treatment of imbibed seeds and varietal differences in the rate of bolting and flowering under high-temperature condition in Eustoma grandiflorum. J. Japan. Soc. Hortic. Sci. 68 (Suppl. 2): 378 (In Japanese).
- Tanigawa, T., N. Kuroyanagi and T. Kunitake. 2002. Effects of low temperature treatment of imbibed seeds of Eustoma grandiflorum (Raf.) Shinn. on their germination and subsequent bolting. J. Japan. Soc. Hort. Sci. 71: 697–701 (In Japanese with English abstract).
- Yoshida, Y., E. Ozaki, K. Murakami and T. Goto. 2012. Flower induction in June-bearing strawberry by intermittent low temperature storage. J. Japan. Soc. Hort. Sci. 81: 343–349.