ORIGINAL ARTICLES
Relationship Between Shoot Growth and Fruit Production of ‘Masui Dauphine’ and ‘Houraishi’ Fig Trees at Various Plant Densities
2020 Volume 89 Issue 4 Pages 375-383

Details
2020 Volume 89 Issue 4 Pages 375-383
This study aimed to determine the optimal shoot vigor to produce large fruit and high yields of the fig (Ficus carica L.) cultivars ‘Masui Dauphine’ and ‘Houraishi’. Fruit production of the two cultivars was compared for seven years on shoots of various vigor induced by different plant densities: 20, 61, 400, 667, and 2000 trees per 1000 m2, and all shoots occupied equal-sized basal areas (0.5 m2 for each shoot). Individual shoot weight (shoot weight) and number of superfluous shoots were greater in the trees grown at higher plant density. The greater shoot growth under higher plant density was consistent for seven years for ‘Masui Dauphine’, but such an increase was suppressed after the third year for ‘Houraishi’ trees. The relationship between fruit production and shoot weight showed that fruit maturation did not last until late autumn and the number of fruits harvested (fruit number) declined on the shoots smaller than approximately 0.15 kg on ‘Masui Dauphine’ trees and by approximately 0.8 kg in ‘Houraishi’ trees. In early autumn, however, the fruit number was larger on the smaller shoots than that on the larger shoots in ‘Houraishi’ trees. The annual means of individual fruit weight (fruit weight) were negatively correlated with shoot weight from the third to seventh years for ‘Houraishi’ trees, but only for the fifth year for ‘Masui Dauphine’ trees. The fruit yield per shoot (fruit yield) was positively correlated with shoot weight based on the fruit number. However, the correlation coefficients between fruit yield and shoot weight were lower in ‘Houraishi’ trees because the fruit weight was negatively correlated with the shoot weight. Relatively large shoots in a compact tree area will be useful for ‘Masui Dauphine’ figs to avoid fruit yield decline. Conversely, smaller shoots in a large tree area will be useful for ‘Houraishi’ figs to produce more and larger fruit in early season of harvesting.
In the Japanese fig industry, ‘Masui Dauphine’ (‘San Piero’ sensu; Condit, 1955) and ‘Houraishi’ (‘Horaigaki’ sensu; Condit, 1955) are two major common-type fig cultivars, and the second crops, which are borne on every node of the shoot and are harvested from summer to autumn, are sold as fresh fruits. Previous studies (e.g. Awamura et al., 1993; Senda et al., 1996) and farmers’ experience suggest that fruit production is correlated with shoot vigor, which is greatly affected by the number of shoots and their distribution on a tree. Methods of tree training, pruning shoots or managing plant density have been selected to induce such suitable shoot vigor in each cultivar.
‘Masui Dauphine’ is a moderately vigorous cultivar (HPACEEE, 1982), typically trained either by base-form training (traditional) or straight-line training (improved). The base-form training involves various branched structures and the distribution of the mother shoots in a planar space. The straight-line training involves bilateral horizontal limbs (primary scaffold limbs), which greatly improves the cultivation efficiency, because the shoots elongate in line in a regular fashion (Kabumoto et al., 1985). In both of the training methods used for the ‘Masui Dauphine’ cultivar, a high plant density (about 100 trees and 2500 shoots per 1000 m2) and short pruning are applied (Mano, 2015), and relatively vigorous shoots are recommended in a range of basal diameters (from 18 to 23 mm) for good fruit production (Senda et al., 1996). Furthermore, extremely dense planting (e.g. about 625 trees and 2800 shoots per 1000 m2) is recommended for early production and early recovery from tree injury (Kadowaki et al., 2010; Mano and Hamada, 2005; Mano et al., 2011).
‘Houraishi’ is a vigorous cultivar (HPACEEE, 1982), typically trained by open-center training, which involves pruning to thin-out shoots. A very low plant density (from 10 to 20 trees per 1000 m2) and many short (0.2–0.4 m) shoots (about 5000 shoots per 1000 m2) are favorable for fruit setting and coloration and this has been typical for this training method (Awamura, 2015; Awamura et al., 1993). Recently, straight-line training with relatively denser planting (from 50 to 80 trees and 2400 to 2500 shoots per 1000 m2), has also been adopted for ‘Houraishi’ fig (APDYPG, 2017) because of its cultivation efficiency. In contrast to open-center training, the short pruning and denser planting of this training are thought to enable excessive shoot growth; however, it is unknown how this excessive shoot growth affects fruit production.
As described above, some cultivation methods are used or recommended for both cultivars. However, these recommendations have not been validated uniformly by the relationship between shoot size and fruit growth. The fruit production industry is selective for various traits such as yield, fruit size, skin color, and flesh taste. Large fruit size and high yield, traits especially important for commercial production, were the primary reasons for the wide cultivation of ‘Masui Dauphine’ and ‘Houraishi’ fig varieties in Japan. In this study, the relationship among individual shoot growth, fruit size, and fruit yield was analyzed with various levels of shoot vigor resulting from various plant densities. The applicability of conventional and newly adopted cultivation methods for both fig cultivars was evaluated in terms of controlling the shoot vigor.
A field experiment was conducted at the Research Institute of Environment, Agriculture and Fisheries, Osaka Prefecture, (34°32'N; 135°36'E, 70.5 m a.s.l) in Osaka, Japan. As shown in Table 1, nursery stocks of ‘Masui Dauphine’ and ‘Houraishi’ fig trees were planted in the summer of 2004 in five different groups consisting of 1, 1, 4, 5, and 6 trees with tree spacing of 20.0, 6.6, 1.0, 0.6, and 0.2 m, respectively. These groups were randomly arranged in 2.5 m × 35 m (North–South) rows with extra trees at row terminals; the order of tree spacing accommodated plant densities of 20, 61, 400, 667, and 2000 trees per 1000 m2, respectively. Initially, the experiment comprised 10 rows (eight rows for ‘Masui Dauphine’ and two rows for ‘Houraishi’) located between the extra rows. Two of the ‘Houraishi’ rows were used for the seven years of this study; however, some of the ‘Masui Dauphine’ rows were used for another study, so the number of these rows (= the number of groups for each density) decreased to five in the third year and to two rows thereafter. The numbers of tested trees used in ascending order of the plant densities of ‘Masui Dauphine’ were 8, 8, 32, 40, and 48 for the first two years; 5, 5, 20, 25, and 30 for the third year; and 2, 2, 8, 10, and 12 for the remaining years. The numbers of ‘Houraishi’ trees were 2, 2, 8, 10, and 12 for all seven years.

Design of the section for plant density test in ‘Masui Dauphine’ and ‘Houraishi’ fig trees.
All trees received straight-line training (Kabumoto et al., 1985), with upward shoots from a 40 cm high horizontal limb, as shown in Figure 1. The mother shoots were arranged alternately on the primary scaffold limbs with equal intervals of ca. 0.2 m. The numbers of mother shoots were 100, 33, 5, 3, and 1 per tree, in ascending order of the plant densities. One shoot per mother shoot was elongated diagonally upward without pinching. As a result, every shoot of each tree occupied an equivalent area of 0.5 m2 (2500 shoots per 1000 m2). In the following February, every shoot was pruned above the second or third node from the base and used as the mother shoot the next season. The measurements described in the next section began from the year in which the training of each density group was completed. In addition, other methods of cultivation, such as irrigation, fertilization, and pest management, followed the normal schedule for fig orchard management.

Planting and training patterns for the tested fig trees. Trees were planted with tree spacing of 20.0, 6.6, 1.0, 0.6, and 0.2 m, in North-South rows with 2.5 m spacing between rows. This tree spacing accommodated plant densities (ρ) of 20, 61, 400, 667, and 2000 trees per 1000 m2. Each shoot was elongated from the mother shoot, which was arranged on the primary scaffold limbs with equal intervals of ca. 0.2 m. The shoots extended diagonally to the front and the opposite side, but for clarity are illustrated here as straight. The numbers of mother shoots were 100, 33, 5, 3, and 1 per tree, in ascending order of the plant densities.
Superfluous shoots were removed weekly from April to November each year. The numbers of these superfluous shoots were recorded as removed shoots (RS) and removed lateral shoots (RLS) for each tree and were converted to quantity per ground area (m2). All the shoots on each tree were used for measurements, but only 14–17 shoots were randomly selected from each tree at a plant density of 20 trees per 1000 m2. Every mature fruit from these shoots was harvested from the end of July to early December in ‘Masui Dauphine’ and from the mid-August to early December in ‘Houraishi’ in each year, and individual fresh fruit weight (fruit weight) was recorded using a digital scale. In February each year, the fresh weight of pruned shoots (shoot weight) was individually measured using a digital scale. In addition, the length and basal diameter (diameter between the second and third node from the base) of individual shoots were also measured for all dormant shoots of sixth year trees. To compare shoot size (weight) in this study with previous studies, the basal shoot width and shoot length taking the shoot weight into account were estimated as shown in Figure 2.

Relationship between shoot weight and shoot size (length and basal diameter) of ‘Masui Dauphine’ (filled circles) and ‘Houraishi’ (open circles) fig trees in the sixth year after planting.
The data for shoot weight and fruit production [number of fruit harvested (fruit number) per shoot (a), mean of fruit weight per shoot (b), and fruit yield per shoot (a by b)] were treated as primary units and aggregated for each age of each cultivar. First, the relationship between shoot weight and annual fruit production for each shoot and its significance were assessed by Pearson’s correlation coefficient analysis. Then, the monthly fruit productions of individual shoots (from the 1st to the 30th or the 31st in ‘Masui Dauphine’ and from the 15th to the 14th of the next month in ‘Houraishi’) were averaged for the following shoot weight classes: small, moderate, slightly large, large, and excessively large, with weight boundaries of 0.15, 0.4, 0.7, and 1.4 kg, respectively. The seasonal change in the monthly fruit production of individual shoots was represented using two-dimensional grids representing fruit production per month on the horizontal axis and the shoot weight classes on the vertical axis. Secondly, the shoot weight and annual fruit production, where fruit yield was converted from per shoot to per ground area (1000 m2), were averaged for each tree. These tree averages were compared among the plant densities using the Tukey–Kramer test. Thirdly, to understand the distribution of shoot sizes at different plant densities, the coefficient of variation (CV) of shoot size (length and basal diameter in the sixth year trees) within each density group was calculated. The above analysis was performed using Ekuseru-Toukei 2008 software (SSRI, Co., Ltd., Tokyo, Japan).
The tested trees grew steadily and were examined from the second year for plant densities of 2000, 667, and 400 trees per 1000 m2 and from the third and sixth years for densities of 61 and 20 trees per 1000 m2. The number of calculated trees (excluding abnormally small individuals) and their shoot growth are shown in Table 2. The shoot growth, i.e., the weight of pruned shoots and the number of superfluous shoots (RS and RLS), was greater in trees grown at higher densities. The shoot growth and its response to plant density were different between the two cultivars. In ‘Masui Dauphine’ trees, plant density continued to increase shoot weight and the number of superfluous shoots until the seventh year, consistent with results from a previous study that used this cultivar (Mano et al., 2011). ‘Houraishi’ trees tended to have larger shoots and fewer superfluous shoots than ‘Masui Dauphine’ trees. The increase in ‘Houraishi’ shoot weight with plant density of over 400 trees per 1000 m2 was suppressed after the third year.

Shoot growth of ‘Masui Dauphine’ and ‘Houraishi’ fig trees at various plant densities.
It is well known that plant growth is affected by plant density, and some studies showed that fig trees are no exception (e.g., Chithiraichelvan et al., 2017; Hosomi et al., 2013; Kumar et al., 2014; Mano et al., 2011). Many plant species obey the “Law of constant final yield” (Kira et al., 1953), which predicts that the total (or partial) biomass of an individual plant growing densely is suppressed with plant age and eventually the biomass yield (the weight per unit area) reaches a constant final value. In other words, the biomass yield remains greater in a higher density condition until the constant biomass is reached. The individual shoot weight and lateral growth in this study reflect the partial biomass of plant per unit area because all shoots were arranged in an equally occupied area of 0.5 m2. How these results reflect previous laws on plant density will be discussed in future reports; this study focused on the consistent enhancement of shoot growth by dense planting. It appears that ‘Masui Dauphine’ trees at all plant densities did not reach this constant stage until the seventh year. In contrast, densely planted ‘Houraishi’ trees reached the constant stage sooner and enhanced shoot growth according to density ceased in the third year after planting. It is known in other fruit trees that the effect of density appears earlier in vigorous trees (Kuroda et al., 1996), and consistent with these findings, the vigorous fig ‘Houraishi’ appeared to reach the constant stage sooner than the moderately vigorous fig ‘Masui Dauphine’.
Relationships between individual shoot growth and fruit productionThe data of all examined trees were pooled and the relationships between individual shoot weight and annual (or monthly) fruit production per shoot were analyzed. The results for ‘Masui Dauphine’ are shown in Figures 3 and 4. The annual fruit number was positively correlated with shoot weight in all years, and the correlation coefficients were relatively high except for the fourth and fifth years (Fig. 3). In terms of seasonal trends, smaller shoots had fewer mature fruit from August to September in the second year, and from September to November in subsequent years (Fig. 4). The tendency to produce more fruit on larger shoots was consistent with previous studies (e.g. Mano et al., 2011; Senda et al., 1996). The annual fruit weight was positively correlated with shoot weight in the second year, but no obvious correlation was observed in subsequent years (Fig. 3). The graphs of the seasonal trends show a complex relationship; in August for example, larger fruit were harvested from larger shoots in the second and sixth years, medium shoots in the third and seventh years, and small shoots in the fifth year (Fig. 4). These results suggest that the fruit weight of ‘Masui Dauphine’ is affected by a factor(s) other than shoot weight. The annual fruit yield per shoot and the seasonal trend were similar to that of the fruit number, reflecting that fruit yield per shoot was determined by the fruit number as no clear tendency existed for fruit weight.
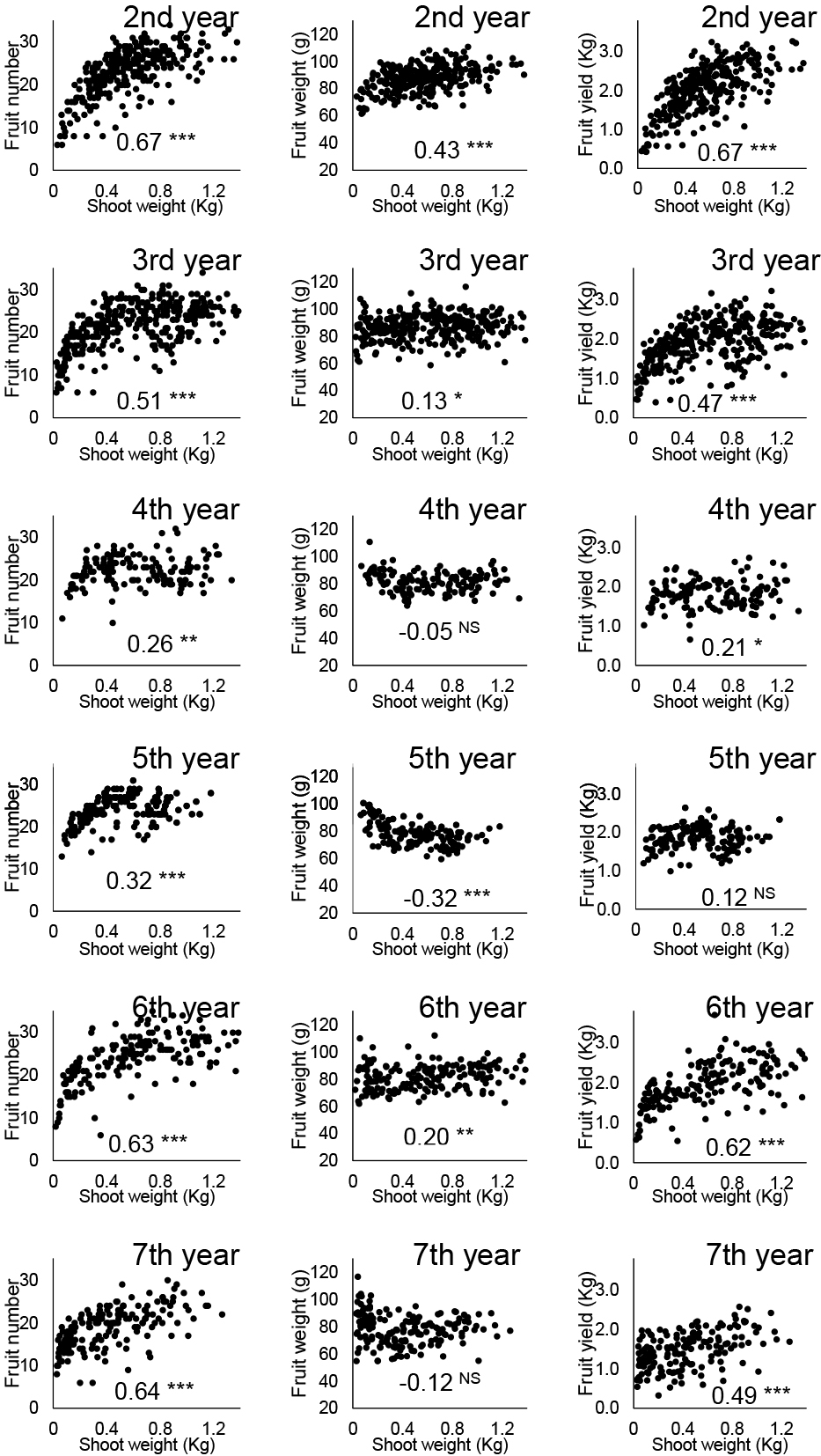
Yearly change of the relationship between individual shoot weight of ‘Masui Dauphine’ fig trees and annual fruit production per shoot, i.e., number of fruit harvested (left column), mean fruit weight (mid column), and fruit yield (right column) are shown with scatter plots. The regression analysis shows significance at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001.

Yearly change in the relationship between the individual shoot weight of ‘Masui Dauphine’ fig trees and monthly fruit production per shoot, i.e., number of fruit harvested (left column), mean fruit weight (mid column), and fruit yield (right column) are shown in a two-dimensional (Shoot weight × Harvest period) grid. The shoot weights are classified 1: Small (< 0.15 kg); 2: Moderate (0.15–0.4 kg); 3: Slightly large (0.4–0.7 kg); 4: Large (0.7–1.4 kg); 5: Excessively large (≥ 1.4 kg). Each grid shows the average with classified gradation pattern as each bottom note and “NC” indicating no calculation due to an insufficient number (n ≤ 2) of applicable shoots.
Results of the analyses for ‘Houraishi’ are shown in Figures 5 and 6. The annual fruit number was positively correlated with shoot weight, and the correlation coefficients were relatively high in all years (Fig. 5). Figure 6 shows that smaller shoots had fewer fruits after mid-September in the second and third years, and after mid-October in subsequent years. However, Figure 6 also shows that smaller shoots had more fruits from mid-August to mid-September in the first, sixth, and seventh years. The annual fruit weight was negatively correlated to shoot weight in all years except the second (Fig. 5). The seasonal trend graphs show that relatively large fruit were produced on smaller shoots, especially from mid-August to mid-September (Fig. 6). The annual fruit yield per shoot and the seasonal trend were related to the fruit number. These tendencies were similar to those of ‘Masui Dauphine’, but the correlation coefficients between shoot weight and annual fruit yield per shoot were relatively low in all years. The contribution to fruit yield by the higher number of fruit on larger shoots was likely negated by the lower fruit weight.

Yearly change in the relationship between individual shoot weight of ‘Houraishi’ fig trees and annual fruit production per shoot, i.e., number of fruit harvested (left column), mean fruit weight (mid column), and fruit yield (right column) are shown with scatter plots. The regression analysis shows significance at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001.

Yearly change in the relationship between the individual shoot weight of ‘Houraishi’ fig trees and monthly fruit production per shoot, i.e., number of fruit harvested (left column), mean fruit weight (mid column), and fruit yield (right column) are shown with a two-dimensional (Shoot weight × Harvest period) grid. The shoot weights are classified 1: Small (< 0.15 kg); 2: Moderate (0.15–0.4 kg); 3: Slightly large (0.4–0.7 kg); 4: Large (0.7–1.4 kg); 5: Excessively large (≥ 1.4 kg). Each grid shows the average with classified gradation pattern as each bottom note and “NC” indicating no calculation due to an insufficient number (n ≤ 2) of applicable shoots.
The fruit production results of the trees subjected to various plant densities are shown in Table 3. Fruit weight showed a tendency towards higher means at the lower plant densities for a few years in ‘Masui Dauphine’ and almost every year in ‘Houraishi’. As mentioned above, the tendency of smaller shoots to produce larger fruit was clear in ‘Houraishi’, but unclear in ‘Masui Dauphine’ (Figs. 3 and 5), and this was reflected in the trees with different plant densities. However, the effects of plant density on the fruit number per shoot and fruit yield per ground area were unclear. It can be seen from Figures 3 and 5 that correlations between shoot weight and annual fruit number, and fruit yield per shoot were low among the shoots larger than about 0.15 kg in ‘Masui Dauphine’ and 0.8 kg in ‘Houraishi’. It is plausible that no clear trend in fruit number or fruit yield per ground area between trees with different plant densities was detected (Table 3) because the shoot size exceeded this certain size except for the trees at the lowest density. In other words, the lowest density was specific. Although no significant difference was observed due to a lack of repetition, the values for fruit number and fruit yield per ground area for ‘Masui Dauphine’ were lower in the trees at the lowest density in the sixth year (Table 3).

Fruit production of ‘Masui Dauphine’ and ‘Houraishi’ fig trees at various plant densities.
These trend differences between the two cultivars provide important information for the application of planting or training methods for each cultivar. In ‘Masui Dauphine’, the positive correlation between shoot weight and fruit yield seen in this study suggests that weak shoots weighing 0.15 kg or less should be avoided to obtain a stable fruit yield. Dense planting (over 61 trees per 1000 m2 as an example in this study) with short pruning would be effective, because the shoot growth enhancement by dense planting is expected to last a long time (at least seven years) due to the slow appearance of the density effect (Table 2). However, it is known that excessive shoot vigor is not beneficial because it may have a negative effect on fruit skin color or increase the number of cracked fruit (Senda et al., 1996), but these characteristics were not measured in this study. An optimal shoot size (18–23 mm in basal diameter) for ‘Masui Dauphine’ has been recommended to prevent such harmful effects of excessive shoot growth (Senda et al., 1996). “Moderate” range (0.15–0.4 kg) shoots in this study were estimated to have a 17.8–23.3 mm basal diameter from the relational expression (3) as shown in Figure 2, and corresponded to such an optimal shoot size. The size of “Moderate” range shoots was similar to the average of the trees with a plant density of 61 trees per 1000 m2 (Table 2). Therefore, the conventional plant density of about 100 trees per 1000 m2 (Mano, 2015) is still a reasonable standard for ‘Masui Dauphine’ figs. The acceptable range of higher plant density could be wide for this cultivar when only the fruit weight and yield are regarded as the most important traits for cultivation. However, if the density causes adverse effects, such as color degradation and cracking of fruit, appropriate tree thinning will be required.
In ‘Houraishi’ trees, a shoot length from 0.2 to 0.4 m is regarded as a cultivation standard (Awamura et al., 1993). Even at the lowest plant density (20 trees per 1000 m2) in this study, the average shoot length was estimated to be 1.6–1.7 m from the relational expression (2) (Fig. 2), and was larger than Awamura’s standard. The current findings showed that small shoots reduced annual fruit number per shoot; however, more fruit setting on small shoots is still the main reason for adopting this cultivation standard (Awamura, 2015). The positive results of the cultivation standard correspond to more and larger fruits on smaller shoots form mid-August to mid-September in this study (Fig. 6). Moreover, the shoot number (about 5000) per unit area of open-center training for ‘Houraishi’ is about twice as high as the standard shoot number (about 2500) for ‘Masui Dauphine’. Even if fruit yields of individual shoots decline in late autumn, it will be economically advantageous for many very small shoots to produce higher fruit yields in early season of harvesting when market prices are higher (Kano et al., 2001). The only way to achieve such small shoots in the straight-line training with short pruning is to elongate the primary scaffold limb by planting at extremely low density. However, straight-line training cannot increase the number of shoots per unit area as much as open-center training. Also, the shoot growth of straight-line training is said to be uneven. In fact, the shoot growth in this study was markedly varied within a tree depending on the distance from the trunk on the primary scaffold limb. The CV of shoot size (length and basal diameter) was largest in the lowest density group in both cultivars (Table 4). Such characteristics of straight-line training showing that obtaining many and uniform short shoots is difficult even if a long primary scaffold limb is achieved, is unfavorable for fruit production of ‘Houraishi’ trees.

Effect of plant density on shoot size distribution of the 6th year fig trees of ‘Masui Dauphine’ and ‘Houraishi’.
Further improvements are needed to the methods for adapting straight-line training to ‘Houraishi’. For example, Nogata et al. (2010) attempted to adapt ‘Houraishi’ trees to form a double straight-line (H-type) with horizontal trellis training. This training may be advantageous to suppress shoot elongation by distributing the tree vigor twice across the primary scaffold limb and bending the shoots horizontally. In addition, “joint tree” training (Shibata et al., 2008), a unique training method developed in recent years, can achieve uniform shoot growth in the straight-line training of various fruit trees such as pear, persimmon, plum, and Japanese apricot, by grafting together the primary scaffold limbs of several trees. The application of joint tree training with straight-line training has been attempted with ‘Masui Dauphine’ fig trees (Kito et al., 2014), but not on ‘Houraishi’, although it could be an interesting approach to avoid irregular shoot growth along the primary scaffold limb in straight-line training.
The results of the fruit growth response to the shoot size at different plant densities were that larger shoots had a higher fruit yield in ‘Masui Dauphine’ trees and smaller shoots produced larger fruit in ‘Houraishi’ trees. It would be interesting to test whether such reactivity could be applied under various shoot growth conditions induced by factors other than plant density.
My special thanks to Dr. Takuo Yamakura for his expert professional advice on the tree planting. I am also grateful to Ms. Kazuko Uchimoto, Ms. Akemi Nanno, Ms. Akemi Matsukiyo, and Ms. Emiko Sakakura for their kind support on the surveys, and Mr. Yoshitaka Saikyo for his assistance in maintaining the trees.