ORIGINAL ARTICLES
Tomato Rootstock Breeding: Evaluation of Tomato Interspecific Hybrid Rootstocks Under Greenhouse Conditions
2020 Volume 89 Issue 5 Pages 575-585

Details
2020 Volume 89 Issue 5 Pages 575-585
Tomato grafting is an important agricultural technique to overcome biotic and abiotic stresses and increase plant growth and productivity. Breeding and selection of resistant and vigorous rootstocks are necessary to achieve grafting goals. Therefore, this study was conducted to evaluate eight tomato interspecific hybrids between accession Solanum lycopersicum L. LA1221 ‘VFNT cherry’, as the female parent and each of wild accessions S. chmielewskii LA1028 (Chm-1028); S. galapagense LA0530 (Gal-530); S. habrochaites CGN 15391 (Habr-15391) and LA1777 (Habr-1777); S. pennellii AusTRCF 312075 (Pen-312075) and LA716 (Pen-716); and S. pimpinellifolum AusTRCF311810 (Pim-311810) and CGN18388 (Pim-18388), as male parents, as rootstocks under greenhouse conditions compared to the commercial rootstock ‘Emperador’, to select the most suitable rootstock. S. lycopersicum ‘Santa Cruz Piedade’ was selected as a scion and used as self-grafted and non-grafted controls. Plant growth and productivity measurements were estimated for the grafted and non-grafted plants. The response to the evaluated rootstocks was not consistent for many of the studied traits, denoting that the scion was differently influenced by the rootstocks. Grafts did not always outperform self-grafts. Grafting onto either Pen-312075, Habr-15391, or Pim-18388. in addition to self-grafting, increased the total and marketable yields compared to the non-grafted plants, with proportions that would enable increased profits and cover the extra cost of grafting. In addition, fruits from these grafts had acceptable quality traits. Therefore, the interspecific hybrids Pen-312075, Habr-15391, and Pim-18388 are considered suitable rootstocks to improve the scion ‘Santa Cruz Piedade’. Greater and continuing efforts are needed to evaluate several interspecific hybrids, as well as different high yielding scion cultivars, to select the most suitable rootstocks for different scion cultivars.
Tomato (Solanum lycopersicum L.) is an important global vegetable crop that it is widely consumed due to its high nutritional value, antioxidant properties, good taste, and various preparation methods. Therefore, cultivated tomato areas are constantly increasing annually to meet demand (<http://faostat.fao.org/>). Tomato grafting is an important global cultivation technique for tomato production under a sustainable agriculture system, especially in greenhouses that can mitigate abiotic and biotic stresses and increase plant growth and productivity (Davis et al., 2008; Keatinge et al., 2014; King et al., 2010; Louws et al., 2010; Rouphael et al., 2010; Schwarz et al., 2010). Grafting is an alternative and quicker approach to improve cultivars compared to the relatively slow breeding technique. The main problems with the breeding of food crops for biotic and abiotic resistance can be overcome by using bred or selected rootstocks to confer resistance (Flores et al., 2010; King et al., 2010). Also, grafting techniques have enabled the rapid utilization of important economic characteristics using these species as rootstocks, particularly in wild species, without affecting their important horticultural traits.
Tomato hybrids (intraspecific hybrids) and interspecific hybrids are the most common tomato rootstocks (King et al., 2010). Interspecific hybrids are more vigorous and often make high quality rootstocks that greatly increase the rootstock’s genetic diversity (Djidonou et al., 2016). There are 12 close wild relatives of tomato that belong to the same Lycopersicum section (Scott et al., 2013). They are varied in their geographical distribution, habitat, reproduction systems and genetic polymorphisms (Bauchet and Causse, 2012). The wild species are of great use in breeding programs as sources of disease resistance and agronomic traits (Scott et al., 2013). The most common commercial rootstocks are S. lycopersicum × S. habrochaites F1 hybrids that have multiple resistance to soil-borne pathogens. However, the genetic potential of Solanum spp. has not yet been fully exploited for rootstock development. Therefore, this study was conducted to evaluate eight interspecific hybrids as rootstocks under greenhouse production conditions compared to the commercial rootstock ‘Emperador’, using self-grafting and non-grafting to select the most appropriate rootstock for commercial production.
This study was conducted in the greenhouses of the Agricultural Experiment Station (AES) of the Faculty of Agriculture, Cairo University, Giza, Egypt (30°01'07''N; 31°12'28''E) during the 2015 to 2017 winter plantings.
Plant materialsAccording to phenotype (Table 1), one accession of each of S. chmielewskii and S. galapagense and two accessions of each of S. habrochaites, S. pennellii, and S. pimpinellifolium were selected as male parents, along with accession S. lycopersicum LA1221 ‘VFNT cherry’, as the female parent, to produce interspecific hybrids. The crossing was carried out under plastic-house conditions at AES during the 2015 winter planting. S. lycopersicum CGN15403 ‘Santa Cruz Piedade’, an indeterminate and cluster-bearing cultivar, was selected as a scion and used as self- and non-grafted controls. As shown in Table 1, the AusTRCFs accessions were kindly provided by the Australian Tropical Crops and Forages Genetic Resources Center, Queensland, Australia; the CGNs accessions were kindly provided by the Centre for Genetic Resources the Netherlands, Wageningen University, the Netherlands; the LAs accessions were kindly provided by the University of California, Davis, USA; and the commercial rootstock ‘Emperador’ was purchased from Rijk Zwaan Company. The eight interspecific hybrids produced were evaluated as rootstocks compared to the commercial rootstock ‘Emperador’, in self-grafting and un-grafting treatments under plastic-house conditions during the 2016 and 2017 winter seasons, during which the weather is cold (Fig. 1).
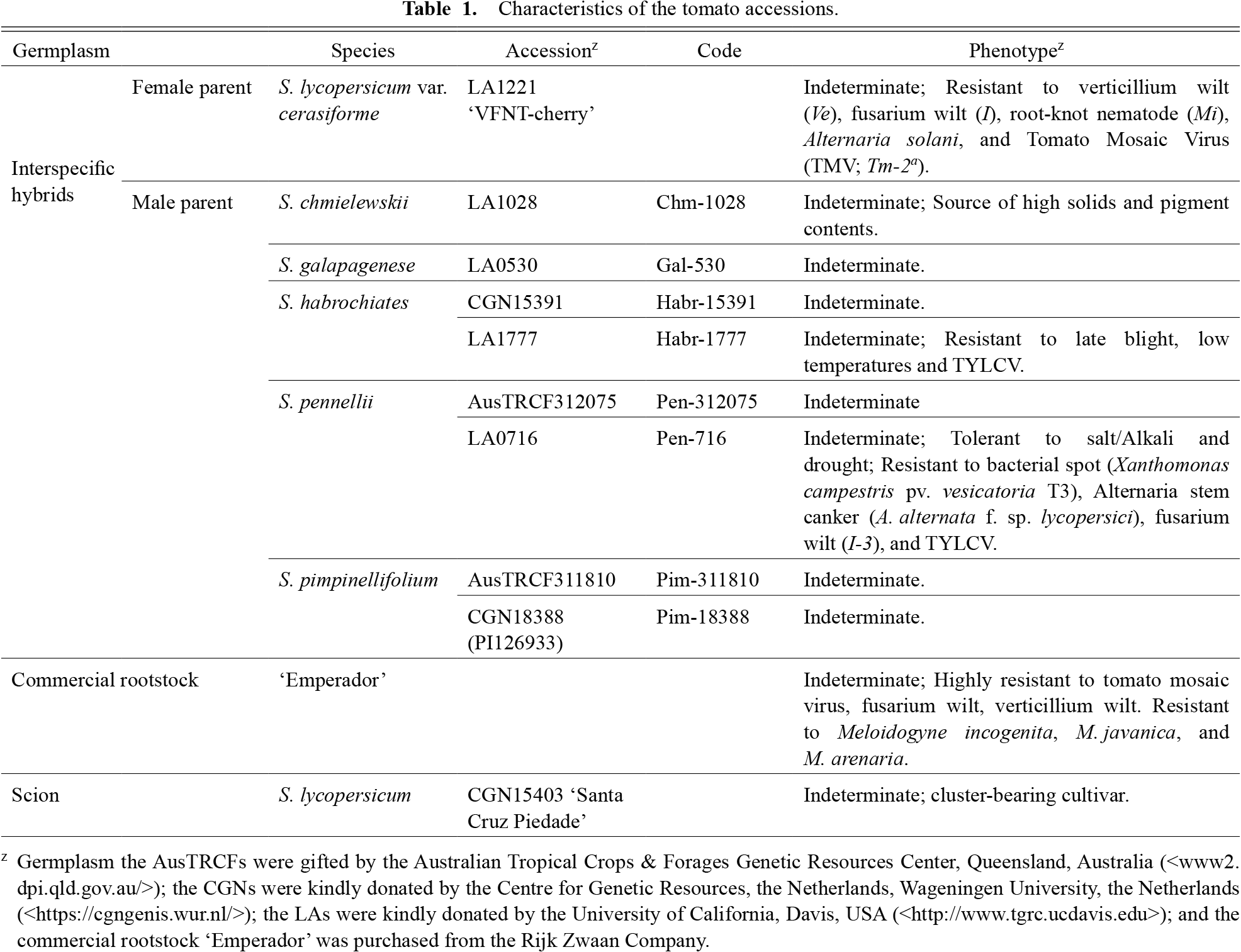
Characteristics of the tomato accessions.

Average monthly maximum and minimum temperatures, relative humidity, sunshine hours, and No. of sunny days during the period from October to May in the 2015 and 2016 seasons. Source: The Egyptian Meteorological Authority (EMA).
Seeds of the scion and rootstocks were sown on the 1st of August in seedling trays (209 and 84 cells, respectively) filled with a mixture of peat-moss and vermiculate (volume 1:1) enriched with macro and microelements. The trays were placed in an anti-virus greenhouse until full germination. Seedlings were fertilized daily using commercial compound fertilizer 20:20:20 (N:P:K). At the four to five true leaf stage, seedlings were cleft grafted as described by Oda (1995) and stems of the scion and rootstock were cut leaving 2–3 leaves remaining on the scion and rootstocks. The tapered stem end of the scion was placed into the cleft of the cut end of the rootstocks, followed by clipping. After grafting, both graft combinations were transferred immediately into a clear, closed, shady plastic tunnel 30 cm high with an inside relative humidity of almost 100%. Exposure to direct sunlight was prevented. From day 5, the tunnel was gradually ventilated daily and the light intensity was increased until the morning of day 7, when the plastic was removed. When wilting was observed, foliar spraying of grafted plants with water was effective at improving survival. Grafts were treated for another five days to ensure full recovery.
The grafted and non-grafted seedlings were transplanted in mid-September in the two seasons in a randomized complete block design (RCBD) with four replicates. The area of the greenhouse (9 × 60 m) was divided into five beds. Each bed was 1.2 m wide and plants were transplanted on both sides of the bed. The in-row distance between plants was 30 cm. Each experimental unit (EU) consisted of 10 plants. All cultural practices (fertilization, irrigation, weeding, and disease and insect control) were performed as recommended for commercial greenhouse tomato production.
Evaluated traitsPlant growth measurements, stem height (from the grafting area to end of the stem) and diameter (1 cm above the graft and the diameter of the root collar in the non-grafted plants), were measured twice 30 and 60 days after transplanting (DAT). The rate of stem elongation (RSE) was calculated as RSE = (H2 − H1)/(t2 − t1), where H2 is the plant height at day 60 after field transplanting, H1 is the plant height at day 30 after field transplanting, and (t2 − t1) the time elapsed during that period (30 days). The fifth round expanded leaf from the top was measured after 60 days from transplanting by the leaf weighting method as described by Pandey and Singh (2011). Also, the number of internodes at the beginning flowering and days from transplanting to produce each flower and fruit were performed in the five most representative plants per graft combination from EU.
Yield [early (the first three harvests), total (all the collected fruits), and marketable (all normal fruits); kg/m2), yield components (No. of flowers and fruits/cluster (from the second to sixth clusters), fruit set %, and average fruit weight), and fruit quality traits (firmness, pH of juice, TSS%, and vitamin C content) were estimated. Cracks, blossom-end rot, external watery translucent tissue, and small immature fruit (< 30 g) were classified as non-marketable yield. At the peak harvesting period, samples of 20 fully red-ripe fruits at the peak harvest from the EU were harvested and washed with distilled water for analysis of fruit traits. Fruit firmness was determined using a food pressure tester (Force Gauge Model M4-200) Mark-10 (Series 4). Then, the fruit extract was obtained by blending and filtering the flesh. TSS was determined using a hand refractometer. pH value was determined by immersing the glass electrode of a pH meter into the fruit extract. Vitamin C (ascorbic acid) was measured using 2,6 dichlorophenol indophenol dye (AOAC, 1990).
Statistical analysisData were statistically analyzed using MSTAT-C v. 2.1 (Michigan State University, Michigan, USA) and mean comparisons were based on Duncan’s multiple range test at a 0.05 probability level (Steel and Torrie, 1981).
Results of stem height, stem diameter, and leaf area showed significant effects of grafting on vegetative growth as illustrated in Figures 2 and 3. Grafting stimulated vegetative growth; self-grafts during the two evaluated seasons had the most significant values of these traits compared to nongrafted plants, which attained the lowest values. The evaluated tomato rootstocks showed a wide range of effects on vegetative growth of the scion with significant differences among them.
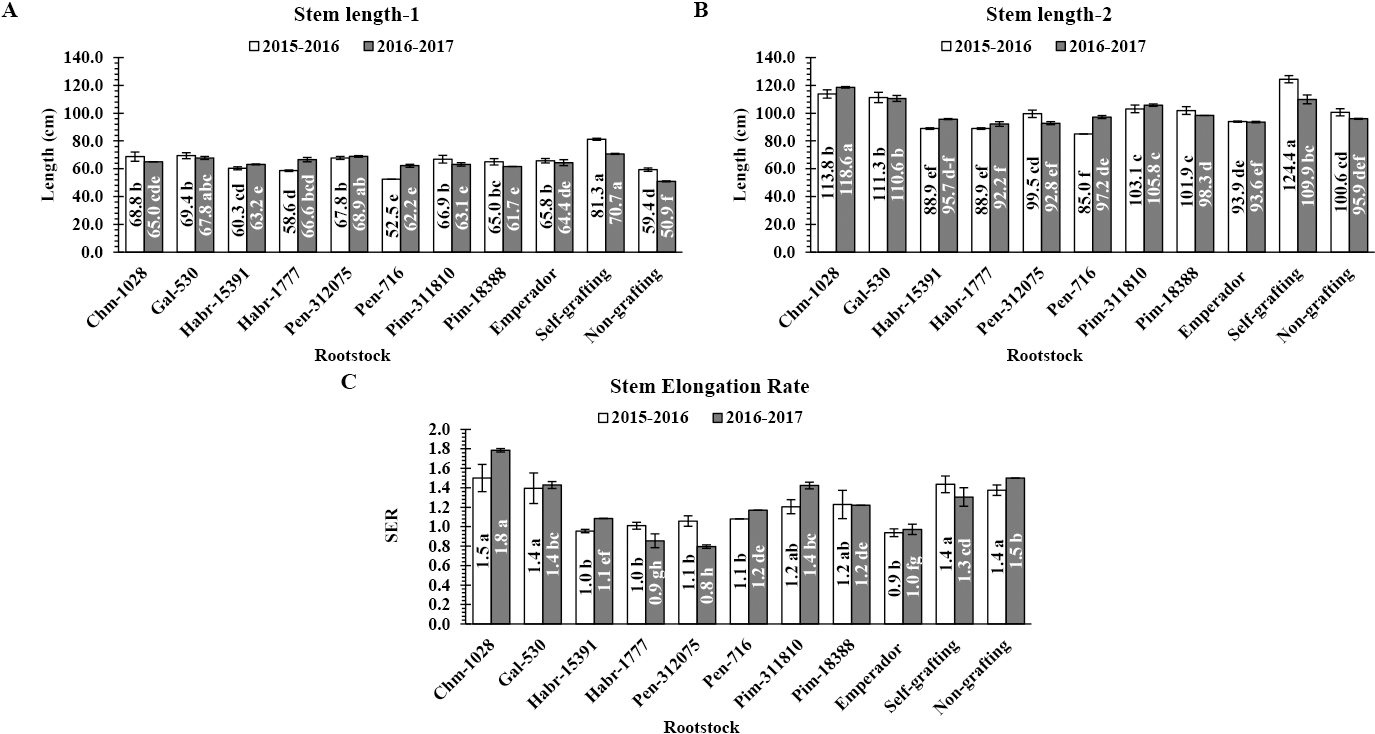
Stem length at 30 (SL-1; A) and 60 DAT (SL-2; B) and stem elongation rate (SER; C) of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated tomato interspecific hybrids, ‘Emperador’, self-grafts and the non-grafted plants during the 2015–2016 and 2016–2017 seasons. Interspecific hybrids were between Solanum lycopersicum LA1221 ‘VFNT cherry’, as a female parent and each of S. chmielewskii LA1028 (Chm-1028), S. galapagense LA0530 (Gal-530), S. habrochaites CGN15391 (Habr-15391), S. habrochaites LA1777 (Habr-1777), S. pennellii LA0716 (Pen-716), S. pennellii AusTRCF312075 (Pen-312075), S. pimpinellifolium AusTRCF311810 (Pim-311810), and S. pimpinellifolium CGN18388 (Pim-18388) as male parents. The AusTRCFs were kindly provided by the Australian Tropical Crops & Forages Genetic Resources Center, Queensland, Australia; The CGNs were kindly provided by the Centre for Genetic Resources the Netherlands, Wageningen University, the Netherlands; The LAs were kindly provided by the University of California, Davis, USA; and the commercial rootstock ‘Emperador’ was purchased from the Rijk Zwaan Company. Columns with the same letter represent values that are not significantly different at the 5% level of probability according to Duncan’s multiple range test. Vertical bars represent ± standard error (S.E.) of the mean. The S.E. was calculated across four replicates for each year.

Stem diameter at 30 (SD-1; A) and 60 DAT (SD-2; B) and the fifth leaf area (C) of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated tomato interspecific hybrids, ‘Emperador’, self-grafts and non-grafted plants during the 2015–2106 and 2016–2017 seasons. Interspecific hybrids were between Solanum lycopersicum LA1221 ‘VFNT cherry’, as a female parent and each of S. chmielewskii LA1028 (Chm-1028), S. galapagense LA0530 (Gal-530), S. habrochaites CGN15391 (Habr-15391), S. habrochaites LA1777 (Habr-1777), S. pennellii LA0716 (Pen-716), S. pennellii AusTRCF312075 (Pen-312075), S. pimpinellifolium AusTRCF311810 (Pim-311810), and S. pimpinellifolium CGN18388 (Pim-18388) as male parents. The AusTRCFs were kindly provided by the Australian Tropical Crops & Forages Genetic Resources Center, Queensland, Australia; The CGNs were kindly provided by the Centre for Genetic Resources the Netherlands, Wageningen University, the Netherlands; The LAs were kindly provided by the University of California, Davis, USA; and the commercial rootstock ‘Emperador’ was purchased from the Rijk Zwaan Company. Columns with the same letter represent values that were not significantly different at the 5% level of probability according to Duncan’s multiple range test. Vertical bars represent ± standard error (S.E.) of the mean. The S.E. was calculated across four replicates for each year.
For plant height at 30 DAT, grafts onto either Gal-530 or Pen-312075 as self-grafts had the tallest grafts over the two seasons without significant differences in the 2016–2017 season only (Fig. 2A). The other rootstocks had significantly longer scion stem lengths than non-grafting, except for Pen-716 in the 2015–2016 season. After 60 DAT, rootstocks of Chm-1028 and Gal-530 in addition to self-grafting had the most significant stem length values over the two seasons with significant differences between them, followed by Pim311810 and Pim-18388 rootstocks (Fig. 2B). The other rootstocks had lower values than non-grafted plants in terms of stem length, except for Pen-716 in the 2016–2017 season. The stem elongation rate (SER) varied over the two seasons (Fig. 2C). There were large significant differences between the evaluated rootstocks during the second evaluation season, whereas in the first season there were few significant differences between treatments. Generally, during these two periods of stem length estimation, grafts onto rootstocks Chm-1028, Gal-530, and Pim-311810 in addition to the non-grafted plants had the most significant SER (Fig. 2C) without significant differences between them in the 2105–2016 season only. Rootstocks Habr-15391, ‘Emperador’, Habr-1777, and Pen-312075 had the lowest SER, especially in the 2016–2017 season, with small significant differences between them.
The stem of grafts onto ‘Emperador’ in the 2015–2016 season and onto either Pen-716, Habr-1777, Pen-312075, or with self-grafting in the 2016–2017 season had the most significant thickness at 30 DAT (Fig. 3A). After 60 DAT and over the two seasons, grafts onto either Pen-312075, Habr-15391, Habr-1777, Pen-716, or ‘Emperador’ had the largest stem thickness after self-grafts, with significant differences between them (Fig. 3B). Grafts onto Gal-530 and Pim-311810 attained the lowest stem thickness along with the non-grafted plants during the estimation periods and the two seasons (Fig. 3A, B).
The largest significant leaf areas were observed with grafts onto Habr-1777 and self-grafts over the two seasons (Fig. 3C). Also, grafts onto either Gal-530 or Chm-1028 had the second largest leaf area and were statistically similar in the 2016–2017 season. The smallest leaves were observed with grafts onto Pim-311810 and un-grafted plants over the two seasons. Grafts onto the Pen-716 rootstock yielded conflicting results for leaf area over the two seasons, giving the smallest leaves during the first season and the largest in the second season (Fig. 3C).
Flowering and yield componentsPositive significant effects of grafting on accelerating flowering and fruit ripening were observed by comparing the data of No. of days from transplanting to the first flower emission or the first fruit emission for grafts and non-grafted plants, as illustrated in Figure 4. Grafts flowered earlier than the non-grafted plants, except grafts onto Pen-312075, Habr-1777, and Pen-716 in the 2015–2016 season and Pim-18388 in the 2016–2017 season (Fig. 4A). Grafts onto either Chm-1028, self-grafting, Gal-530, or Habr-15391 were first to flower in the two seasons. However, both the grafted and un-grafted plants flowered with a fairly similar number of internodes (Fig. 4B). While grafts onto Habr-1777 flowered at the seventh internode, the other grafts mostly flowered at the eighth internode, except self-grafts and grafts onto Chm-1028 and ‘Emperador’ that flowered at about the ninth internode in the second season (Fig. 4B). Consistent with the results of the two seasons as illustrated in Figure 4C, the fruits of grafts onto Habr-15391 were the first to ripen. Furthermore, the fruits of grafts onto Pen-312075, Habr-1777, Pen-716, and ‘Emperador’ ripened earlier than the fruits of the non-grafted plants (Fig. 4C).

Days from transplanting to the first flower emission (A) and the first fruit emission (C) and number of internodes until flowering (B) of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated interspecific hybrids, ‘Emperador’, self-grafts and non-grafted plants during the 2015–2016 and 2016–2107 seasons. Interspecific hybrids were between LA1221 ‘VFNT cherry’, as a female parent and S. chmielewskii LA1028 (Chm-1028); S. galapagense LA0530 (Gal-530); S. habrochaites CGN15391 (Habr-15391) and LA1777 (Habr-1777); S. pennellii LA0716 (Pen-716) and AusTRCF312075 (Pen-312075); and S. pimpinellifolium AusTRCF311810 (Pim-311810) and CGN18388 (Pim-18388) as male parents. The AusTRCFs were kindly provided by the Australian Tropical Crops & Forages Genetic Resources Center, Queensland, Australia; The CGNs were kindly provided by the Centre for Genetic Resources the Netherlands, Wageningen University, the Netherlands; The LAs were kindly provided by the University of California, Davis, USA; and the commercial rootstock ‘Emperador’ was purchased from the Rijk Zwaan Company. Columns with the same letter represent values that were not significantly different at the 5% level of probability according to Duncan’s multiple range test. Vertical bars represent ± standard error (S.E.) of the mean. The S.E. was calculated across four replicates for each year.
Grafting had a markedly positive impact on No. of flowers/inflorescence, No. of fruits/cluster, and fruit setting % for the scions in comparison to nongrafted plants as shown in Table 2. All grafts had a higher No. of flowers/inflorescences and No. of fruits/cluster than the non-grafted plants over the two seasons, except grafts onto Gal-530, which attained the lowest values. Consistently over the two seasons, grafts onto either Habr-1777, Pen-312075, or Habr-15391 had the most significant values of No. of flowers/inflorescence, No. of fruits/cluster, and fruit setting %. Self-grafts have similarly high statistical values to these rootstocks for No. flowers/inflorescence, No. fruits/cluster, and fruit setting ratio. Grafts on the commercial rootstock ‘Emperador’ did not differ significantly from those rootstocks for these traits during the first season and for No. of flowers/inflorescence in the second season.

Number of flowers and fruits/cluster, and fruit setting % of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated interspecific hybrids, ‘Emperador’, self-grafts and the non-grafted plants.
For yield traits, grafting had a clear effect on increasing plant yield as shown in Table 3. Self-grafts had higher early, total, and marketable yields than non-grafts. The effect of rootstocks on plant yields varied. Over the two seasons, rootstocks Habr-15391, Habr-1777, Pen-312075, and Pen-716 attained the highest significant EY along with self-grafts, with significant differences between them. For total yield (TY), Grafts onto Pen-312075 had the highest significant yield/plant over the two seasons followed by self-grafts and grafts onto Habr-15391. Over the two seasons, all interspecific hybrid rootstocks had higher yields than the commercial rootstock ‘Emperador’. Also, rootstock Pen-312075 attained the highest significant MY/plant over the two seasons. Rootstocks Habr-15391 and Habr-1777 in addition to self-grafting had the second most MY over the two seasons. Grafts onto Gal-530 had the lowest MY over the two seasons.

Yield components of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated interspecific hybrids, ‘Emperador’, self-grafts and the non-grafted plants.
The average fruit weight (AFW) was greatly affected by grafting. The AFW of self-grafting plants was higher than that of non-grafting plants. The highest AFW coincided with grafts onto either Pen-312075 or Pen-716. Rootstock Habr-1777 had the next highest AFW of the scion followed by rootstock Habr-15391 and self-grafting. The fruits of grafts on Chm-1028 and Gal-530 had significantly lower weight.
Fruit qualityFor fruit quality traits, firmness, pH of juice and TSS and vitamin C contents were estimated. Statistically, differences were observed between interspecific hybrids and each commercial rootstock, self-grafts, and non-grafted plants as illustrated in Figure 5. Generally, fruits of self-grafts were statistically similar to those of non-grafted plants in terms of fruit quality traits, especially in the first season. Over the two seasons, fruits of grafts onto Gal-530 had significantly higher firmness (Fig. 5A). In contrast, Chm-1028 rootstock produced softer fruits. During the 2016–2017 season, fruits of most interspecific hybrid rootstocks were significantly stiffer than those of non-grafted plants (Fig. 5A).

Tomato fruit quality traits [firmness (A), contents of TSS (B) and vitamin C (C), and pH of fruit juice (D)] of the scion ‘Santa Cruz Piedade’ grafted onto the evaluated interspecific hybrids, ‘Emperador’, self-grafts and non-grafted plants during the 2015–2016 and 2016–2017 seasons. Interspecific hybrids were between S. lycopersicum LA1221 ‘VFNT cherry’, as a female parent and S. chmielewskii LA1028 (Chm-1028), S. galapagense LA0530 (Gal-530), S. habrochaites CGN15391 (Habr-15391), S. habrochaites LA1777 (Habr-1777), S. pennellii LA0716 (Pen-716), S. pennellii AusTRCF312075 (Pen-312075), S. pimpinellifolium AusTRCF311810 (Pim-311810), and S. pimpinellifolium CGN18388 (Pim-18388) as male parents. The AusTRCFs were kindly provided by the Australian Tropical Crops & Forages Genetic Resources Center, Queensland, Australia; The CGNs were kindly provided by the Centre for Genetic Resources the Netherlands, Wageningen University, the Netherlands; The LAs were kindly provided by the University of California, Davis, USA; and the commercial rootstock ‘Emperador’ was purchased from the Rijk Zwaan Company. Columns with the same letter represent values that were not significantly different at the 5% level of probability according to Duncan’s multiple range test. Vertical bars represent ± standard error (S.E.) of the mean. The S.E. was calculated across four replicates for each year.
Fruits of self-grafts and the non-grafted plants had the lowest significant TSS, while the fruits of grafts on ‘Emperador’ had the highest significant TSS over the two seasons (Fig. 5B). The evaluated interspecific hybrid rootstocks, except Pen-312075 during the 2016–2017 season, produced fruits with higher TSS than the fruits of the non-grafted plants, as illustrated in Figure 5B.
The fruits from grafts onto Pen-716 were statistically higher in vitamin C content followed by grafts onto Habr-1777 (Fig. 5C) over the two seasons. In the 2015–2016 season, fruits from all grafts had significantly higher vitamin C content than those from non-grafted plants, while in the 2016–2017 season, fruits from non-grafts had significantly higher vitamin C content.
In terms of fruit juice pH over the two seasons, fruits of grafts onto either Pen-312075 or Habr-1777, in addition to those of self-grafts and non-grafted plants, had higher significant acidity (Fig. 5D), while those of grafts onto Gal-530 and Pim-311810 had lower significant acidity.
Tomato grafting began commercially in the early 1960s (Lee and Oda, 2003), and it is now an important cultivation technique for tomato production in many parts of the world under sustainable agriculture systems (King et al., 2010; Rivard et al., 2010) to overcome biotic and abiotic stresses (Keatinge et al., 2014; Louws et al., 2010; Schwarz et al., 2010) and to increase plant growth and productivity (Davis et al., 2008; Rouphael et al., 2010). This is accomplished by grafting onto resistant, vigorous, and strong-rooted rootstocks, which leads to stress resistance, increased absorption and translocation of water and nutrients (Djidonou et al., 2017), as well as augmented endogenous hormone production (Aloni et al., 2010; Zijlstra et al., 1994). Generally, grafting success depends on many factors, but genotypic factors are most important for the compatibility or incompatibility of the scion and rootstock, in addition to rootstocks traits. Therefore, breeding and selection of resistant and vigorous rootstocks is necessary to achieve grafting goals (King et al., 2010). Taxonomically closer scions and rootstocks have higher graft compatibility (Wang, 2011). There are 12 related wild tomato species, but only S. habrochaites has been bused as a commercial tomato rootstock in interspecific hybrid form (King et al., 2010; Rivard et al., 2010) such as the rootstocks ‘Beaufort’, ‘Maxifort’, ‘Body’, ‘Heman’, ‘42851’, and ‘43965’. Hence, the genetic potential of Solanum spp. has not yet been fully exploited for rootstock development. Interspecific hybrids were more vigorous and often produced higher quality rootstocks than intraspecific hybrids (Djidonou et al., 2013; Leonardi and Giuffrida, 2006; Schwarz et al., 2010, 2013). Additionally, wild tomato species cannot be used as rootstocks due to the different growth rates between them and cultivated tomato cultivars, especially at the seedling stage. Also, grafting onto the wild species S. habrochaites and S. pennellii reduced productivity (Bloom et al., 2004; Mahmoud, 2014; Ntatsi et al., 2014; Zeist et al., 2017). Therefore, this study was conducted to evaluate eight interspecific hybrids with the related wild species S. chmielewskii, S. galapagense, S. habrochaites, S. pennellii, and S. piminellifolium as rootstocks under greenhouse production conditions as compared to the commercial rootstock ‘Emperador’ in order to select the most suitable rootstocks, as well as to elucidate the grafting effects of self-grafts and non-grafted plants.
Compatibilities (i.e. complete survival of grafted plants, graft vigor, speedy flowering and fruit ripening, and high yield) were observed in the scion ‘Santa Cruz Piedade’ when grafted onto the interspecific hybrids and commercial rootstock in the present study. Grafting on a compatible rootstock achieved the following results: graft vigor (Figs. 2 and 3) resulted from increases in vascular bundle formation, hydraulic conduction, absorption and translocation of water and nutrients between the rootstock and the scion (Davis et al., 2008; Djidonou et al., 2017; Goto et al., 2013; Lee and Oda, 2003; Lee et al., 2010; Leonardi and Giuffrida, 2006; Martínez-Ballesta et al., 2010), an increased photosynthesis rate because of the scion canopy, and biomass accumulation, especially under low sunlight and CO2 concentrations under greenhouse conditions during winter months (Hu et al., 2006); faster flowering and fruit ripening of grafts (Fig. 4) by rapid vascular fusion between the scion and rootstock of grafts that were not subjected to stresses during the grafting and adaptation processes; and increased productivity (Tables 2 and 3) because of improved plant vigor, metabolism rate, No. of flowers/cluster, fruit setting %, No. fruits/cluster, and average fruit weight (Al-Harbi et al., 2016; Djidonou et al., 2017; Khah et al., 2006; Lee et al., 2010; Martínez-Ballesta et al., 2010; Turhan et al., 2011). All of these factors were controlled by a hormonal signal from the rootstock to the scion (Aloni et al., 2010). With regard to grafting effects on fruit quality traits, findings demonstrated that an increase, decrease or no change in quality parameters of fruits from grafted plants can occur (Fig. 5). These results agree with Davis et al. (2008), Flores et al. (2010), and Rouphael et al. (2010), who reported on conflicting reports regarding changes in fruit quality parameters following grafting. The inconsistencies could be caused by the different production environments (e.g. light intensity, air temperature), methods (e.g. soilless vs. soil culture, irrigation, and fertilization), rootstock-scion combinations used, and harvest date (Davis et al., 2008; Rouphael et al., 2010). Changes in the scion are controlled by the rootstock through controlled uptake and translocation of water, minerals, and plant hormones (Lee and Oda, 2003). However, it is important that the quality characteristics and composition of the final product of grafted plants remain unchanged or be improved with respect to non-grafted plants.
Contrary to previous reports (Djidonou et al., 2013, 2016, 2017; Flores et al., 2010; Khah et al., 2006; Oztekin et al., 2009), the self-grafts were taller, thicker, had larger leaves (Figs. 2 and 3), and flowering and fruit ripening were earlier (Fig. 4), with a higher yield (Table 3) than self-rooted plants. This is because self-grafting leads to an increase in internal hormone production (Aloni et al., 2010; Dong et al., 2008). Also, the scion ‘Santa Cruz Piedade’ is low-temperature tolerant and has a strong root system (Oztekin et al., 2009). Grieneisen et al. (2018). A meta-analysis of the published results from tomato grafting studies found no differences between self-grafts and non-grafted plants, and self-grafted plants differed from non-grafted ones in only one of the 53 cases with statistical data. Therefore, they suggested that including self-grafting treatment in experiments is probably not necessary.
Most of the observed variability in the estimated morphological and fruit production traits in this study was a response to the different rootstocks used. However, the response to the evaluated rootstocks was not consistent for many of the estimated traits (Figs. 2, 3, 4, and 5; Tables 2 and 3), indicating that the scion was influenced in different ways by the rootstocks. Also, the scion response to the evaluated rootstocks differed over the two seasons due to the different climatic conditions as shown in Figure 1. Grafts did not always outperform self-grafts, and this emphasizes the importance of selecting the correct rootstock to improve a specific scion.
Generally, grafts were more vigorous and productive than self-rooted plants. This is supported by previous findings (Djidonou et al., 2013, 2016, 2017; Khah et al., 2006; Turhan et al., 2011). Consequently, grafting is a useful tool for commercial growers. Over the two seasons, grafts on interspecific hybrids were taller (Chm-1028, Gal-530, Pim-311810, and Pim18388), thicker (all grafts except on Gal-530), had larger leaves (most of grafts) and higher yields (especially grafts onto Pen-312075 and Habr-15391) than self-rooted plants. Also, the fruits of grafts onto interspecific hybrid rootstocks were higher quality in terms of firmness (Gal-530), TSS (all), pH (Pen-312075 and Habr-1777), and vitamin C content (especially Pen-716), than self-rooted plants. Contrary to these results, Riga (2015) found in a similar study conducted in Spain that grafting has negative effects on plant growth when using rootstocks sensitive to low-temperature, as there are no commercially available tolerant rootstocks.
Grafts on interspecific hybrids were significantly taller (Chm-1028, Gal-530, Pim-311810, and Pim-18388) and had significantly larger leaves (Habr-1777, Gal-530, and Chm-1028), stem diameters (Pen-312075 and Habr-15391, especially at 60 DAT), and yields (all interspecific hybrids except Gal-530) than grafts on ‘Emperador’. In addition, the fruits of grafts on interspecific hybrid rootstocks were higher quality in terms of firmness (Pen-312075 and Pen-716), and pH (Pen-31207, Pen-716, and Habr-1777) than grafts on ‘Emperador’.
The costs for growing grafted plants are higher compared to non-grafted plants due to the additional costs of rootstock seeds, labor costs incurred for sowing and managing rootstock seeds and seedlings. There are also labor costs for grafting, healing and acclimatization of grafts, as well as for de-suckering of shoots that develop below the graft interface and removal of adventitious roots; therefore, the productivity of grafts should be increased enough to make a viable return to cover these high costs. This was achieved with grafts onto either Pen-312075, Habr-15391, or Pim-18388, as well as self-grafts. Moreover, the fruits all had acceptable quality traits.
ConclusionThe interspecific hybrids Pen-312075, Habr-15391, and Pim-18388 in addition to self-grafting are considered suitable rootstocks to improve the scion ‘Santa Cruz Piedade’. According to the availability of genetic diversity among tomato germplasm (Bauchet and Causse, 2012), and heterosis of many growth traits, especially abiotic and biotic stress resistance (Birchler et al., 2010), the greater and continued efforts are needed to evaluate several interspecific hybrids as well as different highly yielding scion cultivars to select the highly suitable rootstocks with different scion cultivars.