Abstract
The growth and yield of tomatoes in greenhouses covered by heat insulation film were investigated in Ishigaki, a subtropical area of Japan with high temperature and high solar radiation. High-density planting of tomatoes was carried out in June, July, August, October, and November, with a low node-order pinching system. The plants were grown in two greenhouses, one covered by a thermal barrier film (TBF) for heat insulation and the other covered by a polyolefin film (PO). Under TBF, the maximum temperature was up to 3.6°C lower than that under PO, and the mean temperature was 0.4°C lower. The leaf area index (LAI) of the June planting was 3.0 under TBF and 3.4 under PO, significantly lower than the LAIs of the other plantings, which ranged from 3.9 to 5.4. The specific leaf area (SLA) varied with the cultivation season, with that under TBF and PO being highest in November plantings grown in winter. SLA under TBF was higher than that under PO in the same cultivation seasons. The amount of total aboveground dry matter was significantly lower in the June planting under TBF and showed no difference in other plantings. Distribution to fruit was higher under TBF than under PO, especially in summer cultivation. The yields under TBF were significantly higher than those under PO in the July planting, and the yields under PO were significantly higher in the November planting. Under TBF, the rate of fruit cracking was reduced in the June, July, and August plantings. Under TBF and PO, plantings in October and November produced puffy fruits. The light use efficiency under TBF was higher than that under PO in the same cultivation seasons, apart from the June planting. Tomato cultivation under TBF reduced the damage from intense solar radiation during summer. TBF enabled penetration of sufficient solar radiation to distribute photosynthates to each part of the plant because of a limited sink and abundant solar radiation in a subtropical summer. These results indicate that TBF and sink-limited cultivation are effective systems for use in the high temperature and high solar radiation environments of tropical and subtropical regions.
Introduction
Tomato (Solanum lycopersicum L.) is one of the most important horticultural crops in Japan, where year-round production is necessary to meet the annual demand for tomatoes. Moreover, the demand for tomatoes is also increasing worldwide. Tomato production in Southeast Asia and South Asia increased from 1 million and 15 million tons, respectively, in 2008 to 15 million and 26 million tons in 2018 (FAOSTAT, 2018). Tomatoes are becoming a key fruit even in tropical and subtropical areas.
Tomato plants flourish at a daily mean temperature of 15–25°C (Kawasaki and Yoneda, 2019), but tomato production is adversely affected by high temperatures. The main causes of reduced yield of good-quality tomato fruit under higher temperatures are poor fruit set, reduced plant vigor, and development of physiological disorders, such as fruit cracking, blossom-end rot, and yellow shoulder fruit (Suzuki, 2019). In recent years, the interiors of greenhouses in Japan have become dangerously hot during summer, and it is always more difficult to cool the interiors of greenhouses than it is to heat them (Suzuki, 2019). Establishing tomato cultivation techniques under high temperatures and intense sunlight is an important issue for both horticulture in Japan and for tropical and subtropical agriculture in general.
Various studies have explored methods to counteract high temperatures in greenhouse cultivation; such methods include ventilation (Ide et al., 2007), shading (Wada et al., 2006, 2013), heat pumps (Kawasaki and Ahn, 2015; Kawashima et al., 2011), and fog cooling (Hiei et al., 2015). Covering materials are also useful for reducing solar radiation inside a greenhouse. A specific type of film that blocks ultraviolet (UV) and/or infrared rays has been developed and researched. Luthria et al. (2006) studied the effect of UV radiation on the phenolic content of tomato fruits by comparing the results of using either a covering film that blocked UV radiation or not using such a film. One of the heat insulation films that has been developed is an infrared ray blocking (IRB) film that transmits 40–60% of solar radiation above 700 nm. Fukuoka et al. (2009) reported that a higher watermelon sugar content can be achieved by reducing the temperature with an IRB film covering during the latter half of the fruit developmental period. These kinds of films are a technology that could be applied not only to ordinary greenhouses, but also to large scale modern greenhouses. However, only a few studies using such films have been done.
Low node-order pinching and high-density planting is a method that repeats short-term tomato cultivation with 1–3 trusses. According to Suzuki et al. (2011), the advantages of low node-order pinching and high-density planting are: 1) a short cultivation period and repeatable planting, 2) ease of data collection regarding production, 3) reduced risk of failure, 4) ability to choose suitable cultivars for the season, 5) no need to work at height, and 6) ease of producing tomatoes with a high sugar content. The method of repeating short-term cultivation appears to be effective for use in hot environments, such as subtropical regions.
In this study, we investigated the effects of a heat insulation film on tomato growth in greenhouses in subtropical regions under a low node-order pinching and high-density planting system. Plants were grown in two greenhouses, one of which was covered with a heat insulation film and the other of which was covered with a polyolefin film. We also investigated the effects of seasonal differences on tomato growth in subtropical regions using five cultivations a year to reveal the effects of differences in temperature and photosynthetic active radiation (PAR) on tomato growth and yield in a high-temperature environment.
Materials and Methods
The experiment was conducted in the greenhouses of the Tropical Agriculture Research Front of the Japan International Research Center for Agricultural Sciences (lat. 24.34 N, long. 124.16 E) on Ishigaki island, Okinawa. The location’s climate is classified as subtropical, with high solar radiation in summer and high temperatures and humidity over the whole year.
In 2019, we performed five tomato plantings (‘Momotaro hope’; Takii & Co., Ltd., Kyoto, Japan) in two greenhouses with the ridges oriented North–South. One greenhouse was covered with a thermal barrier film (TBF) (MF-450; Fujifilm Corporation, Tokyo, Japan), and the other was covered with an agricultural polyolefin film (PO) (Diaster, Thickness 0.15 mm; Mitsubishi Chemical Agri Dream Co., Ltd., Tokyo, Japan) as a control. Compared to PO, TBF reduces near-infrared radiation, the wavelength of which ranges from 700 to 1050 nm, by 59%; photosynthetic active radiation (PAR), ranging from 400 to 700 nm, by 22%; and UV radiation, ranging from 250 to 400 nm, by 66% based on the results of actual measurements. The greenhouses were each 6 m wide and 25 m long, with an eaves height of 2.0 m and a ridge height of 3.3 m. A single layer of shade curtain, with a ratio of transmitted light intensity of 65%, was installed under the center of the ceiling of each greenhouse. We also installed a fog cooling system (Coolnet Pro; Netafim Japan Co., Ltd., Tokyo, Japan for TBF, and a similar product under development for PO), a heat pump (GI-P112-A1 and KBHP-GP140-1; Kubota Corporation, Osaka, Japan), a ventilation fan (NK-74DGA; Panasonic Corporation, Osaka, Japan) and circulation fans (CV-3510; Nakatomi Corporation, Nagano, Japan) in each greenhouse, in addition to a roll-up ventilation window (Kankit; Toto Kogyo Co., Ltd., Tokyo, Japan) on the side of each greenhouse. We kept the ventilation windows open and the ventilation fan running when the temperature at a height of 1.5 m in the center of the greenhouses was above 29°C in the morning and above 20°C in the afternoon, we kept the fog cooling system operating at 30°C and above, and we ensured that the shade curtain was closed at 31°C and above. We also operated the heat pump when the inside temperature was above 15°C after sunset and closed the ventilation windows and the shade curtain to cool the air temperature inside the greenhouses during the night. These devices were controlled by an integrated environmental control system (Smart Saien’s Cloud; Panasonic). The integrated environmental control system also recorded air temperature and relative humidity at the center of the greenhouses every minute.
There were three rows of cultivation beds in each greenhouse, with 20 beds in each row. The cultivation beds (1.0 m long × 0.3 m wide × 0.15 m deep) were filled with a commercial medium consisting mainly of cocopeat (BVB No.7-1P; Toyotane Co., Ltd., Toyohashi, Japan). We transplanted seedlings with seven true leaves on June 26 in the east row of the greenhouses, those with six true leaves on July 25 in the center row, those with nine true leaves on August 21 in the west row, those with eight true leaves on October 7 in the east row, and those with six true leaves on November 11 in the center row. The low node-order pinching and high-density planting system was used for all cultivations, and the plant density was 4.25 plant·m−2. We carried out prompt periodic operations for training, fruit thinning, and lateral bud pruning. Plants were trained vertically with a single stem and topped at the second upper leaf above the fourth truss. Three times per week, we sprayed the flowers with a solution containing 4-chlorophenoxy acetate (Tomato Tone; ISK Biosciences K.K., Tokyo, Japan). We adjusted each truss to bear four fruit.
We supplied the plants with a commercial nutrient solution (Otsuka House Solution SA; OAT Agrio Co., Ltd., Tokyo, Japan), which was adjusted to an electrical conductivity of 0.8–1.6 dS·m−1 as the plants grew. We based the amount of irrigation on the rate of drainage, which was maintained at 20% of the supply, and the drainage water was not reused for irrigation.
We measured stem length and number of leaves for six plants in each greenhouse each week until the main stem was topped. We also measured the leaf area, using a leaf area meter (CI-203CA; CID Bio-Science, Inc., WA, USA), and fresh weight (FW) and dry weight (DW) of leaves, stems, and fruits by destructive sampling at transplanting, pinching, and the end of the cultivation. We determined DW by oven-drying at 105°C to a constant weight. We calculated the leaf area index (LAI) and the specific leaf area (SLA) according to the measured leaf area. We harvested mature fruits three times a week and measured the number and FW of the fruits for six plants in each greenhouse. We also counted marketable fruits and fruits with physiological disorders. We measured the weight and soluble solids content (SSC) of fruits to select marketable fruit using a fruit selector (K-SS300-LC; Kubota), which was calibrated for each cultivation. We calculated fruit DW by multiplying the FW by the average dry matter content of fruit that we measured during harvest for each cultivation.
PAR was assumed to be 43% of the global radiation for PO and 58% of the global radiation for TBF based on actual measurements. Light use efficiency (LUE) was calculated based on the method described by Itoh et al. (2020) and Higashide et al. (2012). We estimated the cumulative intercepted PAR by calculating daily LAI based on destructive analysis at transplanting, pinching, and the end of the cultivation and daily PAR based on the climate data recorded by the Ishigakijima local meteorological observatory (lat. 24.20 N, long. 124.09 E) under the Japan Meteorological Agency, 5.6 km southwest from the greenhouses. We then calculated LUE from the total aboveground dry matter (TDM) at transplanting, pinching, and the end of cultivation and the cumulative intercepted PAR at those times.
All statistical analyses were performed using R software (R Foundation for Statistical Computing, Vienna, Austria). Mean separations were conducted using a Tukey’s HSD test protected by Analysis of Variance at P < 0.05 and the Student’s t-test for the yield for each cultivation.
Results
Table 1 shows a summary of each cultivation. During all five cultivations, the daily mean temperature at the center of the TBF greenhouse was 0.1–0.5°C lower than that of the PO greenhouse. In the June planting, tomatoes under both PO and TBF were grown in a hot environment, with a mean daily temperature above 25°C. The mean daytime temperature from June to August was above 30°C on most days in both greenhouses. The cumulative PAR of each cultivation ranged from 373 to 558 MJ·m−2 for TBF and 478 to 716 MJ·m−2 for PO. The cultivation in both greenhouses had a minimum cumulative PAR in the October planting and a maximum cumulative PAR in the June planting. The shortest cultivation period was 93 days for the June planting, and the longest was 122 days for the November planting. In the June and July plantings, TBF harvesting finished two and four days earlier than PO, respectively. There was no difference in the other plantings, and all cultivations in both greenhouses were finished at the same time. Figure 1 shows the daily mean temperature, the maximum and minimum temperatures, and the daily PAR in the greenhouses during each cultivation. In the June, July, and August plantings, daily maximum temperatures exceeded 30°C for most of the growing season. The daily maximum temperature under TBF was up to 3.6°C lower than that under PO on days with clear skies during the day. In the June and July plantings, the daily minimum temperature exceeded 20°C, whereas it remained stable at 15°C in the October and November plantings, partly because of night cooling. During the growing period, the average relative humidity for each cultivation was 80.1% at the lowest and 84.3% at the highest, with no difference regarding the time of cultivation or the greenhouse (data omitted).


The growth of tomatoes varied depending on the cultivation season and the greenhouses. Compared to the other cultivations, the rate of stem elongation for the June planting was slower, at 2.86 cm·day−1 for TBF and 2.81 cm·day−1 for PO, whereas the rate for the November planting was faster, at 4.36 cm·day−1 for TBF and 4.21 cm·day−1 for PO (Fig. 2). We observed no significant difference between TBF and PO regarding the stem elongation rates or stem length in each cultivation season. Stem length was longest in the November planting. The number of leaves also varied with the cultivation season. At the time of pinching, the number of leaves was higher in the July planting, at 23.8 for TBF and 25.2 for PO (Table 2). Conversely, the number of leaves was lower in the June planting, at 18.8 for TBF and 19.3 for PO. In the other cultivations, the range was 19–22 leaves. In the experiment, the rate of leaf appearance was poorly correlated with temperature and solar radiation (data omitted). In all cultivations, the LAI ranged from 3 to 5. We found no significant difference between TBF and PO in terms of LAI in each cultivation season. The SLA of tomatoes planted in June was significantly smaller under both TBF and PO, and it was significantly larger for the October and November plantings. Moreover, the SLA tended to be larger under TBF than under PO during the same cultivation season. The leaf appearance rate ranged from 0.25 leaves·day−1 to 0.45 leaves·day−1, and was faster in the July planting. The June planting under TBF was the lightest in terms of TDM at the end of cultivation, and the TDM of the November planting under PO was the heaviest (Table 3). The TDM for the other cultivations ranged from 230 to 274 g/plant, with no significant differences. Distribution to fruit tended to be higher under TBF than under PO, exceeding 49% in all cultivations, and the difference was particularly large in summer cultivation. Under PO, the November planting showed lower distribution to a side shoot, at 3.7%, and higher distribution to fruit than the other cultivations. Tomatoes planted in June–October showed high distribution to a side shoot. Under both TBF and PO, there was higher distribution to leaves and stems in June, July, and August plantings.



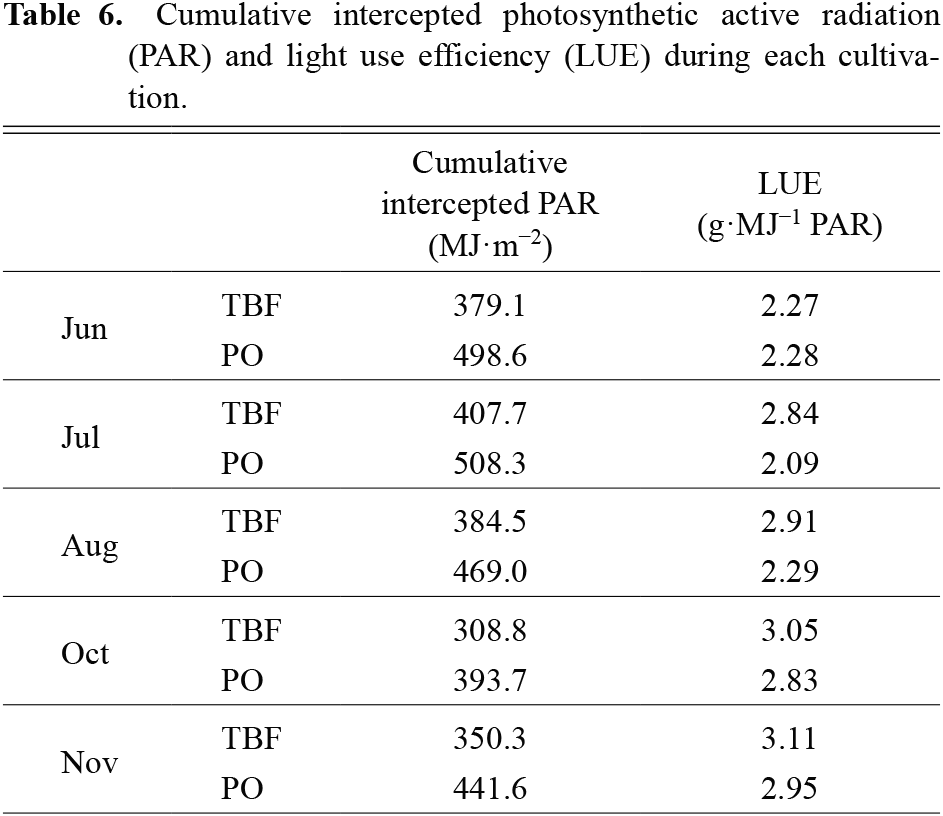
Figure 3 shows the yield per plant in all cultivations. The lowest yield was 1.5 kg/plant under TBF in the June planting, and the highest yield was 2.8 kg/plant under PO in the November planting. Yields were significantly higher under TBF for the July planting and significantly higher under PO for the November planting. We found no significant difference between greenhouses for the other cultivations. There was no significant difference in the number of harvested fruits between planting seasons or greenhouses, with a minimum of 10.7 fruits/plant under TBF in the August planting and a maximum of 15.0 fruits/plant under PO in the November planting (Table 4). There was no difference between greenhouses in terms of in fruit weight or maturity date, which was the number of days from anthesis to harvest. The maturity date increased as the cultivation season grew later. The weight of fruit was heavier in the October and November plantings than in the June and July plantings in both greenhouses. SSC ranged from 5.1 to 6.0% for all cultivations, and there was no correlation with fruit weight. The rate of marketable fruit under TBF was higher than that under PO in the June, July, and August plantings (Table 5). In the October and November plantings, the rate of marketable fruit was higher under PO than under TBF. In the June, July, and August plantings, the rate of fruit cracking was particularly high under PO. In both greenhouses, the rate of puffy fruit was high in the October and November plantings. Table 6 shows the cumulative intercepted PAR and LUE for each cultivation. Intercepted PAR was lowest in the October planting and highest in the July planting under both TBF and PO. LUE was higher under TBF than under PO, except for the June planting. In both greenhouses, LUE declined as the plants encountered longer periods of air temperatures above 31°C (Fig. 4). However, when the air temperature exceeded 31°C, the LUE decreased sharply for PO compared with TBF.

Discussion
Tomato cultivation is strongly affected by the cultivation season, and particularly by changes in air temperature and solar radiation. During the summer in the subtropical region of Ishigaki island, the maximum temperature under TBF was up to 3.6°C lower than that under PO, and the mean temperature was 0.4°C lower. TBF was more effective than PO at reducing the air temperature inside the greenhouse under conditions of high heat and high solar radiation. In the July planting, the yield under TBF was significantly higher than that under PO.
In this experiment, the cultivation season affected tomato plant growth. Under both TBF and PO, when the mean relative humidity was between 80.1 to 84.3%, the stem elongation rate decreased as the mean air temperature of the cultivation increased (Fig. 2). According to Ito (1971), photosynthetic activity in tomato was influenced by leaf temperature, and its peak was at approximately 28°C with a normal carbon dioxide supply. Ito also noted that the leaf temperature was 0.7°C higher on average than air temperature in the range of 11°C–35°C. In this study, we used a different film than Ito and did not measure leaf temperature, so we cannot directly compare the results; however, this indicated that high temperatures reduced both the photosynthesis rate and the amount of photosynthate, resulting in the slower stem elongation rate in our experiment. We transplanted the seedlings with six true leaves in July, but at the time of pinching, the number of leaves was the highest among all cultivations and the cultivation period was 97 days (Table 1), which was shorter than that of all plantings except the June planting. In both greenhouses, the leaf appearance rate was fastest in the July planting (Table 2). Heuvelink et al. (2018) pointed out that the leaf appearance rate linearly increased with increasing temperatures in the range of 17°C–23°C. Our findings supported Heuvelink et al. (2018), because the mean temperature of the July planting was 23°C. The leaf appearance rate of the June planting, for which the mean temperature was above 25°C, was lower than was that of the November planting, for which the mean temperature was 18°C. The October planting had a slightly higher mean temperature than the November planting, and the leaf appearance rate was also higher. This indicated that leaf development was reduced by high mean temperatures of approximately 25°C. Savvides et al. (2014) pointed out that the local photosynthate availability is likely to determine the leaf initiation rate. The photosynthesis rate is reduced in high-temperature environments where the daytime temperature exceeds 30°C (Heuvelink and Dorais, 2005), as in the June planting. However, both greenhouses had the lowest leaf appearance rate in the August planting among the five cultivations, and the reason for this was not clarified in this study. Further study is needed. The LAI did not differ between greenhouses in the same cultivation season. Compared to the other cultivations, where the LAI ranged from 3.9 to 5.4, the LAI of the June planting was significantly lower, at 3.0 under TBF and 3.4 under PO. As reported by Higashide (2018), a yield increase due to increased LAI is expected when the LAI is below 3.0, but the yield does not increase when the LAI is above 4.0. Hosoi (2001) reported that the best LAI was five with the maximum yield in warmer seasons with high solar radiation and long hours of sunshine, whereas the LAI was four in cooler seasons with less solar radiation and short hours of sunshine. In our experiment, we considered LAI to be within the appropriate range for all cultivations. The SLA varied according to the cultivation season; it was highest under both TBF and PO in the November plantings grown in winter, followed by the October, August, July, and June plantings. For TBF, the SLA was higher than that for PO in the same cultivation season. We considered this to be an effect of PAR under TBF being lower than under PO. According to Fan et al. (2013), SLA decreased when the photosynthetic photon flux density increased from 50 to 550 μmol·m−2·s−1. Li et al. (2014) reported that the enhancement of crop photosynthesis in diffuse light depends not only on the direct effects of a more uniform vertical and horizontal light distribution, but also on the physiological and morphological acclimation of the crop. As diffused light reduced leaf temperature, they concluded that diffused light minimized the potential for photodamage or photoprotection arising from the overexcitation of the photosynthetic apparatus. Although we did not measure leaf temperature in our experiment, we considered TBF to reduce leaf temperature due to the reduction of near-infrared radiation and PAR. The effect may be similar to that of diffused light. SLA has been reported to be in the range of 113 cm2·g−1–131 cm2·g−1 for tomato cultivation in Thailand, a humid tropical region (Kleinhenz et al., 2006), and from 175 cm2·g−1–250 cm2·g−1 in the Dutch summer (Heuvelink, 1995). In our experiment, the growing environment of the June planting was similar to that of Thailand, whereas that of the October and November plantings was similar to that of the Dutch summer, particularly with regard to air temperature and solar radiation.
TDM was significantly smaller in the June planting under TBF, and we observed no difference in the other cultivation seasons (Table 3). According to Adams et al. (2001), 44.4%, 61.2%, 61.8%, and 29.1% of the dry matter was partitioned to fruits at 14°C, 18°C, 22°C, and 26°C, respectively, and they concluded that the lower distribution to fruits was due to poor fruit set at high temperature. Kleinhenz et al. (2006) reported that in tomato production in the tropical lowlands in Thailand, a low percentage (16%–19%) of TDM was distributed to fruits under a mean temperature range of 26.5°C–29.6°C. In our study, the rate of distribution to fruits exceeded 46% even when the mean temperature was 26°C. This may be due to cooling at nighttime. Kawasaki and Ahn (2015) reported that cooling at nighttime increased pollen viability, fruit number, fruit weight, and marketable fruit yield. The distribution to fruit under TBF was higher than that under PO in the same cultivation season, except for the November planting. The difference was particularly large in the July and August plantings, with a difference of 10% and 5%, respectively. Heuvelink (1997) reported that the generative sink strength is strongly dependent on the number of fruits per truss. However, in our experiment, the number of fruits was approximately the same in all cultivations (Table 4). Therefore, factors different from the number of fruits were thought to have affected the distribution rate to fruit. Sato et al. (1994) reported that tomato plants under 45% shade partitioned the highest proportion of dry matter to fruit among four shade levels during the harvesting period. We considered that the shading effect of TBF contributed to an increase in the distribution rate to fruit. Compared to PO, TBF did not appear to reduce the rate of distribution to fruit under high temperature and intense solar radiation. Yoshida and Nakazono (2006) noted that 50% of shading was effective in avoiding stress under severe light exceeding 350 W·m−2. Hamamoto et al. (2000) also reported that shading increased the distribution rate to fruit and reduced the distribution rate to roots during the period of fruit enlargement in tomatoes. They attributed this to plants prioritizing fruit growth over other growth under low solar radiation. In our experiment, the distribution rate to side shoots was higher under PO in all cultivation seasons (Table 3). Side shoot development and elongation increased significantly after pinching. This appeared to be due to a reduction in sink strength resulting from pinching the main stem and to the excess photosynthate distributed to side shoots. In both greenhouses, tomato plants showed higher distribution to leaves and stems in June, July, and August plantings than in October and November plantings. However, LAI did not differ between July, August, October, and November plantings. Furthermore, SLA was lower in June and July plantings, confirming a trend toward thicker leaves in summer cultivations.
Yield per plant was lowest for TBF in the June planting and highest for PO in the November planting (Fig. 3). In the same cultivation season, yields under TBF were significantly higher than those under PO in the July planting, and yields under PO were significantly higher than those under TBF in the November planting. As the number of harvested fruits per plant was almost the same for all cultivations, we can infer that the difference in yield was largely due to fruit weight (Table 4). As with yield, fruit weight was also significantly heavier in the November planting than in the June planting. We observed the same trend for maturity date, with lower mean temperatures leading to a later maturity date and heavier fruit weights. In our experiment, SSC was not correlated with either maturity date or fruit weight. The rate of marketable fruit was higher under TBF in June, July, and August plantings. Under PO, there was a higher rate of fruit cracking during the same period. There have been various studies of the causes of fruit cracking, such as the effects of severe solar radiation (Suzuki and Yanase, 2005; Suzuki et al., 2007, 2009) and daily temperature differences (Kimura et al., 2012). The effectiveness of shade curtains (Nomura et al., 2005; Okazaki and Ohta, 2006) and UV-cut films (Kimura et al., 2012) as a countermeasure to fruit cracking has also been reported. Wada et al. (2006) reported that decreasing the solar radiation to 5–6 MJ·m−2 by shading could effectively increase the marketable fruit yield when the average air temperature exceeds 25°C. In our study, TBF reduced the rate of fruit cracking in summer. Conversely, the marketable fruit rate under PO was higher than that under TBF in the October and November plantings. Both TBF and PO produced puffy fruits in the October and November plantings. The occurrence of puffy fruit was increased both by high plant density and number of fruits per plant and by shading (Takao and Tanaka, 1984), and it is thought to be due to lack of photosynthate. In our experiment, the cumulative PAR for PO in the October and November plantings was almost the same as that for TBF in the July and August plantings (Table 1). This suggests that there was no PAR deficiency in our experiment. Meanwhile, in the October and November plantings, the hours of sunlight were shorter, at 4.4 h and 4.0 h, respectively, than in the other cultivations. This indicated that a short duration of sunlight was responsible for the occurrence of puffy fruit. On the other hand, puffy fruit occurred in 6.8% and 7.7% in TBF in the July and August plantings, respectively. This may have been due to other factors, and further study is needed. Malformed fruit were more common in the June and July plantings under PO and in the October and November plantings under TBF. Saito and Ito (1967) reported that higher nighttime temperature and/or lower light intensity retarded the morphological development of flowers, thus resulting in the formation of smaller flowers with smaller sepals, petals, ovaries, and, especially anthers, followed by heavy flower drop. This may also lead to the occurrence of malformed fruit. In addition, under TBF, the flowers of plants planted in October and November received insufficient photosynthate due to low solar radiation through shading of TBF combined with low seasonal hours of sunshine. Kimura et al. (2012) reported that dense planting reduced fruit cracking, but increased puffy fruit. Our plant density was 4.25 plants·m−2, and we considered this to be too dense in the October and November plantings. The marketable fruit rate was higher under TBF than under PO in the June, July, and August plantings, when the mean temperature was 21°C and above and the cumulative PAR under PO was 603 MJ·m−2 and above. Therefore, TBF is suitable for use in summer in subtropical environments.
Under both TBF and PO, cumulative PAR was lowest in the October planting and greatest in the June planting (Table 1). LUE ranged from 2.09 to 3.11 in all cultivations (Table 6). Under TBF, LUE was higher than that under PO in the same cultivation season, except in the June planting, as the difference in TDM between TBF and PO was small and TBF had received less intercepted PAR.
Tomato cultivation under TBF reduced the damage caused by intense solar radiation during summer in this subtropical area. It has been suggested that TBF could improve cultivation under high temperature and high solar radiation, such as is found in tropical areas. Besides, TBF could be used in large modern greenhouses and for other crops. Under TBF, there was no reduction in either TDM or yield even at low PAR in winter. This suggests that sufficient photosynthesis could be achieved even at the low intercepted PAR under TBF in the October and November cultivations because of limited sink strength, as we pinched the stem at the second upper leaf above the fourth truss and picked the fruits, leaving four fruits on each truss. Under TBF, there could be sufficient PAR to distribute photosynthate to each part of plant due to a limited sink and abundant solar radiation in a subtropical summer. These results indicate that TBF and sink-limited cultivation are effective systems for high temperature and high solar radiation environments, such as tropical and subtropical regions. In the future, we need to consider cultivation systems that respond to seasonal changes, such as low sunshine hours in winter, in subtropical areas to improve greenhouse cultivation in the subtropics.
Literature Cited
- Adams, S. R., K. E. Cockshull and C. R. J. Cave. 2001. Effect of temperature on the growth and development of tomato fruits. Ann. Bot. 88: 869–877.
- Fan, X. X., Z. G. Xu, X. Y. Liu, C. M. Tang, L. W. Wang and X. I. Han. 2013. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 153: 50–55.
- FAOSTAT. 2018. <http://www.fao.org/faostat/en/#data/QC>.
- Fukuoka, N., D. Masuda and Y. Kanamori. 2009. Effects of temperature around the fruit on sugar accumulation in watermelon (Citrullus lanatus (Thunb.) Matsum. & Nakai) during the latter half of fruit developmental period. J. Japan. Soc. Hort. Sci. 78: 97–102.
- Hamamoto, H., Y. Shishido, T. Uchiumi and H. Kumakura. 2000. Effects of low light intensity on growth, photosynthesis and distribution of photoassimilates in tomato plants. Environ. Control in Biol. 38: 63–69.
- Heuvelink, E. 1995. Growth, development and yield of a tomato crop: periodic destructive measurements in a greenhouse. Sci. Hortic. 61: 77–99.
- Heuvelink, E. 1997. Effect of fruit load on dry matter partitioning in tomato. Sci. Hortic. 69: 51–59.
- Heuvelink, E. and M. Dorais. 2005. Crop growth and yield. p. 85–144. In: E. Heuvelink (ed.). TOMATOS. CABI publishing. Wallingford
- Heuvelink, E. and R. C. O. Okello. 2018. Developmental processes. p. 59–88. In: E. Heuvelink (ed.). Tomatoes, 2nd edition. crop production science in horticulture series. CABI, Oxfordshire.
- Hiei, K., M. Ito, Y. Ban and T. Oyabu. 2015. Effects of super-micro-fog spraying combined with shading on suppressing atmospheric temperature increase in greenhouse tomato cultivation under natural ventilation. Res. Bull. Aichi Agric. Res. Ctr. 47: 41–49 (In Japanese with English abstract).
- Higashide, T. 2018. Review of dry matter production and light interception by plants for yield improvement of greenhouse tomatoes in Japan. Hort. Res. (Japan) 17: 133–146 (In Japanese with English abstract).
- Higashide, T., I. Gotoh, K. Suzuki, K. Yasuba, K. Tsukazawa, D. Ahn and Y. Iwasaki. 2012. Effects of pinching and lowering on cucumber yield and yield components. Hort. Res. (Japan) 11: 523–529 (In Japanese with English abstract).
- Hosoi, N. 2001. Studies on optimal leaf area for yield in tomato plants grown in hydroponic greenhouses. Bull. National Institute of Vegetable and Tea Science 16: 329–349 (In Japanese with English abstract).
- Ide, O., T. Moriyama, K. Ryu and K. Oku. 2007. Effects of forced ventilation with fan on reduction of inside air temperature in greenhouse and growth and contents of lycopene of tomato fruit. Bull. Fukuoka Agric. Res. Cent. 26: 51–55 (In Japanese with English abstract).
- Ito, T. 1971. Photosynthetic activity of vegetable plants and its horticultural significance I. examination of the methods in measuring the photosynthesis with assimilation chamber. J. Japan. Soc. Hort. Sci. 40: 35–40 (In Japanese with English abstract).
- Itoh, M., C. Goto, Y. Iwasaki, W. Sugeno, D. H. Ahn and T. Higashide. 2020. Modeling and prediction of dry matter production by tomato plants in year-round production based on short-term, low-truss crop management. Hort. J. 89: 417–424.
- Kawasaki, Y. and D. H. Ahn. 2015. Local cooling at night around flowering trusses improved yield of tomato grown under hot conditions. J. SHITA 27: 137–143 (In Japanese with English abstract).
- Kawasaki, Y. and Y. Yoneda. 2019. Local temperature control in greenhouse vegetable production. Hort. J. 88: 305–314.
- Kawashima, H., M. Takaichi and K. Yasuba. 2011. Performance of air-to-air heat pump for tomato greenhouse cooling and reduction of night-time cooling load by mulch to control soil heat flux. Bull. National Institute of Vegetable and Tea Science 10: 95–104 (In Japanese with English abstract).
- Kimura, M., S. Fujitani and K. Itimanda. 2012. Mitigation techniques on fruit cracking in tomato cultivation under rain shelter in summer and autumn. Bull. Oita Pref. Agri. Forest. Fish. Res. Cent. 2: 23–42 (In Japanese).
- Kleinhenz, V., K. Katroschan, F. Schutt and H. Stutzel. 2006. Biomass accumulation and partitioning of tomato under protected cultivation in the humid tropics. Europ. J. Hort. Sci. 71: 173–182.
- Li, T., E. Heuvelink, T. A. Dueck, J. Janse, G. Gort and L. F. M. Marcelis. 2014. Enhancement of crop photosynthesis by diffuse light: quantifying the contributing factors. Ann. Bot. 114: 145–156.
- Luthria, D. L., S. Mukhopadhyay and D. T. Krizek. 2006. Content of total phenolics and phenolic acids in tomato (Lycopersicon esculentum Mill.) fruits as influenced by cultivar and solar UV radiation. J. Food. Compos. Anal. 19: 771–777.
- Nomura, Y., T. Suzuki and T. Enya. 2005. Control of the tomato fruit cracking by shading materials in summer autumn. Bulletin of the Gifu Prefectural Research Institute for Agricultural Sciences in Hilly and Mountainous Areas 5: 11–16 (In Japanese with English abstract).
- Okazaki, T. and H. Ohta. 2006. Effects of shading on growth and yield of summer to autumn harvesting tomato. Tohoku Agric. Res. 59: 185–186 (In Japanese).
- Saito, T. and H. Ito. 1967. Studies on the growth and fruiting in the tomato. IX. Effects of the early environmental conditions and the cultural treatments on the morphological and physiological development of flowers and the flower drop. (I). effects of night temperature, light intensity and fertility of bed soil. J. Japan. Soc. Hort. Sci. 36: 195–205
- Sato, H., T. Yanagi, H. Hirai, Y. Ueda and Y. Oda. 1994. Effects of shading on growth, fruit yield and dry matter partitioning of single truss tomato plants. Environ. Control in Biol. 32: 231–237 (In Japanese with English abstract).
- Savvides, A., N. Ntagkas, W. Ieperen, J. A. Dieleman and L. F. M. Marcelis. 2014. Impact of light on leaf initiation: a matter of photosynthate availability in the apical bud? Funct. Plant Biol. 41: 547–556.
- Suzuki, K. 2019. Physiological disorders and their management in greenhouse tomato cultivation at high temperatures. p. 81–96. In: T. Izumi, R. Hirata, R. Matsuda (eds.). Adaptation to climate change in agriculture. Springer Nature Singapore Pte Ltd. Singapore.
- Suzuki, K., K. Mizukami, K. Tsuchiya, K. Yasuba, Y. Nakano and M. Takaichi. 2011. Control of excessive stem elongation in nursery-grown tomatoes, and the yields with low-node-order pinching system at high planting density. Hort. Res. (Japan) 10: 183–189 (In Japanese with English abstract).
- Suzuki, T. and S. Yanase. 2005. Effects of each irrigation method and training system on radial fruit cracking in summer and fall culture of tomatoes under plastic canopy. Hort. Res. (Japan) 4: 75–79 (In Japanese with English abstract).
- Suzuki, T., Y. Nomura, T. Shimazu and I. Tanaka. 2009. Effects of fruit thinning, covering of the fruit truss and CO2 enrichment on radial fruit cracking in tomato production under rain shelter in cool uplands. Hort. Res. (Japan) 8: 27–33 (In Japanese with English abstract).
- Suzuki, T., S. Yanase, T. Enya, T. Shimizu and I. Tanaka. 2007. Effects of total integrated solar radiation on radial fruit cracking in tomato cultivation under rain shelter in cool uplands. Hort. Res. (Japan) 6: 405–409 (In Japanese with English abstract).
- Takao, M. and Y. Tanaka. 1984. Studies on the preventive measures for the puffy fruits of tomato plants in vinyl house (2) effect of the growth environments of field, defoliation and fruit thinning in forcing tomato culture to development of the puffy fruits. Bull. Fukuoka Agric. Res. Cent. B-4: 31–36 (In Japanese with English abstract).
- Wada, T., H. Ikeda, H. Hirai and Y. Nishiura. 2013. Simulation model for predicting fruit yield of tomatoes grown on a single-truss system under shade in summer. Environ. Control. Biol. 51: 11–16.
- Wada, T., H. Ikeda, K. Matsushita, A. Kambara, H. Hirai and K. Abe. 2006. Effects of shading in summer on yield and quality of tomatoes grown on a single-truss system. J. Japan. Soc. Hort. Sci. 75: 51–58.
- Yoshida, Y. and T. Nakazono. 2006. Development of tomato fruits as affected by automatic shade control for eliminating the excessive solar radiation. Hort. Res. (Japan) 75 (Suppl. 2): 290 (In Japanese).