Abstract
The effects of End-of-Day (EOD)-cooling on floral differentiation and cut flower characteristics in carnation were verified using both short-term model-based experiments in a growth chamber and long-term demonstrative experiments in a greenhouse. In the model experiment, flower buds were undifferentiated until 43 days after pinching in both control and End-of-Night-cooled plants. In contrast, in the EOD-cooling treatment, every set reached the reproductive phase by 22 days after treatment. At the same node position, a three-stage advancement in floral differentiation was observed after EOD-cooling, but not after the other two treatments. This suggested that EOD-cooling promoted both floral differentiation and development in carnation. For the demonstrative experiment, significant differences between EOD-cooled and control plants varied each year in days to flowering, node order of flowering, and stem weeping angle. Because environmental factors could affect variation, correlations between the diurnal temperature range and the difference in each survey component were examined using regression analyses. Moderate (days to flowering) and a strong (node order of flowering) correlations were found with the diurnal temperature range. Thus, the EOD-cooling treatment was advantageous in suppressing delayed flowering over a large diurnal temperature range during floral differentiation. The difference in stem weeping angle showed a strong correlation with the difference in minimum temperature during the stem elongation stage. Improving the stem quality by EOD-cooling tended to be effective under high minimum-temperature conditions.
Introduction
The optimum growing temperature for carnation (Dianthus caryophyllus L.) is 18–24°C, and they prefer relatively cooler climates (Higashiura et al., 2020). Therefore, it is difficult to grow high-quality carnations systematically for cutting during the hot season in Japan. Additionally, delayed flowering and soft stems are serious problems under abnormally high-temperature conditions in summer and autumn (Yamaguchi, 1991). Delayed flowering reduces cut flower yields, and soft stems decrease the product value. As a countermeasure to these problems, the use of heat pumps to cool greenhouses at night while growing the standard carnation ‘Excerea’ has been investigated. In our previous report, an overnight cooling at 21°C improved the quality of cut flowers by hardening stems and promoting flowering (Higashiura et al., 2020). Furthermore, an End-of-Day (EOD)-cooling treatment for 4 h after sunset was as effective as overnight cooling, thereby reducing electricity consumption.
The effects of EOD-cooling on both the quality and quantity of cut flowers were also confirmed in rose (Kajihara et al., 2015). However, unlike in carnation, flowering in rose was not promoted. EOD treatments, in which the temperature is increased for a few hours after sunset and lowered afterward, have also been used to lower the cost of winter heating in greenhouses. This method promoted flowering in marigold (Douzono et al., 2012), chrysanthemum (Kawanishi et al., 2012), and carnation (Baba et al., 2013). In particular, in marigold, an EOD-heating at 30°C for 3 h followed by cooling at 14°C for the rest of the dark period promoted the differentiation and early development of flowers, compared with a treatment of 14°C for the entire dark period, in an artificial chamber. Moreover, the effects of an End-of-Night (EON) heating at 30°C for 3 h were limited compared with those of the EOD treatment. Similarly, the acceleration in flowering time was greater after EOD-cooling than after EON-cooling (Higashiura et al., 2020). However, it is unknown whether this promotive effect is a result of accelerated floral differentiation.
Flowering in carnation is strongly influenced by temperature, with higher daytime temperatures and lower nighttime temperatures resulting in fewer flowering nodes and a shorter time to flower bud differentiation. Dahab (1967) examined combinations of daytime and nighttime temperatures at five levels, between 12 and 24°C, and found that fewer days were required for flower bud differentiation at higher daytime and lower nighttime temperatures. The effect of nighttime cooling on flower bud differentiation was strongly influenced by the daytime temperature. Accordingly, it is essential for practical applications to clarify the factors affecting EOD-cooling in carnation, such as daytime and nighttime temperatures and sunlight conditions.
In this study, we attempted to investigate the effects of EOD- and EON-cooling on flower bud development under artificial conditions by observing flower bud developmental stages. Additionally, as a practical test in a greenhouse, trials were repeated for five consecutive years from 2013 to 2017 to verify the effects of EOD-cooling on both the promotion of flowering and quality of cut flowers grown during the summer season.
Materials and Methods
Experiment 1
The standard-type carnation (Dianthus caryophyllus L.) ‘Excerea’ was used in all the experiments. The rooted cuttings were planted in 7.5-cm white plastic pots filled with mixed soil [65% peat moss, 15% decomposed granite soil, 10% perlite, 10% Akadama (red granular) soil] on December 15, 2017. Planted pots were kept in a glass greenhouse at the Awaji Agricultural Technology Center (34.31 NE, 134.80 E) at 25°C using an air conditioner throughout the day and night. The top buds were pinched a month after planting, leaving five nodes. Each treatment was performed on 96 plantlets, which were placed in three growth chambers (MLR-352H-PJ; PHC Corporation, Tokyo, Japan) with 12-h light periods (PPFD 180 μmol·m−2·s−1). The temperature inside the chamber was set at 30°C. The short-term cooling treatments were set at 21°C for 4 h after the end or before the beginning of light periods as EOD or EON treatments, respectively. Each plantlet was trimmed to leave one lateral shoot. Tap water was poured into the stainless-steel tray in which the pots were placed for bottom watering. A week after planting, one Promic fertilizer tablet (N:P:K = 8:8:8; Hyponex Japan Corp., Ltd., Osaka, Japan) was added to each plastic pot.
Primary lateral branches were collected from each stock once a week from 6 to 8 plants per treatment until 85 days after treatment for microscopic observation of flower buds. The samples were collected from approximately 10-mm apices and fixed in formalin, acetic acid, and alcohol solution. Subsequently, the solution was replaced with 70% ethanol, and samples were stored until observed using a stereomicroscope. Flower bud development was divided into five stages, undifferentiated bud (1), domed primordium (2), sepal forming (3), petal forming (4), and stamen and pistil forming (5) (Fig. 1), in accordance with previous classifications (Furusato et al., 1967). The bud stage (‘6’ in Fig. 2) was determined by visual observation of a flower bud forming. In addition, the number of nodes with expanded leaves per individual plant was recorded.
Experiment 2
The standard-type carnation ‘Excerea’ was grown five times beginning in June of each year from 2013 to 2017. For all the cultivations, a polypropylene bed (0.85-m wide × 2.0-m long × 0.17-m deep; Drain-bed 85; National Federation of Agricultural Cooperative Associations, Tokyo, Japan) filled with mixed soil [65% peat moss, 15% decomposed granite soil, 10% perlite, 10% Akadama (red granular) soil] was installed in a glass greenhouse at the Awaji Agricultural Technology Center.
The seedlings were planted in mid-June (on the 18th to 20th). A trellis net with eight lanes was used to prevent lodging, with rooted cuttings occupying six lanes. The two center lanes remained unplanted. All the beds were mulched with photo-reflective film (Poly-Shine NF; Hitachi AIC, Tochigi, Japan).
Fertilizer was applied as nutrient solution at 1,000 to 1,400 × dilutions (Youekidokou-2-gou; OAT Agrio Co., Ltd., Tokyo, Japan), including 124 g of nitrogen, 65 g of phosphoric acid, and 214 g of potassium chloride per m2 (actual cropping area). At three weeks after planting, in mid-July, the top buds were pinched to leave five nodes, and in mid-August, each plantlet was trimmed to leave four lateral shoots.
Two separate greenhouses (3 m wide × 4 m deep × 3 m high) were used for the two treatments: one was cooled at 21°C for a short time after sunset (EOD-cooling), and the other was a non-cooled control. The start time of the cooling system was adjusted every week based on the sunset time in Kobe City, Japan. The treatment began on the day of pinching and was completed on September 25 in all years. An air conditioner [S50NTEP (5 kW); Daikin Industries, Ltd., Osaka, Japan] was used for cooling. In the non-cooled greenhouse, a controlled ambient temperature was maintained at all hours.
There were three replicated bulks of 11 to 12 strains per treatment area. The first flowers from the primary lateral branch were harvested at eight nodes from the top when the outer petals emerged horizontally. Days to flowering were counted from the day the top bud was pinched to the day that the primary cut flower was harvested until the end of December. In addition, the number of nodes from the branching position of the primary lateral branch and the stem weeping angle (SWA) were investigated for all collected cut flowers. The SWA, the angle between a horizontal line and a line connecting the center of the bud to the held position, was measured at 45 cm from the flower top.
The temperatures inside the greenhouses were measured using a temperature sensor (TR-0106; T&D Corporation, Nagano, Japan). The outside temperatures were recorded using a TR-0106 in 2013 and 2014, a TS-801 (WINS®; Meisei Electric Co., Ltd., Gunma, Japan) in 2015, and an HT-20 (Azbil Corporation, Tokyo, Japan) in 2016 and 2017. Each recorder was placed inside a ventilation duct (TR-0106 and HT-20) or weather observation system (TS-801). All the temperatures were recorded at 10-min intervals. The hours of sunshine were obtained from the Japan Meteorological Agency’s AMeDAS data (Sumoto, Hyogo, Japan).
Statistical analyses
Statistical analyses were performed using EXCEL toukei ver. 7.08 (ESUMI Co., Ltd., Tokyo, Japan). We determined the significance of differences using the Student’s test or Tukey–Kramer’s HSD test. Data shown in Figures and Tables are the mean ± standard error of three biological replicates consists of 12-bulked samples.
Results
Experiment 1
To confirm the short-term cooling effects on floral differentiation in carnation, a small-scale experiment was conducted with young plants in an artificial climate chamber. Floral differentiation was divided into five previously reported classification stages (Furusato et al., 1967) using microscopic observations (Fig. 1). In both control and EON treatments, some flower buds were undifferentiated (stage 1) until 43 days after pinching. In contrast, in the EOD treatment, every set reached the domed primordial stage (stage 2), which was defined as flower differentiation by Furusato et al. (1967), by 22 days (Fig. 2). At 71 days, all the EOD-treated plantlets had reached at least the sepal-forming stage (3), which indicates distinct flower differentiation (Emino and Rasmussen, 1971). The EON treatment delayed the transition to this stage until 78 days. During the EOD treatment, some individuals proceeded to either the petal-forming stage (4) or the ovule- and pollen-forming stage (5) at 71–78 days, and almost all individuals had reached these stages at 85 days. No sets reached bud stage before 85 days under either control or EON-treatment conditions. Floral differentiation at lower node positions occurred significantly earlier in plants receiving the EOD-cooling treatment compared with those receiving the other treatments (Fig. 2), despite similar increases in the numbers of nodes over time in the two other treatments (Fig. S1). Changes in floral developmental stages, along with increased numbers of nodes per treatment, are shown in Figure 3. Noticeable differences were identified at more than 17 nodes. Plants receiving the control and EON treatments remained at stages 2 and 3, while those receiving the EOD treatment advanced to stage 4 or later. Thus, EOD-cooling promoted both floral differentiation and development.

A long-term verification test of the EOD treatment was carried out in a glass greenhouse. Table 1 shows comparisons of temperatures between EOD-cooling and non-cooling, with outside temperatures as references, for five years. The original data were presented every five days (pentads) (Table S1). On the basis of the outside temperature data, 2013, 2016, and 2017 had hotter seasons, with average temperatures of greater than 27°C, while 2015 had a cool season, with the lowest average, maximum, and minimum temperatures. EOD-cooling, but not non-cooling, generally lowered the minimum temperature to less than 21°C, which was the preset temperature of the air conditioner. The hours of sunshine during the treatment days were long in 2013 (585.8 h) and 2016 (558.4 h), but short in 2014 (426.0 h) (Table S2). The effects of EOD-cooling on some agricultural characteristics observed in the five-year trial are summarized in Table 2. The days to flowering were significantly affected by EOD-cooling, being 15 and 13 days in 2013 and 2014, respectively. There was not much difference in other flower characteristics, such as a flower length between the EOD and non-cooling treatments from 2013 to 2017 (data not shown). EOD-cooling reduced the number of nodes at the primary branch compared with the number in the control (non-cooling) every year, with the decrease being significant in 2013, 2014, and 2015. Because stem firmness contributes to the quality of cut flowers, the SWA was measured at harvest. EOD-cooling produced firmer stems with smaller angles than control conditions in all the trial years, and the differences were especially significant in 2013 and 2016. These data suggested that the effects of EOD-cooling, notwithstanding environmental conditions that rarely masked the effects, may shorten the days to flowering and improve the quality of cut flowers.

Discussion
We reported previously that an EOD-cooling treatment reduces both the days to flowering and the number of nodes at flowering compared with the control (non-cooling) in carnation (Higashiura et al., 2020). This may result from the promotion of flower differentiation by short-term cooling. In Experiment 1, direct evidence was obtained using microscopic observations of the apical meristem of carnations grown in an artificial chamber. Both flower differentiation and development under EOD-cooling conditions occurred earlier than under non-cooling conditions (Fig. 2). Furthermore, the developmental stages of flower buds at the same node positions proceeded through three stages under EOD-cooling conditions compared with under either non-cooling or EON-cooling conditions. Thus, EOD-cooling in glass greenhouses promotes both bud initiation and flower development, which may increase flower blooming as previously reported (Higashiura et al., 2020).
Dahab (1967) reported that the lower the night temperature, the lower the node position of flowering in ‘William Sim’ carnation. The advancement of flower differentiation by EOD-cooling may therefore have resulted from the decreased average night temperature in Experiment 1 (Fig. 2). However, the EON-cooling treatment did not have the same effects on flowering as the EOD-cooling treatment, even though the same 4-h treatment period and preset temperature of 21°C were used. Similar results have been obtained from greenhouse experiments (Higashiura et al., 2020). As with African marigold, EOD-heating, but not EON-heating or non-heating, shortens the days required for both flower differentiation and bud formation (Douzono et al., 2012). They reported that the appropriate temperature in the early nighttime promotes the translocation of photosynthetic assimilates from the source to sink. In chrysanthemum, a high temperature delayed capitulum development with the maximum sensitivity towards the end of the night (Nakano et al., 2020). Previous reports have shown that daily fluctuations in heat sensitivity may correlate with the regulation of the FLOWERING LOCUS T (FT) ortholog (FTL3) in chrysanthemum (Nakano et al., 2013, 2015). This time- and temperature-dependent regulation of the florigen gene has been reported in Arabidopsis FT, whose transcript levels were increased at the end of cooler nights (Kinmonth-Schultz et al., 2016). In carnation, it is unclear whether EOD-timing and/or changes in temperature affect the expression level of the FT gene. Genome and post-genome data will help to understand this phenomenon in carnation (Yagi et al., 2014, 2020; Zhang et al., 2018).
The effects of nighttime cooling on flowering differ in response to outside temperature conditions. For Elatior begonia, the higher the day temperature, the greater the nighttime cooling suppression of delayed flowering (Nakashima, 2019). To verify the temperature-dependency of EOD-cooling in carnation, a continuous five-year trial was carried out in Experiment 2. Among the trial years, the significance levels of the differences varied for days to flowering, node order of flowering, and SWA (Table 2). Dahab (1967) reported that the difference in diurnal temperature enhances flower differentiation in carnation. The relationship between the difference in diurnal temperature range and the difference in days to flowering for carnation was examined using regression analysis (Fig. 4). Data were extracted during flower differentiation from 15 days after pinching to the beginning of the cool-night season. A moderate negative correlation (R2 = 0.664) was found for the difference in days to flowering. Similarly, for the difference in the node order of flowering, there was a strong significant negative linear correlation (R2 = 0.824) (Fig. 5). No correlation was observed for any duration, except the 1st to 5th pentad in August. Thus, we confirmed that the EOD-cooling treatment aided the suppression of delayed flowering over a large diurnal temperature range during the hot season. Partial cooling, which reduces cost, also has this effect. For Elatior begonia, only days having a maximum temperature of greater than 30°C were selected for nighttime cooling (Nakashima, 2019). This resulted in the same inhibitory effect on delayed flowering as cooling every night. The cost-effective promotion of flowering in carnation will be accomplished by further optimizing the timing, duration, and temperature conditions.


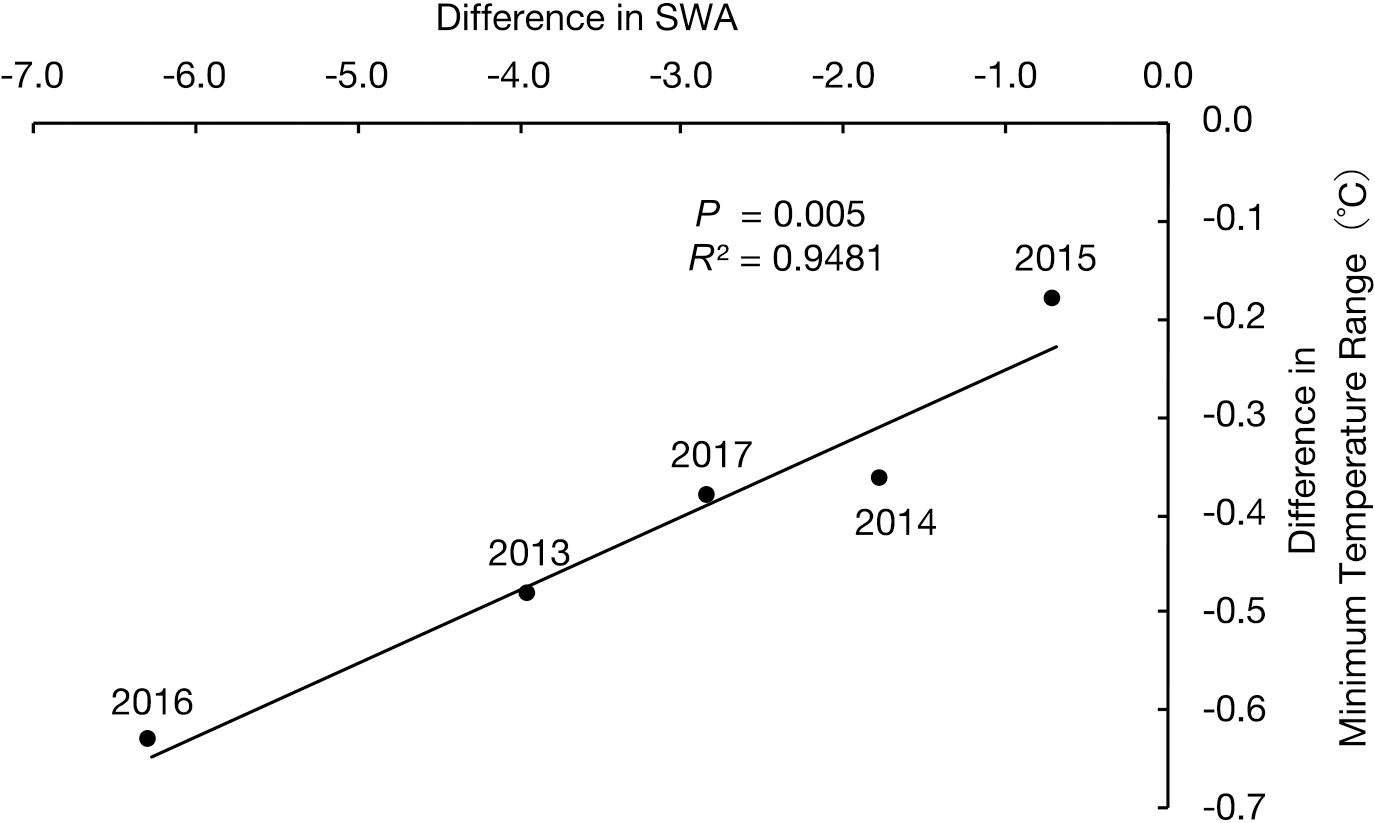
Generally, flower stems of cut carnations harvested during the hot seasons are much softer than those harvested in cooler seasons, which leads to a decrease in the commodity value (Bunt, 1978). The stem hardness of cut flowers was improved by EOD-cooling during the summer season, as indicated by the smaller SWA than in the non-cooled control group. The SWA was significantly smaller in plants receiving the EOD-cooling treatment compared with non-cooling in 2013 and 2016 (Table 2). However, no significant differences were found in the other years; therefore, the nature of the effect remains unclear. For tomato plants grown in a greenhouse with or without cooling during six hot seasons, the relationship between the difference in yield and the night temperature gap was validated by regression analysis (Willits and Peet, 1998). The greater the difference among the night temperatures, the greater the yield from tomato plants receiving the cooling treatment. With a 4°C decrease, an up to 40%–50% gain occurred. This may be caused by the enhanced translocation of assimilates in tomato at low temperatures. A similar effect, in which the low temperature-dependent stem hardiness is caused by the distribution of photosynthetic assimilates in the stem, has been reported in carnation (Harris and Jeffcoat, 1974). Therefore, we used regression analysis to determine whether there was a correlation between the difference in minimum temperature and the difference in SWA. Although no correlation was found between the temperature during the whole experimental period or the partial period from the 1st to 5th pentad in August and the difference in SWA, a strong positive correlation (R2 = 0.9481, P = 0.005) was found from the 6th pentad in August to the 5th pentad in September (Fig. 6). The duration is estimated to be stage 5 or 4 in floral differentiation with eight nodes about 50 days before flowering (Table 2). This suggested that the improvement in the stem quality by EOD-cooling occurs in a limited time when the stem is rapidly elongating, rather than the time immediately after pinching. In the case of rose, stem characteristics (fresh weight, dry weight, and diameter) showed a proportional gain with decreased night temperature (Hopper and Hammer, 1991). Hurd and Enoch (1976) reported a referential result that the growth of carnation was not promoted by low temperature in the night time in early the first 14 days after treatment, but was promoted from 14 to 35 days. Although the difference in minimum temperature was 0.6°C at most (Fig. 6), SWA could be reduced by integrating differences in low temperatures during an effective period. There was no correlation between DIF (difference between day and night temperature) and SWA (data not shown), as reported previously (Higashiura et al., 2020). The effect of EOD-cooling on the stem quality may be masked by other environmental factors, such as hours of sunshine (Table S2). Some moderate negative correlations were found between the difference in SWA and the integrated hours of sunshine during both the whole experimental period (R2 = 0.7485, P = 0.058) and the partial period from the 1st to 5th pentad in August (R2 = 0.8213, P = 0.034) (data not shown). This may result from the increased minimum temperature owing to the long hours of sunshine.
In conclusion, an inhibitory effect on delayed flowering during the hot season was caused by the EOD-cooling-induced promotion of bud initiation and flower development. Furthermore, this inhibition was more effective under increased minimum and maximum temperature conditions. The improvement in the stem quality by EOD-cooling was efficient in years having high minimum temperatures during the stem-elongation stage.
Literature Cited
- Baba, F., K. Ishii, H. Muto and Z. Inaba. 2013. Effects of short-term end-of-day or end-of-night heating on the flowering of winter-cultivated spray-type carnation (Dianthus caryophyllus L.) cultivars. Hort. Res. (Japan) 12: 389–396 (In Japanese with English abstract).
- Bunt, A. C. 1978. Effect of season on the carnation (Dianthus caryophyllus L.) III Flower quality. J. Hort. Sci. 53: 75–84.
- Dahab, A. A. M. 1967. Effects of light and temperature on growth and flowering of carnation (Dianthus caryophyllus L.). Ph.D. Thesis. Wageningen Univ., Wageningen.
- Douzono, M., T. Kando, T. Hisamatsu, A. Ohmiya, K. Ichimura and M. Shibata. 2012. Effect of end-of-day heating treatment on floral initiation and development in Tagetes erecta. Hort. Res. (Japan) 11: 553–559 (In Japanese with English abstract).
- Emino, E. R. and H. P. Rasmussen. 1971. Scanning electron microscope studies of the shoot apex in Dianthus caryophyllus L. cv. scania. Amer. Soc. Hort. Sci. J. 96: 253–256.
- Furusato, K., M. Yoshida and K. Hida. 1967. Study on flower bud initiation in carnation. Agric. and Hortic. 42: 1115–1116 (In Japanese).
- Harris, G. P. and B. Jeffcoat. 1974. Effects of temperature on the distribution of 14C-labelled assimilates in the flowering shoot of carnation. Ann. Bot. 38: 77–83.
- Higashiura, M., S. Kajihara, Y. Uno and M. Yamanaka. 2020. Effects of temperature and timing/duration of night cooling treatments on flowering time and quality of cut flowers of standard type carnation (Dianthus caryophyllus). Hort. J. 89: 61–68.
- Hopper, D. A. and P. A. Hammer. 1991. Regression models describing Rosa hybrida response to day/night temperature and photosynthetic photon flux. J. Amer. Soc. Hort. Sci. 116: 609–617.
- Hurd, R. G. and H. Z. Enoch. 1976. Effect of night temperature on photosynthesis, transpiration, and growth of spray carnations. J. Exp. Bot. 27: 695–703.
- Kajihara, S., S. Ishikura, K. Fukushima and M. Douzono. 2015. Effect of short-period air-conditioning during night on yield and characteristic form of cut roses in summer. Hort. Res. (Japan) 14: 365–369 (In Japanese with English abstract).
- Kawanishi, T., K. Shima, H. Hayashi, M. Douzono and T. Hisamatsu. 2012. Effect of short-term rising temperature treatment performed around the end of the day on the growth, flowering, and cut-flower quality of spray-type chrysanthemums. Hort. Res. (Japan) 11: 241–249 (In Japanese with English abstract).
- Kinmonth-Schultz, H. A., X. Tong, J. Lee, Y. H. Song, S. Ito, S. H. Kim and T. Imaizumi. 2016. Cool night-time temperatures induce the expression of CONSTANS and FLOWERING LOCUS T to regulate flowering in Arabidopsis. New Phytol. 211: 208–224.
- Nakano, Y., Y. Higuchi, K. Sumitomo and T. Hisamatsu. 2013. Flowering retardation by high temperature in chrysanthemums: involvement of FLOWERING LOCUS T-like 3 gene repression. J. Exp. Bot. 64: 909–920.
- Nakano, Y., Y. Higuchi, K. Sumitomo, A. Oda and T. Hisamatsu. 2015. Delay of flowering by high temperature in chrysanthemum: heat-sensitive time-of-day and heat effects on CsFTL3 and CsAFT gene expression. J. Hort. Sci. Biotech. 90: 143–149.
- Nakano, Y., T. Takase, K. Sumitomo, S. Suzuki, K. Tsuda-Kawamura and T. Hisamatsu. 2020. Delay of flowering at high temperature in chrysanthemum: duration of darkness and transitions in lighting determine daily peak heat sensitivity. Hort. J. 89: 602–608.
- Nakashima, T. 2019. Development of stable flowering and quality improvement of autumn cropping-type Begonia × hiemalis Fotsch. using night cold storage and intermittent low-temperature storage (In Japanese). Ph.D. Thesis. Okayama Univ., Okayama.
- Willits, D. H. and M. M. Peet. 1998. The effect of night temperature on greenhouse grown tomato yields in warm climates. Agric. For. 92: 191–202.
- Yagi, M., S. Kosugi, H. Hirakawa, A. Ohmiya, K. Tanase, T. Harada, K. Kishimoto, M. Nakayama, K. Ichimura, T. Onozaki, H. Yamaguchi, N. Sasaki, T. Miyahara, Y. Nishizaki, Y. Ozeki, N. Nakamura, T. Suzuki, Y. Tanaka, S. Sato, K. Shirasawa, S. Isobe, Y. Miyamura, A. Watanabe, S. Nakayama, Y. Kishida, M. Kohara and S. Tabata. 2014. Sequence analysis of the genome of carnation (Dianthus caryophyllus L.). DNA Res. 21: 231–241.
- Yagi, M., K. Shirasawa, H. Hirakawa, S. Isobe, J. Matsuno, Y. Uno, K. Tanase, T. Onozaki and H. Yamaguchi. 2020. QTL analysis for flowering time in carnation (Dianthus caryophyllus L.). Sci. Hortic. 262: 109053.
- Yamaguchi, T. 1991. Recent advances of carnation production in Japan. Japan Agric. Res. Q. 24: 265–272.
- Zhang, X., Q. Wang, S. Yang, S. Lin, M. Bao, M. Bendahmane, Q. Wu, C. Wang and X. Fu. 2018. Identification and characterization of the MADS-box genes and their contribution to flower organ in carnation (Dianthus caryophyllus L.). Genes (Basel). 9: E193.



