Abstract
Split pit in peach [Prunus persica (L.) Batsch] fruit is an internal disorder characterized by the splitting or fracture of the lignified endocarp known as the stone or pit. It is empirically known that split-pit peach fruit mature earlier than normal fruit on trees, thereby leading to frequent preharvest fruit drop in split-pit fruit. However, studies that accurately compare split-pit and normal fruit maturation and/or focus on the physiological mechanism underlying this difference in maturation are limited, partly due to the lack of an effective method to distinguish split-pit fruit from normal fruit on trees. Recently, we demonstrated that an acoustic vibration method could be effectively used for the nondestructive detection of split pit in unpicked peach fruit on trees. In this study, we applied this method to distinguish normal fruit from split-pit fruit and evaluate maturation on trees. Comparison of pairs of normal and split-pit fruit grown at a near-canopy position for three consecutive years (2018–2020) revealed that split-pit fruit matured earlier than normal fruit in most pairs. The average harvest date difference was approximately three days, at least under the experimental conditions in 2020. The amount of ethylene produced from seeds was significantly larger in immature split-pit fruit than immature normal fruit collected at fruit development stage from late June to early July, suggesting that the increased ethylene production from seeds during the fruit development stage is a possible cause of early maturation in split-pit fruit. From the results obtained, we discuss the effect of split pit on the maturation of peach fruit on trees in relation to practical cultivation management to reduce problems caused by split pit.
Introduction
Split pit is a problematic inner disorder typically observed in peach [Prunus persica (L.) Batsch] fruit. As in other Prunus species, peach has a lignified endocarp called a pit or stone at the center of the fruit, in which a seed is contained and protected so that the mature seed falls to the ground to produce offspring. In some peach cultivars, however, pit splitting or fracture occurs during fruit growth (Davis, 1941; Drogoudi et al., 2009; Evert et al., 1988; Miki, 1932; Ragland, 1934). Although the exact cause is unclear, outward forces generated by fruit enlargement during stage II (the period of pit lignification) and/or the early part of stage III (the period of rapid fruit enlargement towards maturity) of fruit growth are thought to pull apart the insufficiently hardened endocarp, resulting in split or fracture at a weak point along the suture line (Claypool et al., 1972; Kritzinger et al., 2017; Ragland, 1934; Woodbridge, 1978). Cultivation practices to produce large fruit, such as intense fruit thinning and pruning, girdling of bearing branches, and excess N2 fertilization, tend to increase split-pit occurrence (Burrell and Reighard, 2017; Claypool et al., 1972; Drogoudi et al., 2009; Fukuda et al., 2006; Kubota et al., 1993; Nakano and Nakamura, 2001). In fact, the size and weight of split-pit fruit were shown to often exceed those of normal fruit (Claypool et al., 1972; Evert et al., 1988). As split-pit fruit are considered to be of inferior fruit quality characterized by low soluble sugar content (SSC), a bitter taste, wooly texture, gumming and pit cracking, and a reduced shelf life (Crisosto et al., 1997; Kubota et al., 1993; Miki, 1932; Mizutani et al., 1998), the contamination of sorted fruit with split-pit fruit may reduce credibility in the market.
In addition to its adverse effects on postharvest properties, split pit is known to affect fruit maturation on trees. It is widely known that split-pit fruit tend to mature earlier than normal fruit on trees, and a high percentage of split-pit fruit is contained in fruit harvested at an early stage of the harvest period in commercial orchards. In this context, preharvest fruit drop as well as contamination of fruit in the advanced stage of maturation relative to the optimum harvest stage is considered to occur mostly in split-pit fruit, which leads to substantial yield reduction and thus has become a major issue for growers. Although this accelerated maturation on trees of split-pit fruit compared with normal fruit is empirically believed by peach growers, experimental evidence for this difference in maturation, as well as the physiological mechanism underlying it, is still unclear, partly because an effective nondestructive method to distinguish normal fruit from split-pit fruit on trees has not been developed until recently, as described below.
Aside from split-pit occurrence, peach maturation is greatly dependent on fruit position in the canopy; fruit grown at a higher position on trees tend to mature earlier in the harvest period than those at a lower position, regardless of split-pit occurrence. Therefore, it is necessary to compare normal and split-pit fruit pairs grown at a near-canopy position for a precise evaluation of the maturation-promoting effect of split pit. However, such a comparison has been difficult to date because there are no clear indications in most growing split-pit fruit, and thus it is difficult to estimate normal and split-pit fruit pairs grown at a near-canopy position by their appearance on the tree (Kritzinger et al., 2017; Ragland, 1934; Woodbridge, 1978).
As a possible solution to the experimental limitation described above, we demonstrated in previous studies that an acoustic vibration method could be successfully used for the nondestructive detection of split pit not only in harvested peach fruit, but also in unpicked peach fruit on trees (Kawai et al., 2018; Nakano et al., 2018). The acoustic vibration method has been developed and used to evaluate flesh firmness and/or optimal ripeness, as well as internal defects in a wide range of fruit and agricultural products (Abbaszadeh et al., 2015; Ali et al., 2017; Kadowaki et al., 2012; Murayama et al., 2006; Oveisi et al., 2014; Takahashi et al., 2010; Taniwaki and Sakurai, 2010; Taniwaki et al., 2009a, b, c, 2010; Terasaki et al., 2006, 2013), in which mostly single second resonant frequency (f2) of acoustic vibration data has been examined as an index of the internal physical traits of the fruit (Cooke, 1972; Drake, 1962; Kadowaki et al., 2012). In our previous experiments on peach fruit, the ratio of the third resonant frequency (f3) to f2 (f3/f2) was used as a good indicator of split pit based on evidence that f2 was shifted to a much lower value in split-pit fruit than normal fruit, whereas f3 showed only small shifts (Kawai et al., 2018; Nakano et al., 2018). Time-course monitoring of f3/f2 values in growing peach fruit could effectively estimate the accurate timing of split-pit occurrence during fruit development (Kawai et al., 2018). It was also revealed that most pit splitting occurred from early June to early July, and split pit at harvest could be predicted with high accuracy thereafter. These results show the high potential of the acoustic vibration method, which is used to predict normal and split-pit fruit pairs grown at a near-canopy position, to evaluate differences in maturation.
In this study, we conducted a precise comparison of the maturation of normal and split-pit fruit on trees by applying the acoustic vibration method for three consecutive years (2018–2020). The early maturation of split-pit fruit was confirmed by examining other ripening-related traits, such as ethylene production, flesh firmness, and acoustic vibration. Ethylene production and flesh firmness of immature fruit and ethylene production of seeds from fruit development stage to harvest period were further investigated to obtain new insights into the physiological mechanism of early maturation in split-pit fruit. From the results, the effect of split pit on the maturation of peach fruit on trees is discussed in relation to practical cultivation management to reduce problems caused by split pit.
Materials and Methods
Plant materials
Three 16-year-old (as of 2018) ‘Shimizu Hakuto’ peach trees [Prunus persica (L.) Batsch] grown at the Research Farm of the Faculty of Agriculture, Okayama University (Okayama, Japan) were used in this study. The trees were grown under a cover against rain, although rainfall outside the cover could affect tree water status through underground water influx. Conventional cultivation methods were used for tree management. The dates when 70–80% of flowers had bloomed were April 2, 7, and 2 in 2018, 2019, and 2020, respectively. The busiest time for ‘Shimizu Hakuto’ peach harvest in the Okayama area is concentrated over a period of 7–10 days from late July to early August, although some early- and late-ripening fruit are also harvested before and after that period, respectively.
Estimation of pairs of normal and split-pit fruit grown at a near-canopy position by acoustic vibration measurement
In our previous study using the acoustic vibration method for the same plant materials, we estimated that most pit splitting occurred from early June to early July (Kawai et al., 2018). We also showed that the split-pit fruit detection rate gradually increased towards the end of that period and pit splitting could be predicted with high accuracy (83.8–94.6%) thereafter when the f3/f2 cut-off value was set at > 1.4. Based on these results, we measured the acoustic vibration of developing fruit on July 8 and 10, July 9 and 10, and July 6 and 8 in 2018, 2019, and 2020, respectively, to prepare pairs of normal and split-pit fruit grown at a near-canopy position; we selected two adjacent fruit from the same lateral branch. These measurement dates were in the middle of stage III of fruit growth of ‘Shimizu Hakuto’ peach, which were 9 to 13 days before the onset of harvest in this study (Fig. 1). A portable system composed of a device for resonant frequency measurement (Model NSP-1; Applied Vibro-Acoustic, Higashi-Hiroshima, Japan), and a tablet personal computer was used to measure the acoustic vibration of fruit on tree (Kawai et al., 2018). A total of 142, 150, and 150 fruit (71, 75, and 75 pairs) were measured in 2018, 2019, and 2020, respectively, when the f3/f2 cut-off value was set at > 1.4. The position of each fruit pair on trees was randomly selected in this study. Tested fruit were covered with fruit bags after a sufficient number of normal and split-pit fruit pairs were predicted by the acoustic vibration method and used for maturation comparison. Fruit pairs in which either one or both fruit fell before harvest were excluded from the results, and 126, 120, and 88 fruit (63, 60, and 44 pairs) remaining on trees until harvest were used in the subsequent investigation.
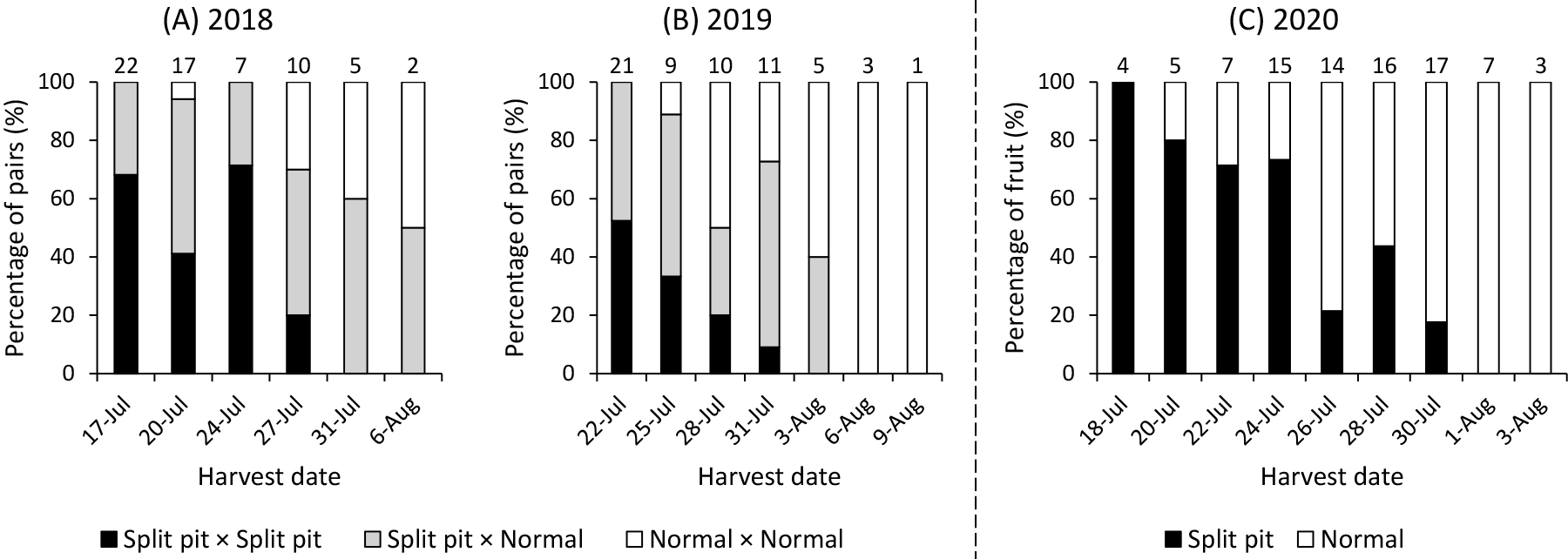
In 2018 and 2019, both fruit in each pair were harvested at the same time when either fruit reached commercial maturity. Commercial maturity was judged visually on the basis of external appearance, including de-greening of fruit skin and slightly pink coloration at the fruit apex. Fruit observation was performed at an interval of several days. To confirm maturation differences not only visually but also from a physiological perspective, harvested fruit were measured for acoustic vibrations (f2 and f3 values and f3/f2 ratio) and ripening traits (ethylene production and flesh penetration force), as described previously (Kawai et al., 2018; Nakano et al., 2018).
In 2020, individual fruit grown at a near-canopy position in each pair was separately harvested at commercial maturity regardless of the maturation of its counterpart, to evaluate differences in harvest date between normal and split-pit fruit in each pair. Fruit observation was performed every two days, and this interval was shorter than in 2018 and 2019. Average harvest dates of normal and split-pit fruit were calculated as the days from July 1. Acoustic vibration, ethylene production, and flesh firmness of harvested fruit were measured as described for fruit in 2018 and 2019 to confirm that fruit were harvested at approximately the same maturity stage.
Evaluation of split pit at harvest and accuracy of split-pit fruit detection by acoustic vibration method
Actual split-pit occurrence in harvested fruit was evaluated after measurement of flesh firmness. First, the pit of the individual fruit was exposed along its suture line and pit splitting was examined visually. The remaining split-pit fruit, which could not be assessed by appearance, were identified after sticking a knife into the suture line of the pit. Fruit whose pit was easily cut into half by the knife were categorized as split-pit fruit, whereas those with pit that could not be cut by the knife, even after repeated thrusts, were categorized as normal fruit. Based on these criteria, split-pit fruit detection rate and normal fruit misdetection rate were expressed as the percentage of each predicted number relative to the actual number of split-pit fruit and normal fruit at harvest, respectively.
Measurement of ethylene production and flesh firmness of immature fruit and ethylene production from seeds
In 2019 and 2020, differences in ethylene production and flesh penetration force of immature fruit and ethylene production from seeds were compared between normal and split-pit fruit collected at the fruit development stage from late June to early July. We measured the acoustic vibration of fruit on June 24, July 1, and July 8 in 2019, and June 29 and July 7 in 2020, to estimate the pairs of normal and split-pit fruit grown at a near-canopy position, as described above. These measurement dates were in early to middle stage III of fruit growth of ‘Shimizu Hakuto’ peach. All fruit pairs were collected immediately after acoustic vibration measurement and their ethylene production, flesh penetration force, and actual split-pit occurrence on the measurement date were evaluated as described above. Seeds were excised from the pits of normal and split-pit fruit pairs and ethylene production was measured according to Kawai et al. (2018) with a slight modification, namely, a small 440 mL plastic container instead of a 1.3 L one was used for incubation at room temperature for 30 min. Measurement of ethylene production from seeds was also performed for mature fruit pairs collected in the harvest period in 2019 and 2020, which were the same fruit pairs as those used for comparison of maturation between normal and split-pit fruit, as described above.
Statistical analysis
Significant differences in the means of all measured traits were evaluated for pairs of normal and split-pit fruit based on the evaluation of actual split-pit occurrence at harvest or sampling point. The paired t-test was used for comparison of normal and split-pit fruit in each pair.
Results and Discussion
Split-pit fruit detection to estimate pairs of normal and split-pit fruit grown at a near-canopy position by acoustic vibration measurement
To prepare pairs of normal and split-pit fruit grown at a near-canopy position efficiently and compare their maturation precisely, acoustic vibrations of fruit on trees were measured at the fruit development stage (July 7 and 10 in 2018, July 9 and 10 in 2019, and July 6 and 8 in 2020). These measurement dates were in the middle of stage III of fruit growth of ‘Shimizu Hakuto’ peach, which were 9 to 13 days before the onset of harvest in this study (Fig. 1). Table 1 shows the number and percentage of fruit categorized as split-pit fruit when f3/f2 cut-off value was set at > 1.4. Among the 126, 120, and 88 fruit remaining on trees until harvest in 2018, 2019, and 2020, 85, 61, and 37 fruit respectively, were categorized as split-pit fruit by actual split-pit evaluation at harvest. Using the f3/f2 ratio of > 1.4 as the cut-off value between split-pit fruit and normal fruit, 88.2%, 91.8%, and 78.4% (75 of 85, 56 of 61, and 29 of 37) of split-pit fruit were successfully sorted, whereas 12.2%, 1.69%, and 0% (5 of 41, 1 of 59, and 0 of 51) of normal fruit were missorted as split-pit fruit. In addition, 14 of 85 (16.5%), 7 of 61 (11.4%), and 12 of 37 (32.4%) split-pit fruit in 2018, 2019, and 2020 could not be assessed by appearance but were identified after sticking a knife into the suture line of the pit. This type of split-pit fruit could be explained by the assumption that pit splitting in the early growing period was narrowed or closed up again by callus and/or resin in the flesh thereafter (Kawai et al., 2018). Some of the fruit with such a “repaired” split pit exhibited similar f3/f2 ratios to normal fruit (data not shown) and thus were difficult to distinguish from normal fruit on the basis of f3/f2 ratio. This seemed to lower the split-pit detection rate especially in 2020, when a high proportion of repaired split-pit fruit was detected in total split-pit fruit. In fact, the detection rate of split-pit fruit in 2018, 2019, and 2020 increased to 93.0%, 96.3%, and 96.0%, respectively, when repaired split-pit fruit were included in normal fruit. It is also possible that pit splitting had not occurred yet in several split-pit fruit on the day of acoustic vibration measurement, and occurred thereafter in the fruit. Nonetheless, measurement of f3/f2 values effectively distinguished normal fruit from split-pit fruit on trees with relatively high accuracy.

Table 2 shows the predicted and actual number of fruit pairs based on acoustic vibration measurement and split-pit evaluation at harvest, respectively. The predicted 30, 27, and 25 pairs of normal and split-pit fruit were obtained on the basis of f3/f2 values, whereas actual numbers of normal and split-pit fruit pairs were 27, 27, and 25 in 2018, 2019, and 2020, respectively. These results collectively showed that pairs of normal and split-pit fruit grown at a near-canopy position could be effectively predicted by acoustic vibration measurement on trees in all three years. The resulting 27, 27, and 25 pairs of normal and split-pit fruit were used for comparison of their maturation in the subsequent experiment.
Evaluation of differences in maturity date between normal and split-pit fruit
Figure 1 shows the number and percentage of pairs (2018, 2019) and fruit (2020) collected on each harvest date. In 2018 and 2019, fruit pairs with at least one split-pit fruit (i.e., split-pit × split-pit or normal × split-pit fruit pair) tended to be harvested at the earlier stage of the harvest period, whereas those with only normal fruit (i.e., normal × normal fruit pair) tended to be harvested at the later stage (Fig. 1A, B). In 2020, when each fruit was separately harvested at commercial maturity regardless of pairs, the same tendency of decreasing and increasing numbers of split-pit and normal fruit, respectively, was observed towards the end of the harvest period. Although not only split pit, but also bearing position of the fruit on trees probably affected this comparison, because the position of each fruit pair was randomly selected, these results showed that a higher proportion of split-pit fruit were harvested earlier than normal fruit at the entire-tree level.
To compare the maturation of normal fruit and split-pit fruit more precisely by minimizing the effect of canopy position, we investigated which of the two, normal and split-pit fruit, grown at a near-canopy position matured earlier in each pair. Figure 2 shows the number of pairs in which split-pit fruit were harvested earlier than their counterpart normal fruit. Among the 27, 27, and 25 pairs, split-pit fruit matured earlier in 25, 27, and 23 pairs (92.6%, 100%, and 92%) in 2018, 2019, and 2020, respectively. This result clearly showed that most split-pit fruit reached maturity earlier than normal fruit on trees even when comparing the fruit at a near-canopy position.
Differences in ripening traits and acoustic vibration profiles between normal and split-pit fruit at harvest
To confirm the observed maturation difference between normal and split-pit fruit from a physiological point of view, ripening traits (ethylene production and flesh penetration force) and acoustic vibrations (f2 and f3 values and f3/f2 ratio) of the harvested fruit were examined. In 2018 and 2019, when both fruit grown at a near-canopy position in each pair were harvested at the same time after either of them reached commercial maturity, the amount of ethylene produced was significantly larger in split-pit fruit than normal fruit, whereas flesh penetration force was significantly lower in split-pit fruit (Table 3). In general, higher ethylene production and lower penetration force indicate progress in fruit ripening in climacteric fruit. Furthermore, the f3 value was significantly lower in split-pit fruit than normal fruit in both years. As it was shown that f3 could be a good indicator to estimate flesh firmness and optimum harvest time of peach fruit minimally influenced by split-pit occurrence (Kawai et al., 2018), the lower f3 value also suggested early maturation of split-pit fruit. These results collectively indicated that the early maturation of split-pit fruit characterized by visual observation of fruit appearance on trees was also supported by other ripening-related traits, such as ethylene production, flesh firmness, and f3 value at harvest.

In 2020, on the other hand, no significant differences were observed for ethylene production, flesh firmness, and f3 value (Table 3). Considering that each fruit grown at a near-canopy position in each pair was separately harvested at commercial maturity in 2020 to estimate the difference in harvest date between normal and split-pit fruit, these results suggested that each fruit was properly harvested at approximately the same maturity stage. Under this experimental condition, the average harvest date of normal and split-pit fruit in 25 pairs was calculated as the days from July 1. It was shown that normal and split-pit fruit were harvested 26.12 ± 0.66 and 23.08 ± 0.78 days after July 1 on average, respectively. The biggest difference of ten days was observed for 2 pairs among the 23 pairs in which split-pit fruit were harvested earlier than their counterpart normal fruit, whereas there was only a 2-day difference for two pairs in which normal fruit were harvested earlier than their counterpart split-pit fruit. As a result, a significant difference in harvest date of approximately three days (P < 0.01 by paired t-test) was observed between normal and split-pit fruit, at least under the experimental conditions in 2020.
Unlike the f3 value, the f2 value was consistently lower and the f3/f2 value was consistently higher in split-pit fruit than normal fruit in all three years, irrespective of the difference in harvest procedure of fruit pairs between 2018–2019 and 2020 (Table 3). These results agreed with the fact that f2 and f3/f2 values were more strongly influenced by the split-pit occurrence than f3, as reported in previous studies (Kawai et al., 2018; Nakano et al., 2018).
Differences in ethylene production, flesh firmness, and ethylene production from seeds between normal and split-pit fruit from fruit development stage to harvest period
To gain an insight into the physiological mechanism of early maturation in split-pit fruit, we investigated ethylene production and flesh firmness of immature fruit and ethylene production from seeds at the fruit development stage. Figure 3 shows ethylene production from seeds excised from normal and split-pit fruit pairs collected on each sampling date from late June to early July (early to middle stage III of fruit growth) as well as in the harvest period in 2019 and 2020. Normal and split-pit fruit pairs grown at a near-canopy position were predicted by acoustic vibration measurement, and the tested fruit pairs were collected immediately after measurement for actual split-pit evaluation. The resultant numbers of normal and split-fruit pairs obtained were as follows: n = 13, 21, and 19 pairs for June 24, July 1, and July 8 in 2019, and n = 13 and 13 for June 29 and July 7 in 2020, respectively (data not shown for detection rate of split-pit fruit by acoustic vibration measurement). Ethylene production from seeds was significantly higher in split-pit fruit than normal fruit in the fruit pairs collected on June 24 and July 1 in 2019 (Fig. 3A) and June 29 in 2020 (Fig. 3B). No significant difference was observed between normal and split-pit fruit on later sampling dates. The amount of ethylene produced gradually decreased as the fruit (or seeds) matured, and no ethylene production from seeds was detected in the fruit collected during the harvest period from late July to early August for both normal and split-pit fruit. These results suggested that the high ethylene production from seeds at the fruit development stage that lasted until early July could be a possible factor that promotes maturation in split-pit fruit.

Differing from seeds, little ethylene production was detected from either immature normal or split-pit fruit in both 2019 and 2020, and no significant difference was observed between them except for June 29 in 2020 (Fig. S1). Correspondingly, no significant differences were observed for the flesh penetration force between normal and split-pit fruit at the fruit development stage (Fig. S2). These results collectively suggested that there could be a gap between the timing of ethylene production from seeds in immature split-pit fruit and the timing when its maturation-promoting effect was observed as the early initiation of fruit ripening on trees with the burst of ethylene production and/or reduction in flesh firmness, as observed in mature fruit collected during the harvest period.
Effect of split pit on the maturation of peach fruit on trees in relation to practical cultivation management to reduce problems caused by split pit
Although the early maturation of split-pit fruit on trees is widely known among peach growers, experimental demonstration and an understanding of the underlying physiological mechanism are lacking. In our previous study, where we used the acoustic vibration method for the same plant materials, it was shown that the method could be effectively used to detect split pit in unpicked peach fruit, and to determine accurately the timing of split-pit occurrence on trees (Kawai et al., 2018). In this study, the acoustic vibration method enabled us to estimate normal and split-pit fruit pairs grown at a near-canopy position quite efficiently and to evaluate the difference of their maturation on trees while minimizing the effect of canopy position (Tables 1 and 2). A three-year comparison of 79 pairs of normal and split-pit fruit revealed that most split-pit fruit matured earlier than normal fruit on trees in terms of external appearance, physiological properties, and acoustic vibration profiles (Figs. 1 and 2; Table 3). In general, peach fruit grown at a higher position on the tree tend to mature earlier regardless of split-pit occurrence. At the same time, pit splitting frequently occurs in fruit grown at a high position on the tree. Therefore, the combined effects of these two phenomena (i.e., split pit and canopy position) may be involved in the early maturation of split-pit fruit in terms of entire-tree level. It was experimentally shown that split-pit fruit itself advances fruit maturation regardless of canopy position. Approximately three days difference in harvest date was observed between normal and split-pit fruit, at least under the experimental conditions in 2020. Considering that the busiest time for ‘Shimizu Hakuto’ peach harvest is concentrated over a period of 7–10 days from late July to early August in the Okayama area, the observed difference in harvest date between normal and split-pit fruit is not negligible. It should be also noted that fruit (or fruit pairs) that dropped before harvest were excluded from the results of this study. As most of such fruit falling during the preharvest stage were split-pit fruit (data not shown), the actual gap in harvest date between normal and split-pit fruit would further widen if these preharvest fruit drops were taken into consideration. As the difference in harvest date was calculated on the basis of only a single year’s investigation of 25 pairs of normal and split-pit fruit in 2020, continuous research on the maturation difference using a larger number of normal and split-pit fruit pairs is essential to evaluate the difference in harvest date more accurately.
To gain an insight into the physiological mechanism underlying early maturation in split-pit fruit, we examined ethylene production and flesh firmness, and ethylene production from seeds in immature normal and split-pit fruit in 2019 and 2020. Acoustic vibration measurement enabled effective comparison of normal and split-pit fruit grown at a near-canopy position. It was shown that ethylene production from seeds was significantly higher in immature split-pit fruit than normal fruit until early July (Fig. 3). Regarding ethylene production from peach seeds, Jerie and Chalmers (1976) and Mizutani et al. (1998) reported that peach seeds produced large amounts of ethylene immediately after they were taken out from pits, and their ethylene producing ability remained high during the early stage of seed development but gradually decreased as they matured. Our results agree with the previous studies. Mizutani et al. (1998) further demonstrated that the high CO2 and low O2 conditions inside the seed suppressed 1-aminocyclopropane-1-carboxylate (ACC) biosynthesis and inhibited ethylene production, and both ACC biosynthesis and ethylene production were enhanced under the opposite conditions of high O2 and low CO2. It was also reported in other studies that oxygen intensified ethylene-induced metabolic and cellular processes in other crops, such as potato and tomato (Chin and Frenkel, 1976, 1977; Frenkel and Garrison, 1976). These findings were discussed in relation to pit splitting and ripening of peach fruit, suggesting that ambient air entering the pit by split-pit occurrence triggers a burst of ethylene production from seeds, which results in premature ripening and preharvest fruit drop in split-pit peach (Mizutani et al., 1998). Although Mizutani et al. (1998) did not evaluate the difference in ethylene production from seeds between normal and split-pit fruit, the comparison results of normal and split-pit fruit in this study, in combination with these previous reports, collectively suggest that higher ethylene production from seeds until early July could affect maturation of split-pit fruit from inside the fruit, which may be a possible factor causing early maturation in split-pit fruit. Interestingly, little ethylene production was detected from the same fruit pairs from which seed was excised, and no significant difference in flesh firmness was observed between immature normal and split-pit fruit, despite a larger amount of ethylene produced from seeds in split-pit fruit (Figs. S1 and S2). Differences in ethylene production and flesh firmness of fruit were observed only when the fruit reached harvest maturity. Such a time lag between ethylene exposure and advanced onset of fruit ripening was also observed in other climacteric fruit, such as tomato (Yang, 1987) and banana (Burg and Burg, 1962); exogenous ethylene treatment of immature fruit failed to initiate immediate ripening but significantly advanced the timing of ripening subsequently. Yang (1987) reported that earlier ripening was initiated in immature tomato fruit treated with larger amounts of ethylene. As an explanation for these observations, Kevany et al. (2007) demonstrated that treatment of immature tomato fruit with ethylene led to early ripening and reduction in the levels of two ethylene receptor proteins (LeETR4 and LeETR6) that negatively regulate ethylene signal transduction and resultant ethylene responses, which is consistent with a model in which the onset of fruit ripening is modulated by receptor protein levels that are dependent on the cumulative amount of ethylene exposure. Although differences in the levels of ethylene receptor proteins between immature normal and split-pit fruit and the direct effects of ethylene from seeds on the protein levels were not evaluated in this study, the same mechanism may be involved in early maturation of split-pit fruit on trees. Further studies are needed to fully understand the whole mechanism of the maturation-promoting effect of split pit.
For practical cultivation management in relation to fruit maturation, split-pit fruit have a higher risk of premature ripening and preharvest fruit drop, which are serious problems for peach growers. Premature ripening before the completion of normal sugar accumulation could also lead to inferior fruit quality, such as lower SSC. Even if these split-pit fruit remain on trees until harvest, it would be difficult to keep fruit quality for a long time after harvest in the case of fruit with advanced ripening and/or originally inferior quality. As most split-pit fruit are harvested in the earlier stage of the harvest season as shown in this study and some split-pit fruit are difficult to sort from appearance, careful attention should be directed to the contamination of split-pit fruit especially when peach fruit are sold as gifts or exported to foreign countries at a high price. The acoustic vibration method could be used as an effective tool to remove these undesired split-pit fruit from trees by fruit thinning, as discussed in a previous report (Kawai et al., 2018). For this purpose, we are now developing and testing new simplified acoustic vibration devices that can detect split pit much more easily and rapidly in commercial orchards. Continuous research, in combination with the development and testing of such new devices, will help us deepen our understanding of the factors that induce split pit and the effect of split pit on fruit maturation on trees, as well as develop effective methods to solve problems caused by split pit before harvest.
Literature Cited
- Abbaszadeh, R., A. Moosavian, A. Rajabipour and G. Najafi. 2015. An intelligent procedure for watermelon ripeness detection based on vibration signals. J. Food Sci. Technol. 52: 1075–1081.
- Ali, M. M., N. Hashim, S. K. Bejo and R. Shamsudin. 2017. Rapid and nondestructive techniques for internal and external quality evaluation of watermelons: A review. Sci. Hortic. 225: 689–699.
- Burg, S. P. and E. A. Burg. 1962. Role of ethylene in fruit ripening. Plant Physiol. 37: 179–189.
- Burrell, R. E. and G. L. Reighard. 2017. Late dormant pruning affects split pit incidence in peach. Acta Hortic. 1177: 157–164.
- Chin, C. K. and C. Frenkel. 1976. Influence of ethylene and oxygen on respiration and peroxide formation in potato tubers. Nature 264: 60.
- Chin, C. K. and C. Frenkel. 1977. Upsurge in respiration and peroxide formation in potato tubers as influenced by ethylene, propylene, and cyanide. Plant Physiol. 59: 515–518.
- Claypool, L. L., K. Uriu and P. F. Lasker. 1972. Split-pit of ‘Dixon’ cling peaches in relation to cultural factors. J. Amer. Soc. Hort. Sci. 97: 181–185.
- Cooke, J. R. 1972. An interpretation of the resonant behavior of intact fruits and vegetables. Trans. ASAE 15: 1075–1080.
- Crisosto, C. H., R. S. Johnson, K. R. Day and T. DeJong. 1997. Orchard factors affecting postharvest stone fruit quality. HortScience 32: 820–823.
- Davis, L. D. 1941. Split-pit of peaches: Estimation of time when splitting occurs. Proc. Amer. Soc. Hort. Sci. 39: 183–189.
- Drake, B. 1962. Automatic recording of vibrational properties of foodstuffs. J. Food Sci. 27: 182–188.
- Drogoudi, P. D., C. G. Tsipouridis and G. Pantelidis. 2009. Effects of crop load and time of thinning on the incidence of split pits, fruit yield, fruit quality, and leaf mineral contents in ‘Andross’ peach. J. Hort. Sci. Biotech. 84: 505–509.
- Evert, D. L., T. P. Gaines and B. G. Mullinix. 1988. Effects of split-pit on elemental concentrations of peach fruit during pit hardening. Sci. Hortic. 34: 55–65.
- Frenkel, C. and S. A. Garrison. 1976. Initiation of lycopene synthesis in the tomato mutant rin as influenced by oxygen and ethylene interactions. HortScience 11: 20–21.
- Fukuda, F., R. Yoshimura, H. Matsuoka and N. Kubota. 2006. Relationship between fruit enlargement and seed development with respect to physiological fruit drop in ‘Shimizu Hakuto’ peach. J. Japan. Soc. Hort. Sci. 75: 213–218.
- Jerie, P. H. and D. J. Chalmers. 1976. Some characteristics of ethylene production in peach (Prunus persica L.) seeds. Planta 132: 13–17.
- Kadowaki, M., S. Nagashima, H. Akimoto and N. Sakurai. 2012. Detection of core rot symptom of Japanese pear (Pyrus pyrifolia cv. Kosui) by a nondestructive resonant method. J. Japan. Soc. Hort. Sci. 81: 327–331.
- Kawai, T., F. Matsumori, H. Akimoto, N. Sakurai, K. Hirano, R. Nakano and F. Fukuda. 2018. Nondestructive detection of split-pit peach fruit on trees with an acoustic vibration method. Hort. J. 87: 499–507.
- Kevany, B. M., D. M. Tieman, M. G. Taylor, V. D. Cin and H. J. Klee. 2007. Ethylene receptor degradation controls the timing of ripening in tomato fruit. Plant J. 51: 458–467.
- Kritzinger, I., E. Lӧtze and M. Jooste. 2017. Stone hardening and broken stones in Japanese plums (Prunus salicina Lindl.) evaluated by means of computed tomography scans. Sci. Hortic. 221: 1–9.
- Kubota, N., N. Nishiyama and K. Shimamura. 1993. Effects of girdling lateral bearing branches on astringency and phenolic contents of peach fruits. J. Japan. Soc. Hort. Sci. 62: 69–73 (In Japanese with English abstract).
- Miki, T. 1932. Studies on the development of peach fruits with special reference to the cause of their split-pit. Bull. Chiba Coll. Hort. 1: 1–118.
- Mizutani, F., G. A. Rabbany and H. Akiyoshi. 1998. Ethylene biosynthesis in peach seeds and its suppression in situ. J. Japan. Soc. Hort. Sci. 67: 147–152.
- Murayama, H., I. Konno, S. Terasaki, R. Yamamoto and N. Sakurai. 2006. Nondestructive method for measuring fruit ripening of ‘La France’ pears using a laser Doppler vibrometer. J. Japan. Soc. Hort. Sci. 75: 79–84.
- Nakano, M. and M. Nakamura. 2001. Cracking and mechanical properties of the stone in peach cultivars after severe thinning. Acta Hortic. 592: 531–536.
- Nakano, R., H. Akimoto, F. Fukuda, T. Kawai, K. Ushijima, Y. Fukamatsu, Y. Kubo, Y. Fujii, K. Hirano, K. Morinaga and N. Sakurai. 2018. Nondestructive detection of split pit in peaches using an acoustic vibration method. Hort. J. 87: 281–287.
- Oveisi, Z., S. Minaei, S. Rafiee, A. Eyvani and A. Borghei. 2014. Application of vibration response technique for the firmness evaluation of pear fruit during storage. J. Food Sci. Technol. 51: 3261–3268.
- Ragland, C. H. 1934. The development of the peach fruit, with special reference to split-pit and gumming. Proc. Amer. Soc. Hort. Sci. 31: 1–21.
- Takahashi, M., M. Taniwaki, N. Sakurai, T. Ueno and H. Yakushiji. 2010. Changes in berry firmness of various grape cultivars on vines measured by nondestructive method before and after veraison. J. Japan. Soc. Hort. Sci. 79: 377–383.
- Taniwaki, M. and N. Sakurai. 2010. Evaluation of the internal quality of agricultural products using acoustic vibration techniques. J. Japan. Soc. Hort. Sci. 79: 113–128.
- Taniwaki, M., T. Hanada and N. Sakurai. 2009a. Postharvest quality evaluation of “Fuyu” and “Taishuu” persimmons using a nondestructive vibrational method and an acoustic vibration technique. Postharvest Biol. Technol. 51: 80–85.
- Taniwaki, M., T. Hanada, M. Tohro and N. Sakurai. 2009b. Nondestructive determination of the optimum eating ripeness of pears and their texture measurements using acoustical vibration techniques. Postharvest Biol. Technol. 51: 305–310.
- Taniwaki, M., M. Takahashi and N. Sakurai. 2009c. Determination of optimum ripeness for edibility of postharvest melons using nondestructive vibration. Food Res. Int. 42: 137–141.
- Taniwaki, M., M. Tohro and N. Sakurai. 2010. Measurement of ripening speed and determination of the optimum ripeness of melons by a nondestructive acoustic vibration method. Postharvest Biol. Technol. 56: 101–103.
- Terasaki, S., N. Sakurai, S. Kuroki, R. Yamamoto and D. J. Nevins. 2013. A new descriptive method for fruit firmness changes with various softening patterns of kiwifruit. Postharvest Biol. Technol. 62: 85–90.
- Terasaki, S., N. Sakurai, J. Zebrowski, H. Murayama, R. Yamamoto and D. J. Nevins. 2006. Laser Doppler vibrometer analysis of changes in elastic properties of ripening ‘La France’ pears after postharvest storage. Postharvest Biol. Technol. 42: 198–207.
- Woodbridge, C. G. 1978. Split-pit in peaches and nutrient levels. J. Amer. Soc. Hort. Sci. 103: 278–280.
- Yang, S. F. 1987. The role of ethylene and ethylene synthesis in fruit ripening. p. 156–165. In: W. Thompson, E. Nothagel, R. Huffake (eds.). Plant Senescence: Its Biochemistry and Physiology. The american society of plant physiologists, Rockville, MD.


