Abstract
The effects of pulse treatments with 0.2 mM silver thiosulfate complex (STS), 5 and 10% sucrose, and 0.2% calcium chloride (CaCl2) and their combinations on the vase life and soluble carbohydrate and aurone levels in cut snapdragon (Antirrhinum majus) flowers were investigated. In the pulse treatment, the cut flowers were treated with these chemical solutions for 24 h before being transferred to distilled water. The uptake of solution for pulse treatments was decreased in the treatments with STS, sucrose, and CaCl2 and the combined STS and sucrose treatment further suppressed uptake. The sucrose treatment significantly increased the number of open flowers and extended the vase life, with no marked differences in these effects between the 5% and 10% sucrose solutions. The STS treatment slightly promoted flower opening but did not significantly extend vase life. The CaCl2 treatment did not affect flower opening or vase life. The relative fresh weight (RFW) and water uptake of cut flowers were increased in the sucrose treatments and were only slightly affected by the STS and CaCl2 treatments. The petal aurone levels were significantly higher in the sucrose treatments, particularly at 10% sucrose. The glucose, fructose, and sucrose petal contents were generally higher in the sucrose treatments than in the treatments that did not include sucrose. The ethylene production of flowers was relatively low at harvest and somewhat increased during senescence. Therefore, the infectiveness of STS on vase life may be due to low ethylene production. We conclude that pulse treatments with solutions containing sucrose increase the sugar contents and aurone levels in petals, as well as improve the vase life of cut snapdragons.
Introduction
Snapdragons (Antirrhinum majus) are popular ornamental plants with a wide range of petal colors and a good fragrance, making them an ideal choice for cut flower arrangements. Snapdragons have indeterminate inflorescences with many flowers, including buds. The vase life of cut snapdragon flowers is relatively short, terminating due to petal abscission, incomplete flower opening, suppression of pigmentation, and bending of flower spikes (Ҫelikel et al., 2010; Ichimura et al., 2008; Nowak, 1981).
Snapdragon petals are abscised when exposed to ethylene (Ichimura et al., 2008; Woltering and van Doorn, 1988), indicating that snapdragon flowers are sensitive to ethylene. To extend the vase life of cut flowers, treatment with chemicals for a short period after harvest is called pulse treatment (Halevy and Mayak, 1981). In many ethylene-sensitive cut flowers, such as carnation (Veen, 1979), Delphinium (Ichimura et al., 2009b), and sweet pea (Mor et al., 1984), their vase life can be extended by pulse treatment with silver thiosulfate complex (STS), an inhibitor of ethylene action. However, when applied to snapdragons, pulse treatment with STS only slightly extends the vase life of cut snapdragons (Ichimura et al., 2008; Nowak, 1981). Although Campanula medium (Kato et al., 2002) and Japanese morning glory flowers (Yamada et al., 2006) are sensitive to ethylene, treatments with ethylene inhibitors do not delay their petal senescence. Some reports attribute the ineffectiveness of ethylene inhibitors to the absence of climacteric-like increases in ethylene production (Kato et al., 2002; Yamada et al., 2006).
Treatments with sugars, such as glucose and sucrose extend the vase life of many cut flowers, including rose (Ichimura et al., 1999), carnation (Pun et al., 2016), and Eustoma grandiflorum (Shimizu and Ichimura, 2005). In particular, treatment with sucrose promotes flower opening and extends the vase life of cut flowers with many floral buds, such as hybrid Limonium (Doi and Reid, 1995; Shimamura et al., 1997), freesia (Woodson, 1987), and sandersonia (Eason et al., 1997).
Snapdragon flower spikes have many floral buds and continuous treatment with sucrose markedly extends their vase life (Ichimura and Hisamatsu, 1999; Larsen and Scholes, 1966; Marousky and Raulston, 1970). Although pulse treatment with STS slightly extends the vase life of cut snapdragons, combining the STS treatment with sucrose significantly extends vase life (Ichimura et al., 2008). Aurone is a pigment responsible for the yellow color of snapdragons (Ichimura et al., 2008; Nakayama et al., 2000). Pulse treatment with sucrose increases aurone levels in the petals of cut snapdragon, but STS treatment does not (Ichimura et al., 2008).
Snapdragon flowers wilt from base of the spikes. In gladiolus and daylily spikes, which have many open flowers and floral buds, sugars in senescing petals are transported to the buds, contributing to flower opening (Bieleski, 1995; Yamane et al., 1995). Thus, sugars may be transported from basal flowers to upper flowers in snapdragon. Treatment with sucrose increases the sugar content of floral buds in some cut flowers, including hybrid Limonium (Shimamura et al., 1997) and sweet pea (Ichimura and Suto, 1999), as well as snapdragon (Ichimura et al., 2008). However, to our knowledge, it has not yet been investigated whether sucrose treatment increases soluble carbohydrate contents of opened flowers in spikes. Therefore, determination of soluble carbohydrate contents of flowers at basal and upper positions would contribute to understanding the role of endogenous and exogenous soluble carbohydrates in cut snapdragon flowers.
Calcium plays a key role in cross-linking acidic pectin residues in the cell wall (Hepler, 2005). Gerasopoulos and Chebli (1999) reported that treatment with calcium chloride (CaCl2) suppresses stem bending in cut gerbera flowers. The same treatment was effective at suppressing the bending of cut snapdragon spikes (Fukai and Uehara, 2006). However, the effect of a combined treatment of STS, CaCl2, and sucrose on the vase life of cut snapdragons has not yet been investigated.
In pulse treatments, the cut flowers must uptake the solution in order for the effects to be observed and extend their vase life. However, pulse treatment with STS reduces the uptake of vase solution in cut Gypsophila (Doi et al., 1999). Similarly, in cut roses, the uptake of vase solution was decreased with the addition of sucrose (Ichimura and Shimizu-Yumoto, 2007). Thus, the uptake of solution in a pulse treatment may be reduced further in a combined treatment with STS and sucrose. However, the effect of a combined treatment of STS, CaCl2, and sucrose on the uptake of solution for pulse treatment has not yet been investigated.
Ichimura et al. (2008) previously reported that pulse treatment with sucrose and STS extended the vase life of cut snapdragons, but the cut flowers used in the study were short. In addition, the temperature for the treatment was 23°C, which is higher than typical pulse treatment. In the present study, we examined the effects of pulse treatment with STS, sucrose, and CaCl2 on the uptake of pulse solution, vase life, and petal aurone and soluble carbohydrate levels in cut snapdragons. It was expected that the combined treatment with STS, sucrose, and CaCl2 would delay the senescence of open flowers, promote flower opening and pigmentation and suppress bending of spikes in cut snapdragons. We used cut flowers with relatively long stems and a treatment temperature of 15°C. These experimental conditions are similar to practical conditions for pulse treatment (personal observation). Therefore, our results can be applied to practical treatment. In addition, we investigated the ethylene production of flowers during senescence.
Materials and Methods
Plant materials, chemical treatment and evaluation of vase life
Snapdragon ‘Yellow Butterfly’ plants were grown in a greenhouse under natural daylight conditions with a minimum temperature of 15°C and a maximum temperature of 25°C. When four or five flowers opened, flower spikes were cut from the plants and immediately transported to the laboratory. Flower spikes were re-cut to 55 cm and treated with STS at two levels (0 and 0.2 mM), sucrose at three levels (0, 5 and 10%) and CaCl2 at two levels (0 and 0.2%) or their combinations. The treatment compounds were dissolved in distilled water. The flower spikes were held at 15°C and 70% relative humidity in the dark for 24 h. After the pulse treatment, uptake of the solutions was measured. The flower spikes were then cut to 50 cm and placed in distilled water. The spikes were held at 23°C and 70% relative humidity, under 10 μmol·m−2·s−1 light with a 12-h photoperiod. The number of open and wilted flowers, fresh weight and water uptake of the cut spikes were measured daily until all flowers had wilted. Open flowers were defined as having unfolded petals on stems that had not bent. The vase life of the whole spike was determined as the time from the end of pulse treatment to when the numbers of open flowers became lower than the number of open flowers at harvest or when the stems became bent. Relative fresh weight (RFW) was calculated by dividing the fresh weight of each day by the fresh weight at the start of vase life.
In a separate experiment, flowers at the 1st and 2nd, or 10th and 11th position from the base were collected 5 days after the start of vase life and aurone levels and soluble carbohydrate contents were measured.
Determination of aurones
Pieces of the upper petal lobes (200 mg) were immersed in 5 mL of methanol and kept at 4°C in the dark for 7 days. The methanol was decanted and the petal pieces were washed twice with 2.5 mL of methanol. All washes were combined for a final volume of 10 mL and the absorbance was measured using a spectrophotometer at 405 nm.
Determination of soluble carbohydrates
Soluble carbohydrates were extracted following the methods of Norikoshi et al. (2008). In brief, the upper petal lobes were immersed in 80% ethanol at 70°C for 20 min. After cooling, galactose was added to the sample as an internal standard. The immersion was filtered and dried in vacuo below 50°C. The concentrate was dissolved in the minimum volume of water. The eluate was then separated by high-performance liquid chromatography (HPLC) with a refractive index detector on a Shodex SUGAR SP0810 column (Showa Denko, Tokyo, Japan) at 80°C and eluted with water at a flow rate of 0.8 mL·min−1. The identity of each peak was confirmed with authentic carbohydrates. The peak areas were determined by an integrator and the amounts of each carbohydrate in each sample were calculated as described by Norikoshi et al. (2008).
Measurement of ethylene production
On the day of anthesis, the flowers were cut from the plants, placed in distilled water and maintained at 23°C, 70% relative humidity, under 10 μmol·m−2·s−1 light with a 12 h photoperiod. The individual flowers were enclosed in a 30-mL Erlenmeyer flask (38.1 mL volume) for 1 h at 23°C. A 1 mL gas sample was withdrawn with a hypodermic syringe and injected into a gas chromatograph (GC-14B; Shimadzu, Kyoto, Japan) equipped with an alumina column and flame ionization detector. The GC conditions were as follows: injection temperature, 100°C; column temperature, 80°C; and carrier gas flow rate: 40 mL·m−1.
Statistical analysis
Analysis of variance and Tukey-Kramer’s multiple range test were performed using SigmaPlot software (v.12.5; Systat Software, San Jose, CA, USA).
Results
Effects of STS, sucrose, and CaCl2 on uptake of solutions for pulse treatment
In a preliminary experiment, leaf injury was observed in cut snapdragons treated with CaCl2 at 0.4% or higher concentrations. Thus, CaCl2 concentration at 0.2% was used for subsequent experiments.
The ANOVA revealed that STS, sucrose and CaCl2 treatments significantly reduced solution uptake by the flower spikes (Table 1). The STS and sucrose treatments reduced solution uptake more than the CaCl2 treatment. The reduction in solution uptake in the sucrose treatment tended to be greater at 10% than at 5%. The combined STS and sucrose treatments reduced the solution uptake more than either compound alone.
Effects of STS, sucrose, and CaCl2 on flower opening, vase life, RFW, and water uptake
In the distilled water control, the average number of open flowers increased up to 7.2 at day 2 and decreased thereafter. The combined treatment with STS and CaCl2 delayed the decrease in the open flower number. In the 5% sucrose treatment, the number of open flowers peaked at day 5 at 10.7. The number of open flowers was slightly suppressed when sucrose was combined with STS and/or CaCl2. Treatment with 10% sucrose increased the open flower number to 10.8 on day 5, and this number was lower with the combination of STS and/or CaCl2.
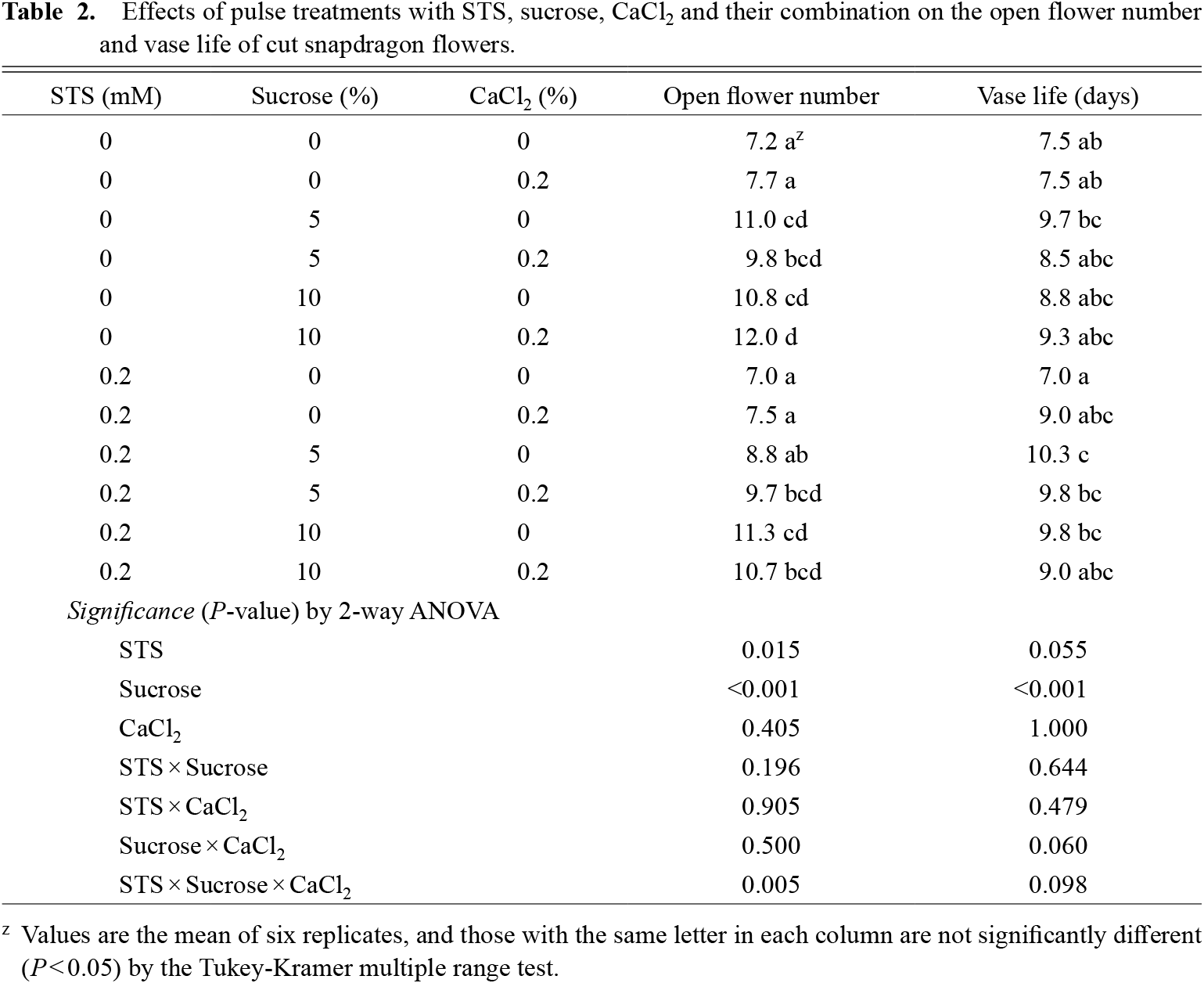
ANOVA revealed that sucrose treatments significantly increased the number of open flowers in comparison to the control (Table 2; Fig. 1). STS treatment only slightly increased the open flower number and the CaCl2 treatment had no effect.
The RFW of flowers in the control increased by 14% from day 0 to day 3 and decreased thereafter (Fig. 2). The decrease in the RFW of flowers was slightly suppressed in the CaCl2 treatments. The RFW of flowers treated with STS, CaCl2, and a combination of them showed similar trends to the control. Treatment with 5% sucrose increased the RFW by 22% from day 0 to day 4. There was a slight difference in RFW between the 5% sucrose and 5% sucrose plus CaCl2 treatments. Treatment with 5% sucrose in combination with STS suppressed the increases in RFW irrespective of the CaCl2 treatment. Treatment with 10% sucrose increased RFW by 25% from day 0 to day 6 and this increase was slightly lower in the treatments with a combination of STS and/or CaCl2.

ANOVA revealed that the sucrose treatment significantly extended the vase life of flower spikes, while the STS and CaCl2 treatments had no significant effect (Table 2). The 10% sucrose treatment was more effective in extending vase life than the 5% sucrose treatment.
The water uptake by the cut flowers decreased with time in all treatments (Fig. 3A–C). Treatment with CaCl2 did not affect water uptake (Fig. 3A). Treatment with STS suppressed water uptake and this suppression was greater in the combination treatment with CaCl2. Treatment with 5% and 10% sucrose increased the water uptake until day 3 or 4 (Fig. 3B, C). Water uptake tended to be suppressed in treatments with 5% sucrose in combination with STS and/or CaCl2 (Fig. 3B). Water uptake was slightly affected in treatments with 10% sucrose in combination with STS and/or CaCl2 (Fig. 3C).
Effect of STS and sucrose on aurone levels of petals
The aurone levels and carbohydrate contents in petals were investigated using cut spikes treated with STS, sucrose or their combinations. Those treated with CaCl2 were excluded because the treatment did not extend vase life.
There were no significant differences in the aurone levels of the 1st and 2nd flowers between the treatments (Table 3). The treatments with solutions including sucrose increased aurone levels in the petals of 10th and 11th flowers and 10% sucrose was more effective than 5% sucrose. The treatments with solutions including STS slightly affected aurone levels in the petals of these flowers. The pigmentation of the petals was improved in treatments with solutions including sucrose (data not shown).
Effect of STS and sucrose on soluble carbohydrate contents in petals
Glucose, fructose, sucrose, and mannitol were detected and the fructose content was the highest of the soluble carbohydrates in the petals (Table 4). ANOVA revealed that the fructose content was higher in 10th and 11th flowers than in 1st and 2nd flowers, but the sucrose content was higher in 1st and 2nd flowers than in 10th and 11th flowers. Treatment with 5% and 10% sucrose similarly increased the glucose, fructose and sucrose contents in the petals. The STS treatment slightly affected these sugar contents. The mannitol content was slightly affected by sucrose and STS treatments.
Ethylene production
At harvest, the ethylene production of the flowers was 1.2 nL·g−1FW·h−1. The ethylene production of the flowers remained almost constant for the first 8 days and then slightly increased on day 10 (Fig. 4).
Discussion
In this study, we investigated the effects of pulse treatments with STS, sucrose, and CaCl2 and their combinations on the uptake of pulsed solution, vase life, soluble carbohydrate contents, and aurone levels of cut snapdragon flowers. The sucrose treatment improved vase life, but the STS and CaCl2 treatments did not. The sugar content in petals was higher in treatments with sucrose than those without.
Treatments with STS and sucrose reduced the uptake of vase solution (Doi et al., 1999; Ichimura and Shimizu-Yumoto, 2007). In the present study, uptake of solution was reduced by STS, sucrose, and CaCl2 and the interaction between STS and sucrose was significant (Table 1). To our knowledge, this is the first report to demonstrate the interaction between STS and sucrose in solution uptake. For some cut flowers, including Eustoma (Shimizu and Ichimura, 2005), Gypsophila (Downs et al., 1988) and sweet pea (Mor et al., 1984), pulse treatment with STS combined with sucrose is effective in extending vase life. In these flowers, STS and sucrose may not be taken up in sufficient amounts to extend vase life. An extension of the treatment time may be more effective to improve the vase life of these flowers.
Fukai and Uehara (2006) reported that cut snapdragon spikes were prone to bending during vase life but treatment with 1% CaCl2 suppressed the bending. The effectiveness of CaCl2 to prevent spike bending could not be confirmed in the present study because the bending of control cut spikes was not observed. In preliminary experiments, we observed that the stems tended to bend in cut flowers with relatively thin stems (unpublished result). In the present study, we used cut snapdragons with relatively thick stems, so the lack of bending may have been due to variations in stem thickness and hardness. Moreover, the difference in the effectiveness of CaCl2 may be attributed to the use of different cultivars in the two studies. Although Fukai and Uehara (2006) did not mention chemical damage to the leaves in CaCl2 treatment, we observed leaf damage in the spikes with a CaCl2 concentration as low as 0.4% (data not shown). Therefore, the availability of CaCl2 during the vase life of cut snapdragons appears to be limited.
In previous studies, pulse treatment with sucrose at 7.5% was the most effective in extending vase life when sucrose concentrations up to 7.5% were investigated (Nowak, 1981). Ichimura et al. (2008) reported that 10% sucrose was the most effective when sucrose concentrations up to 12.5% were investigated. We found that treatment with 5% and 10% sucrose extended the vase life and increased aurone levels (Table 3; Fig. 1), although 10% sucrose was slightly more effective in promoting flower opening than 5% sucrose (Table 2). Thus, treatment with 5% sucrose appears to be effective to improve the vase life in terms of practical application. In the present study, cut flower length was longer than that used in the previous study; the sugar content of snapdragon stems is considerable (Norikoshi et al., 2008). In addition, the vase life of untreated snapdragon flowers was much longer in the present study than in the previous study (Ichimura et al., 2008). Therefore, the effectiveness of 5% sucrose in the present study may be linked to the amounts of sugars contained in the stems.
Snapdragon flowers are sensitive to ethylene (Ichimura et al., 2008), and pulse treatment with STS is generally recommended to improve the vase life of flowers (Ichimura, 2020). However, we found that pulse treatment with STS did not significantly extend vase life (Table 2). Similar results have been reported by Nowak (1981) and Ichimura et al. (2008). Ethylene production of snapdragon flowers was relatively low throughout vase life and the typical climacteric-like increase in ethylene production was not observed (Fig. 4). Flowers of carnation (Wu et al., 1991), Delphinium (Ichimura et al., 2009b) and sweet pea (Kebenei et al., 2003; Mor et al., 1984) are highly sensitive to ethylene and showed typical climacteric increases in ethylene production during senescence. Pulse treatment with STS markedly extends the vase life of these cut flowers (Kuroshima et al., 2009; Mor et al., 1984; Veen, 1979). In cut Campanula medium (Kato et al., 2002) and Japanese morning glory (Yamada et al., 2006) flowers, which are also sensitive to ethylene, ethylene production does not increase during senescence and treatment with ethylene inhibitors does not delay petal senescence. Thus, the ineffectiveness of STS treatments on improving vase life was possibly due to the absence of a climacteric-like increase in ethylene production. Snapdragon flowers are sensitive to ethylene (Ichimura et al., 2008; Woltering and van Doorn, 1988) and STS treatment reduces sensitivity to ethylene (Veen, 1979). In fact, STS treatment suppresses petal senescence induced by exogenous ethylene in cut snapdragons (Serek et al., 1995). Thus, we can recommend pulse treatment of cut snapdragon flowers with STS even if the effectiveness is limited.
Glucose, fructose, sucrose, and mannitol are present in snapdragon petals (Ichimura and Hisamatsu, 1999). We found that sucrose treatments significantly increased the glucose and fructose content in the snapdragon petals. This suggests that the applied sucrose is metabolized to glucose and fructose, resulting in accumulation in the petals. In the control flowers, the fructose content in the petals of the 1st and 2nd positions was lower than that in the petals of the 10th and 11th position flowers (Table 4), suggesting that the sugars at the lower part of the spikes may be transported to those at the upper part. The glucose and fructose contents in the petals were significantly higher in the treatments with sucrose than in the treatment without sucrose, irrespective of flower positions. This indicates that sucrose treatment increases sugar contents of floral buds as well as open flowers. We observed a longer vase life of the 1st and 2nd flowers treated with sucrose compared to the flowers in the control (unpublished result). Sugars act as a substrate for respiration and osmotica, which effectively extends the vase life of flowers (Halevy and Mayak, 1979). Thus, the increase in sugar content in the petals that occurred with sucrose application may contribute to the extended the vase life of flowers at a lower position.
The pulse treatments with solutions that included sucrose increased the aurone levels in the 10th and 11th flowers, but the STS treatments did not (Table 3). This increase in aurone levels may be due to the supply of a substrate for aurone glycosylation. Moreover, the expression of PhCHS-A encoding chalcone synthase, a key enzyme involved in flavonoid biosynthesis, is increased by exogenous sucrose in petunia (Moalem-Beno et al., 1997; Tsukaya et al., 1991). In snapdragon petals, the expression of AmCHS1 increased and AmAS1 encoding aureusidin synthase decreased in response to sucrose treatment (Ichimura et al., 2009a). These findings suggest that increases in aurone levels may be involved in AmCHS1 gene expression.
Combined treatment with STS and sucrose is effective in extending the vase life of cut flowers that are sensitive to ethylene and have floral buds, including Delphinium (Kuroshima et al., 2009), Eustoma (Shimizu and Ichimura, 2005), Gypsophila (Downs et al., 1988), hybrid Limonium (Doi and Reid, 1995) and sweet pea (Ichimura and Hiraya, 1999). The present study revealed that the sucrose treatment extended vase life significantly more than STS treatment (Table 2). In contrast, STS extends vase life significantly more than sucrose in Eustoma (Shimizu and Ichimura, 2005). Further studies are required to determine whether STS or sucrose is more effective in extending the vase life of other cut flowers.
The RFW and water uptake of the cut spikes were increased by pulse treatments with solutions that included sucrose (Figs. 2 and 3). Similar findings were reported with sucrose pulse treatments in other cut flowers, including roses (Ichimura and Shimizu-Yumoto, 2007) and Eustoma (Shimizu-Yumoto and Ichimura, 2010). Sugars act as osmotica that facilitate water uptake in cut flowers (Yamada et al., 2009). The treatments with sucrose solutions increased the sugar content of petals (Table 4). In addition, sucrose treatment promotes water uptake in cut hybrid Limonium (Doi and Reid, 1995). Thus, sucrose treatments supply sugars that are required as osmotica for water uptake.
Mannitol content was not significantly affected by any treatments (Table 4). Similarly, the mannitol content was not markedly changed by continuous treatments with glucose and sucrose and was metabolized less than glucose and sucrose in snapdragons (Ichimura et al., 2016). These results suggest that mannitol has little effect on the vase life of cut snapdragons.
In conclusion, in the pulse treatments with STS, sucrose, and CaCl2 the uptake of solution by cut snapdragons decreased. The reduction of solution uptake was greater in the combined STS and sucrose treatment. Pulse treatments with 5% and 10% sucrose promoted flower opening and extended the vase life of cut snapdragon flowers. RFW and water uptake of the cut flowers were increased by the sucrose treatments. The sugar content and aurone levels in the petals were also higher in the flowers treated with sucrose. Pulse treatments with STS and CaCl2 did not significantly extend vase life. The ethylene production of the cut flowers increased slightly during senescence, suggesting that the low effectiveness of STS for improving vase life may be due to this low ethylene production.
Literature Cited
- Bieleski, B. L. 1995. Onset of phloem export from senescent petals of daylily. Plant Physiol. 109: 557–565.
- Ҫelikel, F. C., J. C. Cevallos and M. S. Reid. 2010. Temperature, ethylene and the postharvest performance of cut snapdragons (Antirrhinum majus). Sci. Hortic. 125: 429–433.
- Doi, M. and M. S. Reid. 1995. Sucrose improves the postharvest life of cut flowers of a hybrid Limonium. HortScience 30: 1058–1060.
- Doi, M., N. Nagai and H. Imanishi. 1999. Postharvest quality of cut Gypsophila paniculata L. as influenced by environmental conditions during transport. J. Japan. Soc. Hort. Sci. 68: 635–639 (In Japanese with English abstract).
- Downs, C., G. M. Reihana and H. Dick. 1988. Bud-opening treatments to improve Gypsophila quality after transport. Sci. Hortic. 34: 301–310.
- Eason, J. R., L. A. de Vré, S. D. Somerfield and J. A. Heyes. 1997. Physiological changes associated with Sandersonia aurantiaca flower senescence in response to sugar. Postharvest Biol. Technol. 12: 43–50.
- Fukai, S. and K. Uehara. 2006. Effects of calcium treatment on vase life of cut snapdragon flowers. Hort. Res. (Japan) 5: 465–471 (In Japanese with English abstract).
- Gerasopoulos, D. and B. Chebli. 1999. Effects of pre- and postharvest calcium applications on the vase life of cut gerberas. J. Hort. Sci. Biotechnol. 74: 78–81.
- Halevy, A. H. and S. Mayak. 1979. Senescence of postharvest physiology of cut flowers, part 1. Hort. Rev. 1: 204–326.
- Halevy, A. H. and S. Mayak. 1981. Senescence of postharvest physiology of cut flowers, part 2. Hort. Rev. 3: 59–143.
- Hepler, P. K. 2005. Calcium: A central regulator of plant growth and development. Plant Cell 17: 2142–2155.
- Ichimura, K. 2020. Postharvest technology in cut snapdragon flowers. Agriculture and Horticulture 95: 858–862 (In Japanese).
- Ichimura, K. and T. Hiraya. 1999. Effect of silver thiosulfate complex (STS) in combination with sucrose on the vase life of cut sweet pea flowers. J. Japan. Soc. Hort. Sci. 68: 23–27.
- Ichimura, K. and T. Hisamatsu. 1999. Effects of continuous treatment with sucrose on the vase life, soluble carbohydrate concentrations, and ethylene production of cut snapdragon flowers. J. Japan. Soc. Hort. Sci. 68: 61–66.
- Ichimura, K. and H. Shimizu-Yumoto. 2007. Extension of the vase life of cut rose flowers by treatment with sucrose before and during simulated transport. Bull. Natl. Inst. Flor. Sci. 7: 17–27.
- Ichimura, K. and K. Suto. 1999. Effects of the time of sucrose treatment on vase life, soluble carbohydrate concentrations and ethylene production in cut sweet pea flowers. Plant Growth Regul. 28: 117–122.
- Ichimura, K., K. Kojima and R. Goto. 1999. Effects of temperature, 8-hydroxyquinoline sulphate and sucrose on the vase life of cut rose flowers. Postharvest Biol. Technol. 15: 33–40.
- Ichimura, K., T. Niki, K. Shibuya and H. Shimizu-Yumoto. 2009a. Effects of metabolic sugars on bud growth and aurone biosynthesis in cut snapdragon flowers. Hort. Res. (Japan) 8 (Suppl. 1): 255 (In Japanese).
- Ichimura, K., H. Shimizu-Yumoto and R. Goto. 2009b. Ethylene production by the gynoecium and receptacle is associated with sepal abscission in cut Delphinium flowers. Postharvest Biol. Technol. 52: 262–267.
- Ichimura, K., S. Yoshioka and H. Shimizu-Yumoto. 2008. Effects of silver thiosulfate complex (STS), sucrose and combined pulse treatments on the vase life of cut snapdragon flowers. Environ. Control Biol. 46: 155–162.
- Ichimura, K., S. Yoshioka and T. Yamada. 2016. Exogenous mannitol treatment stimulates bud development and extends vase life of cut snapdragon flowers. Postharvest Biol. Technol. 113: 20–28.
- Kato, M., H. Shimizu, T. Onozaki, N. Tanikawa, H. Ikeda, T. Hisamatsu and K. Ichimura. 2002. Role of ethylene in senescence of pollinated and unpollinated Campanula medium. flowers. J. Japan. Soc. Hort. Sci. 71: 385–387.
- Kebenei, Z., E. Sisler, T. Winkelmann and M. Serek. 2003. Effect of 1-octylcyclopropene and 1-methylcyclopropene on vase life of sweet pea (Lathyrus odoratus L.) flowers. J. Hort. Sci. Biotechnol. 78: 433–436.
- Kuroshima, M., K. Ichimura and M. Ubukata. 2009. Extension of vase life and Ag content in cut flowers of Delphinium treated with STS. Hort. Res. (Japan) 8: 353–357 (In Japanese with English abstract).
- Larsen, F. E. and J. F. Scholes. 1966. Effects of 8-hydroxyquinoline citrate, N-dimethyl amino succinamic acid, and sucrose on vase-life and spike characteristics of cut snapdragons. Proc. Amer. Soc. Hort. Sci. 89: 694–701.
- Marousky, F. J. and J. C. Raulston. 1970. Interaction of flower preservative components and light on fresh weight and longevity of snapdragon cut flowers. Proc. Florida State Hort. Soc. 83: 446–448.
- Moalem-Beno, D., G. Tamari, Y. Leitner-Dagan, A. Borochov and D. Weiss. 1997. Sugar-dependent gibberellin-induced chalcone synthase gene expression in petunia corollas. Plant Physiol. 113: 419–424.
- Mor, Y., M. S. Reid and A. M. Kofranek. 1984. Pulse treatments with silver thiosulfate and sucrose improve the vase life of sweet peas. J. Amer. Soc. Hort. Sci. 109: 866–868.
- Nakayama, T., K. Yoneura-Sakakibara, T. Sato, S. Kikuchi, Y. Fukui, M. Fukuchi-Mizutani, T. Ueda, M. Nakao, T. Tanaka, T. Kusumi and T. Nishio. 2000. Aureusidin synthase: A polyphenol oxidase homolog responsible for flower coloration. Science 290: 1163–1166.
- Norikoshi, R., M. Kato and K. Ichimura. 2008. A simple extraction method of soluble carbohydrates from various organs of several ornamental plants using a vacuum manifold. Environ. Control Biol. 46: 49–55.
- Nowak, J. 1981. Chemical pre-treatment of snapdragon spikes to increase cut-flower longevity. Sci. Hortic. 15: 255–262.
- Pun, U. K., T. Yamada, M. Azuma, K. Tanase, S. Yoshioka, H. Shimizu-Yumoto, S. Satoh and K. Ichimura. 2016. Effect of sucrose on sensitivity to ethylene and enzyme activities and gene expression involved in ethylene biosynthesis in cut carnations. Postharvest Biol. Technol. 121: 151–158.
- Serek, M., E. C. Sisler and M. S. Reid. 1995. Effects of 1-MCP on the vase life and ethylene response of cut flowers. Plant Growth Regul. 16: 93–97.
- Shimamura, M., A. Ito, K. Suto, H. Okabayashi and K. Ichimura. 1997. Effects of α-aminoisobutyric acid and sucrose on the vase life of hybrid Limonium. Postharvest Biol. Technol. 12: 247–253.
- Shimizu, H. and K. Ichimura. 2005. Effects of silver thiosulfate complex (STS), sucrose and their combination on the quality and vase life of cut Eustoma flowers. J. Japan. Soc. Hort. Sci. 74: 381–385.
- Shimizu-Yumoto, H. and K. Ichimura. 2010. Postharvest physiology and technology of cut Eustoma flowers. J. Japan. Soc. Hort. Sci. 79: 227–238.
- Tsukaya, H., T. Ohshima, S. Naito, M. Chino and Y. Komeda. 1991. Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol. 97: 1414–1421.
- Veen, H. 1979. Effects of silver on ethylene synthesis and action in cut carnations. Planta 145: 467–470.
- Woltering, E. J. and W. G. van Doorn. 1988. Role of ethylene in senescence of petals—morphological and taxonomical relationships. J. Exp. Bot. 39: 1605–1616.
- Woodson, W. R. 1987. Postharvest handling of bud-cut freesia flowers. HortScience 22: 456–458.
- Wu, M. J., L. Zacarias and M. S. Reid. 1991. Variation in the senescence of carnation (Dianthus caryophyllus L.) cultivars. II. Comparison of sensitivity to exogenous ethylene and of ethylene binding. Sci. Hortic. 48: 109–116.
- Yamada, K., R. Norikoshi, K. Suzuki, H. Imanishi and K. Ichimura. 2009. Determination of subcellular concentrations of soluble carbohydrates in rose petals during opening by nonaqueous fractionation method combined with infiltration–centrifugation method. Planta 230: 1115–1127.
- Yamada, T., Y. Takatsu, M. Kasumi, K. Ichimura and W. G. van Doorn. 2006. Nuclear fragmentation and DNA degradation during programmed cell death in petals of morning glory (Ipomoea nil). Planta 224: 1279–1290.
- Yamane, K., Y. Kotake, T. Okada and R. Ogata. 1995. Export of 14C-sucrose, 3H-water, and fluorescent tracers from gladiolus florets to other plant parts associated with senescence. Acta Hortic. 405: 269–276.