摘 要
本研究ではコムギの穂発芽性“極難”に関わる遺伝的要因を解明することを目的に,“極難”系統の「北系1802」と「きたほなみ」の交雑による分離集団を北海道内の地域と年次の異なる6環境下で栽培し,穂発芽耐性と種子休眠性に対するQTLの探索を行った.穂発芽耐性は晩刈りした切り穂を用いた人工降雨処理後の穂発芽指数にて,種子休眠性は同じく晩刈りした種子を用いた15℃での発芽率にて評価した.穂発芽耐性では4個のQTLが検出された.そのうち2D染色体長腕のQTL(以下QPhs-2D)の効果は,6環境条件全てで安定して認められた.種子休眠性遺伝子MFT-3Aの穂発芽耐性に対する効果は3環境で認められ,加えて1A染色体と7B染色体にもそれぞれ1環境でQTLの効果が認められた.これらQTLはいずれも「北系1802」アレルが耐性を高めていた.一方,15℃での発芽率で示される種子休眠性は栽培環境全てでMFT-3Aにのみ明確な効果が認められ,「北系1802」アレルが種子休眠性を維持する方向に作用していた.穂発芽耐性QTL,QPhs-2Dはいずれの条件でも種子休眠性に対しては明らかな効果を示さなかった.これらの結果から,「北系1802」の穂発芽性“極難”には穂発芽耐性と種子休眠性が複合的に影響し,穂発芽耐性には種子休眠性以外の要因に関与するQPhs-2Dが,種子休眠性にはMFT-3Aが大きく貢献していることが明らかになった.
Abstract
To identify quantitative trait loci (QTL) that confer excellent resistance to pre-harvest sprouting (PHS) in wheat, levels of sprouting in matured spikes and grain dormancy were investigated in a population of doubled haploid lines derived from a cross between the breeding line “Kitakei 1802” (highly tolerant to PHS) and the local leading variety “Kitahonami”. The doubled haploid population and parental genotypes were grown in six different environments. As sprouting resistance and grain dormancy were gradually lost after maturity, spikes were collected 10 days after physiological maturity. Assessment for PHS resistance was based on a sprouting assay conducted during 7–10 days artificial rain treatment on intact spikes, whereas grain dormancy was evaluated in germination tests at 15°C. Four QTL in total were detected in the sprouting assay. A QTL on chromosome 2D, designated as QPhs-2D, was found to be a stable QTL with a large effect across all six environments. The effect of the MOTHER OF FT AND TFL1 (MFT-3A) gene was detected in three environments. The remaining two QTL with minor effects were identified on chromosomes 1A and 7B in single environments. The resistant alleles at these QTL were all derived from “Kitakei 1802”. The effect of MFT-3A on grain dormancy was evident whereas the effect of QPhs-2D was unclear. These results demonstrated that incorporating QPhs-2D for sprouting tolerance along with MFT-3A for grain dormancy could markedly contribute to excellent PHS resistance.
緒言
コムギの穂発芽とは,降雨の影響により収穫期前後の立毛状態で種子が発芽する現象である.穂発芽は収量低下の直接的な要因となるばかりではなく,子実の外観品質の悪化,α-アミラーゼ等の酵素の影響による内部品質の劣化を引き起こし,コムギの商品価値を著しく低下させる.穂発芽の生じやすさは種子休眠性と強く関連している.種子が成熟する前の期間が高温で推移した時は種子の休眠が浅く(Lunn et al. 2002,中園ら 2013),さらに,成熟期後の低温や降雨に遭遇することで種子休眠が打破されやすい(Osanai and Amano 1993).
穂発芽回避のためには,梅雨等の雨害回避を目的とした早生品種の選定や,コムギの成熟後に速やかに収穫,乾燥を行うなど栽培上の対策があるものの,最も有効な対策は遺伝的改良による耐性品種の開発である.アジアモンスーンの影響を受けるわが国では古くから雨害に強いコムギが選抜されており,関東以西で栽培されてきた品種は世界的に見て穂発芽耐性に優れるものが多い(Nakamura 2018).その一方で,北海道のコムギ品種はその来歴や環境,栽培条件から穂発芽耐性が中程度のものが多く(柳沢 2004),品種開発に際して耐性強化が図られてきた.そのアプローチとして穂発芽耐性に優れるコムギの選抜は,種子休眠性が弱まっている成熟期1週間後にサンプルを採取し(晩刈り),切り穂を用いた人工降雨処理による穂発芽検定と休眠打破を誘導する低温での種子休眠性評価とを組み合わせて実施してきた(柳沢 2004).この選抜から穂発芽性“極難”にランクされる「北系1802」が育成された.この系統は穂発芽耐性に極めて優れており,強い種子休眠性をもち,長期降雨を模した条件下でも穀粒のアミロースが低下しにくいという特徴をもつ(西村ら 2004,Yanagisawa et al. 2005).
これまでコムギでは穂発芽耐性とその主要因である種子休眠性に関する遺伝的知見が数多く蓄積されてきており,21対の染色体全てにQTLが報告されている(Mares and Mrva 2014, Kumar et al. 2015).そのなかで注目されているQTLの一つに,「ゼンコウジコムギ」(Zen)で発見された3A染色体短腕の種子休眠性QTL,QPhs.ocs-3A.1がある(Osa et al. 2003, Mori et al. 2005).Nakamura et al.(2011)は登熟期の温度の違いによって種子胚での発現量が大きく異なる遺伝子としてMOTHER OF FT AND TFL1(MFT-3A)を見いだし,QPhs.ocs-3A.1の原因遺伝子であることを明らかにした.Zenでは,MFT-3Aの発現量を調節するプロモーター領域に1塩基の置換(SNP)があり,それによってZenではMFT-3Aの転写レベルが上昇し,強い種子休眠性を誘導すると考えられた.
Chono et al.(2015)は日本で育成された183品種を含む324品種を供試して,MFT-3AのSNPを調査した.その結果関東以西では,Zen型の品種が多数見られ,系譜からMFT-3A遺伝子の来歴は多岐にわたることが明らかとなった.一方北海道で育成された品種あるいは在来品種計25品種・系統中にZen型のSNPは認められなかった.
このように穂発芽耐性や種子休眠性に関わる遺伝的要因については数多くの知見が蓄積されてきた.しかし,穂発芽性“極難”ランクの耐性に関する解析はほとんどなされていない.そこで本研究では穂発芽性“極難”に関わる遺伝的要因を解明することを目的とし,「北系1802」を片親とする分離集団を北海道内の地域と年次の異なる環境下で栽培し,穂発芽耐性と種子休眠性に対するMFT-3Aの効果を明らかにするとともに,新たなQTLの探索を行った.
材料および方法
1. 供試材料
実験には「北系1802」,「きたほなみ」および「きたほなみ」×「北系1802」の交雑から葯培養により育成した倍加半数体117系統(以下DH系統)を供試した.「北系1802」は道総研北見農業試験場とホクレン農業総合研究所との共同事業により育成された穂発芽性が“極難”の系統である(西村ら 2004,Yanagisawa et al. 2005).「きたほなみ」は現在の北海道の基幹品種で,もともと多収品種を目的に育成された,穂発芽性が“やや難”の品種である(柳沢ら 2007).
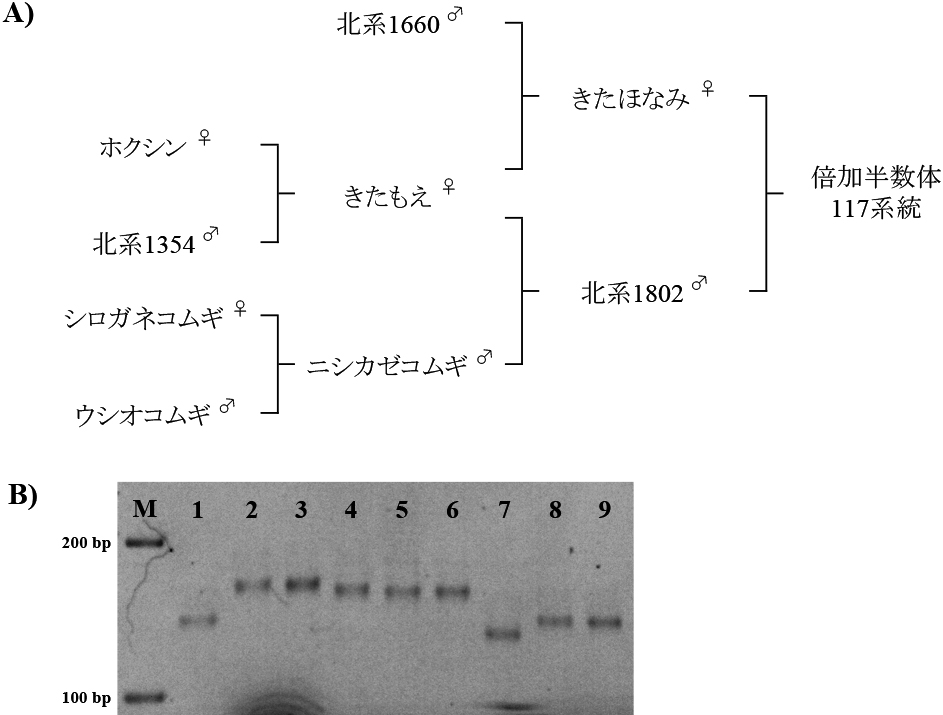
さらに検出されたQTLについて,穂発芽耐性あるいは強種子休眠性アレルの来歴を知るために,「北系1802」と「きたほなみ」の系譜上にある以下の品種のDNAを解析した.「北系1802」については,種子親の「きたもえ」とその親の「ホクシン」および「北系1354」,ならびに花粉親の「ニシカゼコムギ」とその親の「シロガネコムギ」および「ウシオコムギ」を供試した.「きたほなみ」については種子親が「北系1802」と共通の「きたもえ」で,花粉親は「北系1660」である.
2. 圃場栽培
親品種とDH系統を2014,2015および2016年に帯広畜産大学実験圃場(以後,帯広)で秋播き栽培した.同様に2014年に道総研北見農業試験場(北見)で,また,2015年と2016年に道総研上川農業試験場(上川)で秋播き栽培した.以降では収穫年次で示す.帯広での3ヶ年の試験および上川での2ヶ年の試験では,対照品種として「北系1802」と「きたほなみ」の系譜上にある「ホクシン」,「きたもえ」も加えた.
播種日は9月中旬で,栽植密度,施肥および病害虫防除は各試験地の慣行法に従って行った.各系統の全茎の40~50%が出穂,開花した時をそれぞれ出穂期,開花期として記録した.成熟期は子実が黄熟し蝋状の硬さを示した時(生理的成熟期)とした.2015年の帯広,北見および2016年の上川,2017年の帯広と上川の試験では,成熟期10日後に収穫した(以後,晩刈り).2016年帯広の試験では成熟期の7月下旬から降雨が続き,さらに8月上旬に台風の通過が予想されたため,晩刈りを成熟期後の3~5日目に早めた.帯広と上川の試験では「北系1802」,「きたほなみ」,「ホクシン」,「きたもえ」の4品種では成熟期の穂もサンプリングした.
刈り取った穂は,40℃の通風乾燥機で24時間乾燥させた後,−20℃で冷凍保存した.その後,室温で24時間解凍した.切り穂のサンプルは穂発芽耐性の評価に用い,また,手もみ脱穀した種子は休眠性を評価するための発芽試験に用いた.
3. 穂発芽耐性の評価
穂発芽耐性は,切り穂を降雨処理した後の穂発芽指数にて評価した.Yanagisawa et al.(2005)の方法に従い,人工気象室内にて各系統10~15穂(2017年の上川のサンプルは7穂)を立て,15℃の暗黒条件下で朝夕人工降雨処理を10日間(2015年は7日間)行った後,外観上で各穂の発芽・発根数を数え,0~5までのスコアで穂発芽指数を求めた.穂内で発芽・発根が全く認められないものをスコア0とし,発芽あるいは発根した種子が10個以上の穂をスコア5とした.
4. 種子休眠性の評価
帯広と上川で栽培し,晩刈りした材料を用いて15℃での発芽試験を行った.1,500倍に希釈したベフラン液剤25で消毒した種子を直径90 mmのシャーレにろ紙を敷き5 mLの蒸留水を入れて置床した.1シャーレに50粒を置床し,これらのシャーレを暗黒下のインキュベーター内に静置した.試験は2反復で行った.ただし,2017年の上川の材料は前年10月下旬の根雪によって雪腐れ病の防除ができず十分な個体数を確保できなかったため,実験からは除いた.
胚の果皮が破れたものを発芽と見なし,10日間の累積発芽種子数を調査し,発芽率(%)を算出した.
5.DNA抽出とMFT-3AのSNP解析
品種系統のDNA抽出はCao et al.(2016)に従った.前述したChono et al.(2015)の報告から,「きたほなみ」のMFT-3Aプロモーター領域のSNPは非Zen型であることが分かっていたが,「北系1802」とDH系統は未解析であった.そのため,これら材料についてCAPSマーカー(Nakamura et al. 2011)でMFT-3Aプロモーター領域のSNPを解析した.
6. 連鎖地図の構築とQTL解析
連鎖地図の作成にはSomers et al.(2004)とTorada et al.(2006)の報告にあるSSRマーカー408種およびXue et al.(2008)が開発したSTSマーカー4種について,まず親品種系統間で多型調査を行い,多型の見られたマーカーについてDH系統の遺伝子型を判別した.
遺伝子型データをもとに,JoinMap® 4(van Ooijen 2006)を利用して連鎖地図を構築した.DH系統の穂発芽指数と発芽試験データに対してMapQTL® 6(van Ooijen 2002)のintervelマッピング法およびmultiple-QTL models(MQM)マッピング法を用いてQTL解析を行った.LOD値の閾値については各形質でpermutation testにより算出した.
結果
1. 穂発芽耐性
図1は2015年帯広で成熟期刈りと晩刈りし,7日間人工降雨処理した後の「きたほなみ」,「北系1802」および対照品種の「ホクシン」と「きたもえ」の穂の状態である.成熟期刈りでは4品種系統の穂発芽指数は1.8以下で変異も小さかった(図1A).晩刈りしたときの「北系1802」の穂発芽指数は0.14と低かったのに対し,「きたほなみ」は4.70,他の2品種も4.60~5.00と大きな差異があった(図1B).他の環境条件で栽培した時も「ホクシン」と「きたもえ」は同様の傾向を示した.
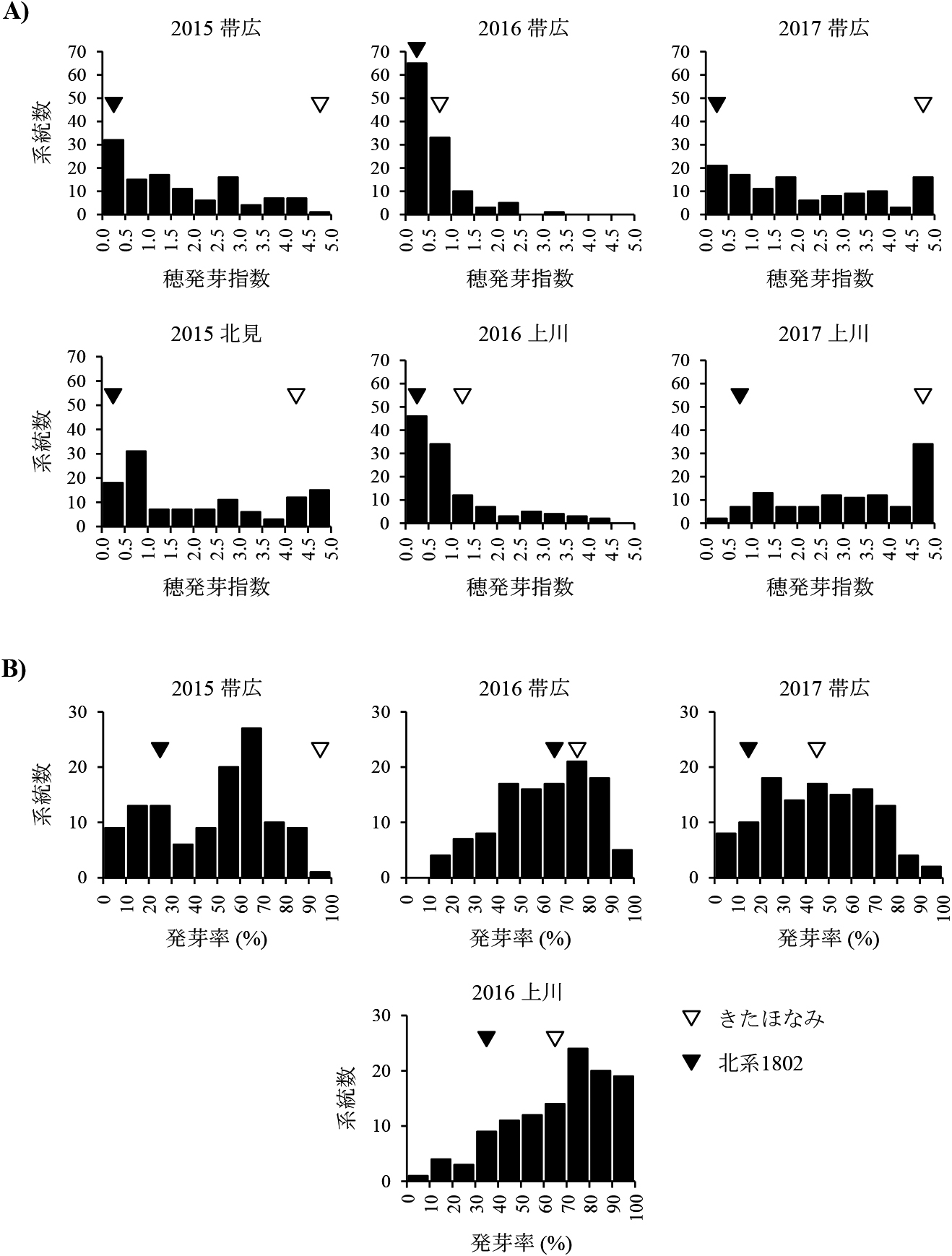
親およびDH117系統の晩刈りした穂サンプルにおける穂発芽指数の頻度分布を図2Aに示した.帯広で栽培した「北系1802」の穂発芽指数は3ヶ年とも0.5以下で,安定して強い穂発芽耐性を示した.一方「きたほなみ」は2016年が0.93と低かった.この年の「きたほなみ」は上川でも1.32と低い穂発芽指数であった.他の2ヶ年は帯広で4.50以上と高かった.北見,上川でも同様の値であった.DH系統の穂発芽指数は,2015年の帯広,北見,2017年の帯広,上川で0.0~5.0まで幅広い分布を示した.これら4環境でのDH系統の平均値は2015年帯広の1.64から2017年上川の3.20の間であった.それに対して,「きたほなみ」の穂発芽指数が低かった2016年の帯広と上川では,DH系統の平均値がそれぞれ0.60と1.00で,分布の幅は小さく穂発芽指数1.0以下のDH系統が90%以上であった.特に帯広ではDH系統の分布幅が狭く,穂発芽指数が2.5以上の系統はわずか1系統であった.上川では「きたほなみ」よりも穂発芽指数が高く超越的に分離するDH系統が見られた.
2. 種子休眠性
親品種とDH117系統の発芽率の頻度分布を図2Bに示した.親の「北系1802」と「きたほなみ」の発芽率には2016年の帯広を除いて明確な差異があり,常に「北系1802」の発芽率が低く強い種子休眠性を示した.成熟期の7月下旬から連続した降雨のため晩刈りを成熟期3~5日後に早めた2016年の帯広では,「北系1802」の発芽率は63.0%と他の栽培環境を大きく上回っていた.
DH系統は4条件いずれでも連続した分布を示した.2016年の帯広と上川ではともに半数以上の系統が60%以上の高い発芽率を示した.2016年の上川と2017年の帯広では,「きたほなみ」よりも高い発芽率を示すDH系統が多く見られた.逆に2016年の帯広では「北系1802」より発芽率が低い超越的な系統が見られた.「北系1802」と「きたほなみ」の成熟期には全ての環境で差異はなかった.
3. 穂発芽耐性と種子休眠性の相関
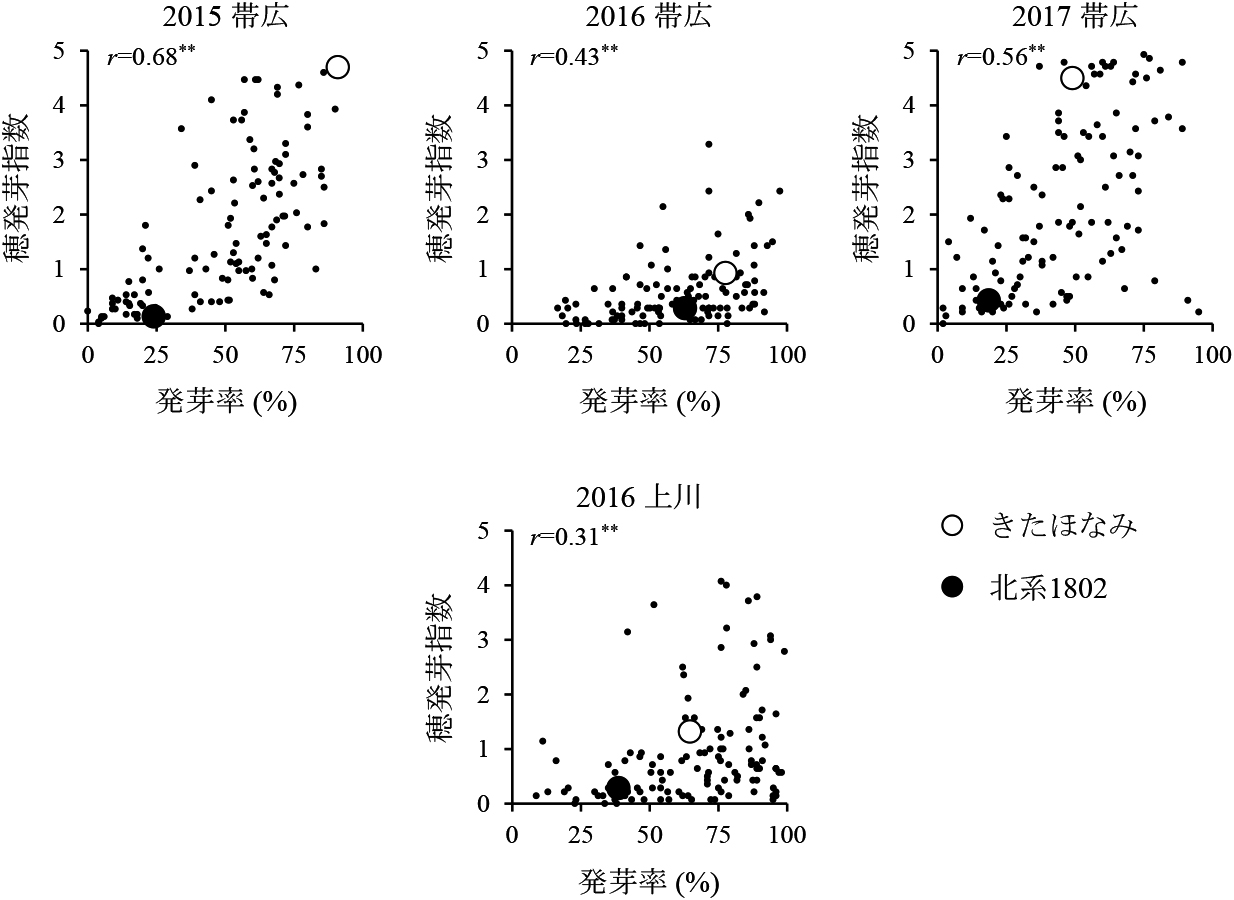
2015年の北見と2017年の上川を除いた4環境でのDH117系統の穂発芽指数と発芽率との関係を図3に示した.4環境全てにおいて穂発芽指数と発芽率の間には正の相関があり(P<0.01),2015年と2017年の帯広では相関係数が比較的高かった.一方,2016年の帯広と上川では穂発芽指数が低いにも関わらず発芽率が高いDH系統が多数見られたため相関係数が低くなった.
4.MFT-3AのSNP解析
CAPSマーカーを用いたMFT-3Aプロモーター領域のSNP解析により,「北系1802」はZen型であることが分かった.DH117系統は,Zen型が46系統,残り71系統は「きたほなみ」と同じ非Zen型に分類された.
5. 連鎖地図の構築
親間で多型のあったマーカー数は全体で125種あった(表1).このうち,近接するマーカーを除いた86種を用いてDH系統の遺伝子型を決定し,連鎖地図を構築した.コムギの21本の染色体のうち,1D,5D,6D,7D染色体を除く17染色体で連鎖地図が構築できた.
表1.
両親間の多型解析に使用した染色体毎のマーカー数と検出した多型数
| 染色体 |
1A |
1B |
1D |
2A |
2B |
2D |
3A |
3B |
3D |
4A |
4B |
4D |
5A |
5B |
5D |
6A |
6B |
6D |
7A |
7B |
7D |
合計 |
| マーカー数 |
10 |
16 |
19 |
11 |
21 |
37 |
16 |
20 |
22 |
17 |
14 |
11 |
23 |
19 |
25 |
43 |
18 |
13 |
16 |
21 |
20 |
412 |
| 多型数 |
7 |
4 |
2 |
6 |
4 |
5 |
2 |
13 |
6 |
5 |
4 |
3 |
9 |
4 |
3 |
21 |
6 |
3 |
9 |
7 |
2 |
125 |
穂発芽指数および発芽率から検出されたQTLを表2にまとめた.穂発芽耐性を示す穂発芽指数に対しては6環境条件で合計11個のQTLが検出できた.そのうち2D染色体長腕のSSRマーカー座gwm539近傍領域のQTL(以下QPhs-2D)効果は,6環境条件全てで認められた.LOD値は3.29~6.89で寄与率は10.4%~23.7%であった.「北系1802」由来のアレルが常に穂発芽を抑える効果をもっていた.このアレルは,2017年帯広で効果が最大で,「きたほなみ」のアレルに比較し,穂発芽指数を1.56低下させることが分かった.MFT-3Aの効果は2015年,2017年の帯広と2017年上川の3環境で認められ,LOD値,寄与率は2015年帯広で最も高かった.QPhs-2D同様「北系1802」由来のアレルが穂発芽指数を低下することが分かった.残り2個のQTLは2015年の北見で7B染色体と2016年に帯広で1A染色体に検出された.それぞれ1環境でのみ検出されたことやLOD値と寄与率から見て,QPhs−2DやMFT-3Aに比べマイナーな効果をもつQTLであった.ともに「北系1802」由来のアレルが穂発芽指数を低くした.
表2.
MQMマッピング法により検出された穂発芽耐性QTLと種子休眠性QTLの位置とその効果
| 形質 |
年 |
栽培地 |
染色体 |
マーカーあるいは遺伝子 |
LOD値 |
寄与率(%) |
相加効果1) |
| 穂発芽耐性 |
2015 |
帯広 |
3A |
MFT-3A |
5.49 |
16.7 |
0.55 |
| 2D |
gwm539 |
4.41 |
13.1 |
0.50 |
| 北見 |
2D |
gwm539 |
5.47 |
15.0 |
0.67 |
| 7B |
gwm297 |
3.54 |
9.3 |
0.53 |
| 2016 |
帯広 |
2D |
gwm539 |
3.29 |
10.4 |
0.20 |
| 1A |
gwm99 |
2.80 |
8.7 |
0.18 |
| 上川 |
2D |
gwm539 |
4.48 |
16.3 |
0.42 |
| 2017 |
帯広 |
2D |
gwm539 |
6.89 |
23.7 |
0.78 |
| 3A |
MFT-3A |
2.74 |
7.7 |
0.44 |
| 上川 |
2D |
gwm539 |
5.90 |
18.7 |
0.66 |
| 3A |
MFT-3A |
3.42 |
10.3 |
0.48 |
| 種子休眠性 |
2015 |
帯広 |
3A |
MFT-3A |
12.82 |
39.6 |
15.6 |
| 2016 |
帯広 |
3A |
MFT-3A |
5.55 |
18.1 |
9.1 |
| 上川 |
3A |
MFT-3A |
10.63 |
34.2 |
13.8 |
| 2017 |
帯広 |
3A |
MFT-3A |
12.20 |
38.2 |
14.3 |
1) 「北系1802」のアレルにより各形質値を低下させる効果を示す.穂発芽耐性は穂発芽指数(0~5),種子休眠性は発芽率(%)による.
種子休眠性を示す発芽率に対しては4栽培環境全てでMFT-3Aの効果のみ検出された.LOD値は2016年帯広で5.55とやや低かったが,他の3環境では10以上で,寄与率も33.2~39.6%と大きな効果を示した.「北系1802」のアレルが発芽率を抑えていた.穂発芽指数で検出されたQTLのQPhs-2Dはいずれの条件でも発芽率に対しては明らかな効果を示さなかった.
8.MFT-3AとQPhs-2Dの組み合わせによる効果
MFT-3A座とQPhs-2D座のアレルの違いによってDH系統を4組み合わせに分類し,それらの平均穂発芽指数を表3に示した.6環境条件いずれでもこれら2座に「北系1802」アレルをもつと,他の3組み合わせより平均穂発芽指数は低下していた.統計的には2環境条件で有意であった.逆に「きたほなみ」アレルの組み合わせは4組み合わせの中で平均穂発芽指数が最も高く,「北系1802」アレルの組み合わせとの間で常に有意差があった.親品種と比較すると「北系1802」アレルの組み合わせは「北系1802」より穂発芽指数は高い傾向にあり,一方「きたほなみ」アレルの組み合わせは「きたほなみ」より低い傾向にあった.
表3.
DH系統の平均穂発芽指数におよぼす
QPhs-2Dと
MFT-3Aとの組合わせ効果
| 遺伝子型 |
系統数 |
2015 |
2016 |
2017 |
| gwm539 |
MFT-3A |
帯広 |
北見 |
帯広 |
上川 |
帯広 |
上川 |
| A |
A |
19 |
0.35 a |
0.75 a |
0.20 a |
0.39 a |
0.72 a |
1.71 a |
| A |
B |
27 |
1.47 b |
1.54 ab |
0.48 ab |
0.52 a |
1.50 ab |
2.85 b |
| B |
A |
23 |
1.24 b |
2.37 bc |
0.61 ab |
1.02 ab |
2.09 b |
3.17 b |
| B |
B |
38 |
2.61 c |
3.04 c |
0.81 b |
1.54 b |
3.24 c |
4.12 c |
| 親品種・ |
北系1802 |
0.14 |
0.10 |
0.29 |
0.29 |
0.43 |
0.93 |
| 系統 |
きたほなみ |
4.70 ** |
4.25 |
0.93 + |
1.32 + |
4.50 * |
4.50 ** |
遺伝子型のA,Bはそれぞれ「北系1802」,「きたほなみ」のアレルを示す.英文字が付された値はTukey-Kramerの有意差検定において異文字間に5%水準で有意差があることを示す.+,*,**はt検定でそれぞれ10%,5%,1%水準で親間に有意差があることを示す.
穂発芽耐性あるいは強種子休眠性アレルが「北系1802」または「きたほなみ」のいずれの系譜から遺伝したかを近傍マーカーの遺伝子型から推定した.図4Aと図4Bに示した結果から,穂発芽耐性を高めるQPhs-2D座の「北系1802」アレルは「シロガネコムギ」から「ニシカゼコムギ」を経て遺伝したと推定された.MFT-3Aおよび1A染色体QTLおよび7B染色体QTLに連鎖するマーカーについても「北系1802」アレルは「ニシカゼコムギ」由来であった.
考察
本研究では,年次と地域の異なる6環境条件でDH系統を栽培し,種子休眠性が低下している晩刈りした切り穂を穂発芽検定のサンプルに用いた.15℃の暗黒条件下で朝夕人工降雨処理を10日間(2015年は7日間)行って得た穂発芽指数を指標とした時,4個のQTLが検出できた.そのうち2D染色体長腕のSSRマーカーgwm539近傍領域にQPhs-2Dの効果が6環境条件全てで認められた.このQTLに次いで効果があったのはMFT-3Aで,3環境で検出できた.加えて1A染色体と7B染色体にそれぞれ1環境で小さな効果をもつQTLが検出できた.これら4個のQTLはいずれも「北系1802」のアレルが常に穂発芽を抑える効果をもっており,全て「ニシカゼコムギ」に由来することが分かった.したがって,「北系1802」のもつ高い穂発芽耐性は遺伝的にはQPhs-2D座とMFT-3A座の「ニシカゼコムギ」に由来するアレルが大きく貢献していることが明らかになった.さらに環境条件によってはマイナーなQTLの効果も認められた.
ただし,本研究では117系統のDH集団を用いてSSRマーカーによる連鎖地図の構築を目指したが,マーカーが極少数だった染色体もあり,ゲノム全体を網羅した解析にはなっていない.これは両親が共通親として「きたもえ」をもち(図3),また実用品種の開発を目指した交配組み合わせだったため遺伝的に多様性が低いことが一因と考えられた.したがって,それら染色体に検出できなかったQTLが存在する可能性があり,その解析が今後の課題である.
2D染色体に座乗する穂発芽耐性QTLについてはこれまでいくつか報告があり,その多くが短腕あるいは動原体領域にマップされている(Kulwal et al. 2004, Imtiaz et al. 2008, Munkvold et al. 2009, Kulwal et al. 2012).一方,Mohan et al.(2009)は2D染色体長腕に2個のQTLを検出し,その一つが本試験でのQPhs-2Dと同じSSRマーカーgwm539と連鎖していた.そのためQPhs-2Dと同一のQTLである可能性もあるが,マップ間で近傍領域に共通するマーカーがこれ以外ないため結論はできない.ともあれ,今後QPhs-2Dの実用育種への活用を図る上で,利便性や正確さを向上させるためにも,この領域でQPhs-2Dを正確にマップする必要がある.
次にMFT-3Aについてであるが,府県で育成された品種では,MFT-3AのZen型アレルが広く分布し,穂発芽耐性の向上に大きく貢献してきた(Chono et al. 2015).その一方で,北海道で育成された品種あるいは在来品種にZen型のSNPを保有するものはなかった.しかし,現在,道総研北見農業試験場や農研機構北海道農業研究センターではこのアレルを取り込んだ北海道向け新品種の開発が進んでいる.本試験の結果から,北海道の栽培環境でもMFT-3AのZen型アレルは確かに強い種子休眠性を通じて穂発芽耐性の向上に貢献することが実証できたため,今後北海道品種への導入に期待がもたれる.
穂発芽耐性は環境要因に加え,穂や種子の生理的特性が関与する複雑な形質である.その中でも特に種子の休眠性と密接な関係にあることがよく知られている.本研究では,穂発芽耐性と並行して種子休眠性も評価し,QTLの検出を行った.その結果,両形質で検出したQTLの間に違いが見られた.穂発芽指数では安定して認められたQPhs-2Dの効果が,栽培環境に関わらず種子休眠性に対しては明確ではなかった.図3に示した2016年の帯広と上川のように,穂発芽指数が低い系統の中に発芽率の高いものも複数含まれていた.その一方で種子休眠性は,4栽培環境で共通してMFT-3Aのみに大きな効果が認められた.この結果はMFT-3Aが種子胚で発現し直接休眠性に関わるのに対し(Nakamura et al. 2011),QPhs-2Dの効果には種子休眠性以外の要因が関与していることを示唆している.
これまで穂発芽耐性と種子休眠性を同時に評価し,QTL解析を行った結果がいくつか報告されている.それらを要約すると4A染色体長腕の種子休眠性遺伝子Phs1(Torada et al. 2008)のように,両形質に安定して効果を示す場合(Chen et al. 2008, Ogbonnaya et al. 2008, Cabral et al. 2014, Lin et al. 2015)もあれば,本試験のQPhs-2Dと同様に,穂発芽耐性には常に効果を発揮するQTLが種子休眠性には明確な効果を示さない場合(Lin et al. 2015),あるいはその逆の場合など様々である.確かに穂発芽耐性は種子休眠性と密接な関係にあるものの,両形質の遺伝的支配は一義的ではないと言える.穂発芽耐性に関わる種子休眠性以外の要因として,中津(2000)は穂に直接加水したときの穂中種子の吸水速度を調査した結果から,吸水速度が遅いほどアミラーゼ活性が低く穂発芽の発生が抑えられたことを指摘し,頴の透水性や穂の構造が影響していると推測している.QPhs-2Dがこれらに関与しているかは今のところ明らかでない.
現在の育種現場においては,F2個体から世代毎に穂発芽指数や発芽率などの表現型による穂発芽耐性の選抜を行うことで,育種材料全体の穂発芽耐性系統の頻度を高めている.今後,QPhs-2Dに対する高精度なDNAマーカーが開発され,MFT-3Aとともに育種プログラムに組み込まれることで,穂発芽耐性育種がさらに効率化することが期待される.
謝辞
本研究で用いた「ホクシン」,「きたもえ」,「北系1354」および「北系1660」は道総研北見農業試験場より分譲を受けた.「ニシカゼコムギ」,「シロガネコムギ」および「ウシオコムギ」の3品種は農業生物資源研究所(現農研機構)遺伝資源センターより分譲を受けた.ここに記して謝意を表します.
引用文献
- Cabral, A.L., M.C. Jordan, C.A. McCartney, F.M. You, D.G. Humphreys, R. MacLachlan and C.J. Pozniak (2014) Identification of candidate genes, regions and markers for pre-harvest sprouting resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 14: 340.
- Cao, L., K. Hayashi, M. Tokui, M. Mori, H. Miura and K. Onishi (2016) Detection of QTLs for traits associated with pre-harvest sprouting resistance in bread wheat (Triticum aestivum L.). Breed. Sci. 66: 260–270.
- Chen, C.X., S.B. Cai and G.H. Bai (2008) A major QTL controlling seed dormancy and pre-harvest sprouting resistance on chromosome 4A in a Chinese wheat landrace. Mol. Breed. 21: 351–358.
- Chono, M., H. Matsunaka, M. Seki, M. Fujita, C. Kiribuchi-Otobe, S. Oda, H. Kojima and S. Nakamura (2015) Molecular and genealogical analysis of grain dormancy in Japanese wheat varieties, with specific focus on MOTHER OF FT AND TFL1 on chromosome 3A. Breed. Sci. 65: 103–109.
- Imtiaz, M., F.C. Ogbonnaya, J. Oman and M.V. van Ginkel (2008) Characterization of quantitative trait loci controlling genetic variation for preharvest sprouting in synthetic backcross-derived wheat lines. Genetics 178: 1725–1736.
- Kulwal, P.L., R. Singh, H.S. Balyan and P.K. Gupta (2004) Genetic basis of pre-harvest sprouting tolerance using single-locus and two-locus QTL analyses in bread wheat. Funct. Integr. Genomics 4: 94–101.
- Kulwal, P., G. Ishikawa, D. Benscher, Z. Feng, L.X. Yu, A. Jadhav, S. Mehetre and M.E. Sorrells (2012) Association mapping for pre-harvest sprouting resistance in white winter wheat. Theor. Appl. Genet. 125: 793–805.
- Kumar, S., R.E. Knox, F.R. Clarke, C.J. Pozniak, R.M. DePauw, R.D. Cuthbert and S. Fox (2015) Maximizing the identification of QTL for pre-harvest sprouting resistance using seed dormancy measures in a white-grained hexaploid wheat population. Euphytica 205: 287–309.
- Lin, M., S. Cai, S. Wang, S. Liu, G. Zhang and G. Bai (2015) Genotyping-by sequencing (GBS) identified SNP tightly linked to QTL for pre-harvest sprouting resistance. Theor. Appl. Genet. 128: 1385–1395.
- Lunn, G.D., P.S. Kettlewell, B.J. Major and R.K. Scott (2002) Variation in dormancy duration of the U.K. wheat cultivar Hornet due to environmental conditions during grain development. Euphytica 126: 89–97.
- Mares, D.J. and K. Mrva (2014) Wheat grain preharvest sprouting and late maturity alpha-amylase. Planta 240: 1167–1178.
- Mohan, A., P. Kulwal, R. Singh, V. Kumar, R.R. Mir, J. Kumar, M. Prasad, H.S. Balyan and P.K. Gupta (2009) Genome-wide QTL analysis for pre-harvest sprouting tolerance in bread wheat. Euphytica 168: 319–329.
- Mori, M., N. Uchino, M. Chono, K. Kato and H. Miura (2005) Mapping QTLs for grain dormancy on wheat 3A and group 4 chromosomes, and their combined effect. Theor. Appl. Genet. 110: 1315–1323.
- Munkvold, J.D., J. Tanaka, D. Benscher and M.E. Sorrells (2009) Mapping quantitative trait loci for preharvest sprouting resistance in white wheat. Theor. Appl. Genet. 119: 1223–1235.
- Nakamura, S. (2018) Grain dormancy genes responsible for preventing pre-harvest sprouting in barley and wheat. Breed. Sci. 68: 295–304.
- Nakamura, S., F. Abe, H. Kawahigashi, K. Nakazono, A. Tagiri, T. Matsumoto, S. Utsugi, T. Ogawa, H. Handa, H. Ishida et al. (2011) A wheat homolog of MOTHER OF FT AND TFL1 acts the regulation of germination. Plant Cell 23: 3215–3229.
- 中津 智史(2000)北海道における低アミロ小麦の発生とその要因に関する研究.北海道立農業試験場報告 93: 1–57.
- 中園 江・ 大野 宏之・ 吉田 ひろえ・ 中川 博視(2013)登熟段階別の気象要因がコムギの穂発芽発生に及ぼす影響.日作紀 82: 183–191.
- 西村 努・ 柳沢 朗・ 天野 洋一・ 乕田 淳史・ 新発田 修司(2004)穂発芽性極難秋播小麦系統の育成.2. 穂発芽性極難系統の特性.育種学研究 6(別1): 310.
- Ogbonnaya, F.C., M. Imtiaz, G. Ye, P.R. Hearnden, E. Hernandez, R.F. Eastwood, M. van Ginkel, S.C. Shorter and J.M. Winchester (2008) Genetic and QTL analyses of seed dormancy and preharvest sprouting resistance in the wheat germplasm CN10955. Theor. Appl. Genet. 116: 891–902.
- Osa, M., K. Kato, M. Mori, C. Shindo, A. Torada and H. Miura (2003) Mapping QTLs for seed dormancy and the Vp1 homologue on chromosome 3A in wheat. Theor. Appl. Genet. 106: 1491–1496.
- Osanai, S. and Y. Amano (1993) Selection of tolerant lines to low temperature germinability. In: Walker-Simmons, M.K. and J.L.Ried (eds.), “Pre-Harvest Sprouting in Cereals 1992”, American Association of Cereal Chemistry, MN, USA, 76–83.
- Somers, D.J., P. Isaac and K. Edwards (2004) A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 109: 1105–1114.
- Torada, A., M. Koike, K. Mochida and Y. Ogihara (2006) SSR-based linkage map with new markers using an intraspecific population of common wheat. Theor. Appl. Genet. 112: 1042–1051.
- Torada, A., M. Koike, S. Ikeguchi and I. Tsutsui (2008) Mapping of a major locus controlling seed dormancy using backcrossed progenies in wheat (Triticum aestivum L.). Genome 51: 426–432.
- van Ooijen, J.W. (2002) MapQTL® 6, Software for the mapping of quantitative trait loci in experimental populations of diploid species. Kyazma B.V., Wageningen, The Netherlands.
- van Ooijen, J.W. (2006) JoinMap® 4, Software for the calculation of genetic linkage maps in experimental population. Kyazma B.V., Wageningen, The Netherlands.
- Xue, S., Z. Zhang, F. Lin, Z. Kong, Y. Cao, C. Li, H. Yi, M. Mei, H. Zhu, J. Wu et al. (2008) A high-density intervarietal map of the wheat genome enriched with markers derived from expressed sequence tags. Theor. Appl. Genet. 118: 181–189.
- 柳沢 朗(2004)穂発芽耐性評価と穂発芽性極難系統の育成.冬作物研究 4: 13–20.
- Yanagisawa, A., T. Nishimura, Y. Amano, A. Torada and S. Shibata (2005) Development of winter wheat with excellent resistance to pre-harvest sprouting and rain damage. Euphytica 143: 313–318.
- 柳沢 朗・ 吉村 康弘・ 天野 洋一・ 小林 聡・ 西村 努・ 中道 浩司・ 荒木 和哉・ 谷藤 健・ 田引 正・ 三上 浩輝ら(2007)秋まきコムギ新品種「きたほなみ」の育成について.北海道立農試集報 91: 1–13.