Abstract
Sheath blight is considered the most significant disease of rice and causes enormous yield losses over the world. Breeding for resistant varieties is the only viable option to combat the disease efficiently. Seventeen diverged rice genotypes along with 17 QTL-linked SSR markers were evaluated under greenhouse conditions. Pearson’s correlation showed only the flag leaf angle had a significant correlation with sheath blight resistance under greenhouse screening. Multivariate analysis based on UPGMA clustering and principal component analysis (PCA) indicated that the flag leaf angle, flag leaf length, and plant compactness were significantly associated with the following SSR marker alleles: RM209 (116,130), RM202 (176), RM224 (126), RM257 (156), RM426 (175), and RM6971 (196), which are linked to the SB QTLs: QRlh11, qSBR11-3, qSBR11-1, qSBR9-1, qShB3-2, and qSB-9. A Mantel test suggested a weak relationship between the observed phenotypes and allelic variation patterns, implying the independent nature of morphological and molecular variations. Teqing and Tetep were found to be the most resistant cultivars. IR65482-4-136-2-2, MR219-4, and MR264 showed improved resistance potentials. These results suggest that the morphological traits and QTLs which have been found to associate with sheath blight resistance are a good choice to enhance resistance through pyramiding either 2 QTLs or QTLs and traits in susceptible rice cultivars.
Introduction
Sheath blight (SB) is a fungal disease of rice caused by a necrotrophic soil-borne fungus Rhizoctonia solani (teleomorph: Thanaptephorus cucumeris (Frank) Donk) which was first identified as a parasite of potato in 1898 by Kuhn (Almasia et al. 2008). SB in rice was first reported in 1910 in Japan (Ou 1985). This disease can cause up to 50% yield losses under favorable environmental conditions (Marchetti and Bollich 1991, Rush and Lee 1992). It has been reported that in China 15 to 20 million hectares of rice field affected by SB instigates yield losses of 6 million tons every year (Chen et al. 2012). In 2012, SB affected about 491,932 ha of rice in Japan (JPPA 2013). In the USA, crop losses due to rice SB have been recorded as up to 50% in susceptible cultivars (Prasad and Eizenga 2008). A crop loss of up to 40% has been recorded in Bangladesh (Shahjahan et al. 1986). These losses could be reduced by application of chemical fungicides, but these are of limited utility, primarily due to problems related to timing and cost of application, weather dependencies, and a potential damaging environmental impact (Mew et al. 2004, Tang et al. 2002, Xie et al. 2008). Due to these circumstances, development and use of resistant genotypes may be a highly effective method to control the disease.
This disease is characterized by infection of plants at the late tillering stage, especially at internode elongation time. Accordingly, the disease develops an oval-shaped lesion appearing on the leaf sheath at the water line, often at the junction of the leaf and sheath at the early stage. Gradually, lesions turn pale green to off-white with a narrow purple-brown to brown border. At the severe stage of the disease, lesions amass and the disease moves upward, leaves are attacked, and irregular spots develop. SB resistance in rice might be due to variations in morphological and ecological characters (Groth and Nowick 1992) such as plant height (Li et al. 1995a, Pinson et al. 2005, Zou et al. 2000), days to heading (Pan et al. 1999, Pinson et al. 2005, Zou et al. 2000), tiller angle (Pinson et al. 2005, Zou et al. 2000, Zuo et al. 2014), plant compactness (Pinson et al. 2005, Zou et al. 2000), width of flag leaf and leaf (Pinson et al. 2005, Zou et al. 2000), and leaf morphology (Pinson et al. 2005, Zou et al. 2000). Environmental factors, such as sunlight, humidity, temperature (Rush and Lee 1992), soil fertility (Rush and Lee 1992), and silicon level in the soil (Rodrigues et al. 2003), influence plant growth which indirectly affects disease infestation. As such, correlations between morphological traits and SB resistance have been frequently observed in many findings (Channamalikajjuna et al. 2010, Groth and Nowick 1992, Li et al. 1995a).
Besides morphological traits, more than 60 quantitative trait loci (QTLs) have been found to have links with SB resistance (Zeng et al. 2011). Among the identified QTLs, a few are independent of plant morphology, for example, the QTLs qSB-9 and qSB9-1, which have been mapped on chromosome 9, and qSBR11-1, which has been mapped on chromosome 11, are linked to the SSR (simple sequence repeat) markers RM205 (Han et al. 2002), RM257, and RM224 (Channamallikarjuna et al. 2010), respectively. Moreover, Taguchi-Shiobara et al. (2013) reported that the QTL qSBR-9 showed a stable resistance reaction to SB under 3 years of field trials of backcross introgression lines (BILs). The QTLs qSB-9 (Han et al. 2002) and qSB3-2 (Liu et al. 2009) were also mapped on chromosome 9 and 3 in Teqing and in Jasmine85, respectively, which are linked to the SSR markers RM205 and RM426, respectively. These findings indicate that there is a potential link between SSR markers and sheath blight resistance QTLs in rice. Complete resistance through introgression of these QTLs alone has not yet been reported. Very recently, the tiller angle gene TAC1 has been pyramided with qSB-9TQ and a sufficient quantity of positive influence on SB resistance has been recorded (Zuo et al. 2014). This report indicates that pyramiding of QTLs and morphological traits could indeed bring added value to SB resistance. Moreover, the selected QTLs showed higher additive effects suggesting the possible pyramiding of 2 or more QTLs in a single genotype might be a strategy to improve SB resistance in rice.
Nevertheless, identification of SB resistance becomes challenging because disease pathogenicity is highly influenced by physiological traits, variation of disease pressure with location and years, lack of appropriate method to evaluate the disease precisely, variations in the rating system among scientists, variations in experimental conditions in various studies, lack of resistant adapted germplasm, the limited efficiency and effectiveness of available screening methods, and the polygenic nature of the resistance phenotype (Gonzalez et al. 2006). Hence, the multitude of factors influencing SB epidemics makes the evaluation of the disease incidence difficult. This is why developing SB resistant genotypes through conventional breeding has reached only limited success (Jia et al. 2007). Previous studies on SB resistance indicated that there are no rice varieties or lines with resistance to this disease in rice landraces (Jia et al. 2007, Liu et al. 2009, Zeng et al. 2011). Hence, detailed study on the association of QTLs with morphological traits which are controlling SB resistance necessitates this study to find out the key responsible elements for quantitative SB resistance for pyramiding them in a single genotype to further improve resistance.
Therefore, the present study was undertaken to: i) identify and validate the reported QTL effects on local R. solani field isolates and discover their association with morphological traits, ii) identify desirable morphological traits and SSR marker alleles which contribute to SB resistance, iii) discover new SB resistance sources against local R. solani field isolates in local genotypes.
Materials and Methods
Plant materials
A total of 17 rice genotypes (Table 1) were selected from diverse geographic origin based on their importance regarding SB resistance potentialities, high-yielding attributes, and sources of resistance elements against other major diseases. Six of the 17 genotypes, Teqing, Tetep, Jasmine85, Pecos, Azucena, and Taducan, were selected based on previous reports for their moderate resistance capabilities to SB. Teqing was reported as a major source of SB resistance having QTLs such as qSB-9 and qSB-9-2 (Pan et al. 1999, Pinson et al. 2005) as well as QRlh6, QRlh7a, QRlh7b, and QRlh11 (Xie et al. 2008). Tetep was found to have substantial resistance potential with SB QTLs like qSBR7-1, qSBR9-1, qSBR11-1, qSBR11-2, and qSBR11-3 (Channamallikarjuna et al. 2010). Moreover, the QTL qSBR-11-1 was also fine mapped to the 0.85 Mp region on chromosome 11 (Channamallikarjuna et al. 2010). Sato et al. (2004) reported that the rice line WSS2, derived from Tetep, displays a highly partial resistance to sheath blight. On the other hand, the QTLs qShB3-2, qShB-9-1, and qShB-9-2 conferring resistance to SB were also mapped on chromosome 9 of Jasmine85 (Liu et al. 2009). Resistance QTLs to SB in Pecos (Bollich et al. 1985, Sharma et al. 2009) were also reported. However, the three cultivars Teqing, Tetep, and Jasmine85 were used in our experiment as standard QTL donors by which we can make comparisons with our unknown genotypes and predict the resistance potentialities based on the presence or absence of these resistance QTLs with validation under greenhouse screening. On the other hand, MR219 is a high-yielding mega variety of Malaysia, whereas BRRIdhan28, BRRIdhan29, and MR11 are high-yielding mega varieties of Bangladesh. UKMRC2, 9, and 11 are newly developed high-yielding varieties in Malaysia. The remaining materials are also promising with high-yield potentials. Hence, the main objective of selecting these 17 materials was to find out resistant donors and susceptible recipients against local R. solani field isolates from previously reported sources and local unknown popular varieties, respectively.
Table 1
Basic information on the 17 rice genotypes used in this study
| SL No. |
Genotype |
Source |
Origin |
| 1 |
Teqing |
IRRI |
China |
| 2 |
Jasmine85 |
IRRI |
USA |
| 3 |
Tetep |
IRRI |
Vietnam |
| 4 |
Azucena |
IRRI |
Philippines |
| 5 |
Taducan |
IRRI |
unknown |
| 6 |
MR264 |
MARDI |
Malaysia |
| 7 |
UKMRC2 |
UKM |
Malaysia |
| 8 |
UKMRC3 |
UKM |
Malaysia |
| 9 |
UKMRC9 |
UKM |
Malaysia |
| 10 |
UKMRC11 |
UKM |
Malaysia |
| 11 |
MR219 |
MARDI |
Malaysia |
| 12 |
MR219-4 |
MNA |
Malaysia |
| 13 |
MR219-9 |
MNA |
Malaysia |
| 14 |
IR65482-4-136-2-2 |
IRRI |
Philippines |
| 15 |
BR11 |
BRRI |
Bangladesh |
| 16 |
BRRIdhan28 |
BRRI |
Bangladesh |
| 17 |
BRRIdhan29 |
BRRI |
Bangladesh |
IRRI = International Rice Research Institute, MARDI = Malaysian Agricultural Research and Development Institute, UKM = Universiti Kebangsaan Malaysia, MNA = Malaysian Nuclear Agency, BRRI = Bangladesh Rice Research Institute.
Seeds of 17 rice genotypes were germinated on moist filter paper in petri dishes. Fifty seeds of each entry were treated with Bavistin DF (Carbendazim 0.5% w/w, BASF, Germany) for 24 h to control seed-borne pathogens. After treatment, seeds were thoroughly cleaned with distilled water several times and put on moist filter paper in the petri dishes. These petri dishes with about 0.5 cm water were placed on a lab bench at 30°C, and incubated at 12 h dark and 12 h light conditions for 7 days. Every day, seed germination and seedling growth were monitored. A necessary amount of water was added to the petri dishes for normal growth of seedlings.
Transplanting of seedlings
A ratio of 4:1 of top soil and cow-dung compost were filled into an autoclave bag. Soil was autoclaved at 121°C and 1.05 kgcm−2 for 15 min. A total of 51 plastic pots (15-cm-diameter) were filled with steam-sterilized soil, which ensured the absence of soil-borne R. solani inocula. To decompose soil, pots were soaked with water for 7 days. Fifty mg of Furadan 3G (Carbofuran 3% w/w) insecticide was used per pot during pot preparation to control soil-borne nematodes. Five 7-day-old seedlings of each entry were transplanted into a prepared pot 10 cm apart.
Application of fertilizers
Urea (N), triple super phosphate (P2O5), and potassium chloride (K2O) were applied at 850 mg, 650 mg, and 575 mg per pot, respectively. Fertilizers were applied with 4 equal portions during pot preparation, 21 days and 42 days after planting, and 1 day before inoculation. Plants were grown on the greenhouse bench (Fig. 1A) until inoculation.
Artificial plant inoculation
Preparation of fungal inoculums
Collected diseased plant tissue of rice stems were dipped in 70% ethanol and cut into 1.5- to 2.0-cm pieces. The pieces were then surface-sterilized with 10% Clorox solution for 30 s. The tissues were cultured on rose bengal agar (RBA) to facilitate fungal growth while inhibiting bacterial contaminants. After 1 week of incubation in the dark at 28°C, the petri dishes were examined for R. solani growth, which was then confirmed by observation under an Olympus TH4-200 compound microscope (Olympus, Australia). Verified isolates of R. solani were sub-cultured by transferring a 5-to 7-mm RBA block onto PDA containing tetracycline (0.005%, w/v). Culture media were incubated in the dark for 3 days at 28°C. Single tips of mycelia were then transferred onto fresh PDA and maintained at 27 to 30°C (Jia et al. 2007).
Pathogen inoculation
Three tillers of each plant, i.e. 9 tillers in each pot per replication were inoculated, at 60 days old. Five- to 7-mm PDA blocks were placed at the base of each stem and covered with aluminum foil (Fig. 1B). A single pot with 3 plants of each genotype was used as a control. Pots were arranged following the completely randomized design (CRD), with 3 replications. The aluminum foil was removed after 3–5 days of inoculation when disease lesions appeared. The inoculated plants were kept for 2 weeks inside the walk-in humidity chamber (Fig. 1C), and returned to the greenhouse bench for 1 week to maintain the normal growth conditions of the plants. Pots were again placed in the walk-in humidity chamber for another week to provide favorable conditions for infection. The average day-night temperatures and relative humidity in the screening chambers were maintained from 28 to 34°C and >80%, respectively. The average temperature and humidity in the greenhouse was 35°C and 85%, respectively, during experimental periods.
Disease evaluation
Plants were evaluated after 4 weeks of inoculation. Relative lesion height (RLH) based disease scores were used to assess SB severity. RLL was calculated using the following formula: (lesion length/plant height)*100 (Sharma et al. 1990). RLH was then converted into the SB disease index based on a 0 to 9 scale, where 0 = no lesion and 9 = lesion visible up to the tip of the longest leaf (Prasad and Eizenga 2008). Visual ratings (VRT) were also assigned to the infected plants based on the actual visible damage of the plant tissues. VRTs were also scored according to the SB disease index on a 0 to 9 scale, where 0 = no lesion, 9 = complete damage of the plant, and <4 was considered moderately resistant (Jia et al. 2007, Prasad and Eizenga 2008).
Data recording on morphological traits
Data from the following parameters were collected according to the guidelines described in standard evaluation systems for rice (IRRI 2002).
Plant height (PHT)
The average height of 5 plants from the ground level to the tip of the main panicle was measured in centimeters (cm) at maturity.
Days to heading (DTH)
The number of days required for the plants to show panicle emergence or blooming from the date of sowing.
Flag leaf length (FLL)
Flag leaf length was measured from the collar joint to the tip of a flag leaf in centimeters using a ruler.
Flag leaf width (FLW)
Flag leaf width was measured from the middle across the width of a flag leaf in centimeters using a ruler.
Flag leaf angle (FLA)
Flag leaf angles were recorded from the angle produced between the stem and flag leaf using a protractor.
Second leaf length (SLL)
Second leaf length was measured from the collar joint to the tip of a flag leaf in centimeters using a ruler.
Second leaf width (SLW)
Second leaf width was measured from the middle across the width of a flag leaf in centimeters using a ruler.
Second leaf angle (SLA)
Second leaf angles were recorded from the angle produced between the stem and flag leaf using a protractor.
Tiller number per hill (TNH)
The number of tillers was counted from the primary and secondary culms of a hill.
Plant compactness (PCP)
Plant compactness was calculated according to the following formula: PCP = (total number of tillers per plant)/(sum of the total angles among the tillers of a hill).
SSR marker genotyping
A total of 17 SSR markers were selected from the previously reported 60 QTLs conferring resistance to SB. Points were considered in selection of markers especially on consistencies of effectiveness of QTLs which were frequently authenticated and validated, LOD scores (>3.0), percent phenotypic variances, and successful uses in previous breeding programs.
Isolation of total DNA
Leaf samples were collected from 40-day-old rice plants. Fifteen- to 20-cm-long second leaves were cut from 5 plants and put into a plastic bag. An ice box was used to collect leaf samples. Leaf samples were stored at −20°C for immediate further use. DNA was extracted following the step-by-step procedures using Qiagen DNeasy mini kit (Qiagen, Germany) as described in the manufacturer’s guidelines. Two μL of DNA was pipetted on the peddle stand of the Nanodrop-ND1000 spectrophotometer system (Thermo Scientific, USA). Absorption at 260/280 and 260/230 nm wavelengths was recorded. The purity of the extracted DNA was also checked on 1% agarose gel.
Polymerase chain reaction (PCR)
Each polymerase chain reaction (PCR) was conducted in 10 μL reaction volume containing 1.0 μL 10x reaction buffer (10 mM) (MgCl2 included), 0.2 μl dNTPs (200 μM), 0.2 μL Taq polymerase (1 U/L), 1 μL of each primer (5 pM), 1 1 μLof template DNA (10 ng), and 5.6 μL of ddH2O. The following temperature profiles and cycles were maintained using Eppendorf Nexus Gradient Thermocycler: 1 cycle at 94°C for 4 min, followed by 35 cycles of at 94°C for 45 s, annealing at 55°C or 61°C (depending on primers) for 45 s, 1 cycle at 72°C for 1 min, a final extension at 72°C for 10 min, and storage at 10°C. A list of primers is given in Table 2.
Table 2
Detailed information on alleles detected in rice genotypes for 17 SSR markers and their linked SB QTLs
| SSR marker |
Alleles detected (bp) |
Number of genotypes for the allele detected |
Rice genotypes for the allele detected |
QTLs associated with SB resistance |
Mapping population |
Reference |
| RM209 |
110 |
8 |
MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, IR65582-4-136-2-2, BR11 |
QRh11, qSBR11-2 |
Teqing/Lemont, Tetep/HP2218 |
Liu et al. 2009
Channamalikarjuna et al. 2010 |
|
116 |
3 |
Teqing, Jasmine85, Azucena |
|
|
|
|
125 |
5 |
Taducan, MR219-4, MR219-9, BRRIdhan28, BRRIdhan29 |
|
|
|
|
130 |
1 |
Tetep |
|
|
|
| RM245 |
110 |
1 |
Tetep |
qShB9-2 |
Jasmine85/Lemont |
Liu et al. 2009 |
|
116 |
6 |
Azucena, MR219-4, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
qSB9-2 |
Teqing/Lemont |
Wang et al. 2011 |
|
125 |
10 |
Teqing, Jasmine85, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-9 |
|
|
|
| RM257 |
136 |
4 |
Tetep, Taducan, MR219-4, MR219-9 |
qSBR9-1 |
Tetep/HP2218 |
Channamalikarjuna et al. 2010 |
|
141 |
12 |
Teqing, Jasmine85, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
|
156 |
1 |
Azucena |
|
|
|
| RM434 |
144 |
14 |
Jasmine85, Tetep, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
qShB9-1 |
Jasmine85/Lemont |
Liu et al. 2009 |
|
151 |
3 |
Teqing, UKMRC2, UKMRC3 |
|
|
|
| RM439 |
246 |
16 |
Teqing, Jasmine85, Tetep, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
QRlh6 |
Teqing/Lemont |
Xie et al. 2008 |
|
282 |
1 |
Azucena |
|
|
|
| RM3691 |
112 |
1 |
BRRIdhan28 |
qSBR7-1 |
Tetep/HP2218 |
Channamalikarjuna et al. 2010 |
|
124 |
2 |
Azucena, UKMRC3 |
|
|
|
|
130 |
14 |
Teqing, Jasmine85, Tetep, Taducan, MR264, UKMRC2, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan29 |
|
|
|
| RM1339 |
122 |
2 |
Tetep, Taducan |
– |
Pecos/Lemont |
Sharma et al. 2009 |
|
134 |
14 |
Teqing, Jasmine85, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
|
148 |
1 |
Azucena |
|
|
|
| RM478 |
234 |
8 |
Teqing, Tetep, Azucena, Taducan, MR219-4, MR219-9, UKMRC9, IR65582-4-136-2-2 |
QRlh7b |
Teqing/Lemont |
Xie et al. 2008 |
|
253 |
9 |
MR264, UKMRC2, UKMRC3, UKMRC11, MR219, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
| RM205 |
102 |
8 |
MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9 |
qSB9 |
Teqing/Lemont |
Xie et al. 2008 |
|
114 |
9 |
Teqing, Jasmine85, Tetep, Taducan, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
| RM3685 |
106 |
10 |
Teqing, Jasmine85, Azucena, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219 |
– |
Pecos/Lemont |
Sharma et al. 2009 |
|
112 |
7 |
Tetep, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
| RM6971 |
176 |
16 |
Teqing, Jasmine85, Tetep, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
qSB-9 |
Teqing/Lemont |
Han et al. 2002 |
|
196 |
1 |
Azucena |
|
|
|
| RM182 |
392 |
13 |
Jasmine85, Tetep, Azucena MR264, UKMRC2, UKMRC3, UKMRC9, MR219-4, MR219-9, BR11, BRRIdhan28, BRRIdhan29 |
QRlh7a |
Teqing/Lemont |
Xie et al. 2008 |
|
412 |
1 |
Taducan |
|
|
|
|
436 |
3 |
Teqing, UKMRC11, MR219 |
|
|
|
| RM201 |
182 |
1 |
Azucena |
qSB-9 |
Teqing/Lemont |
Han et al. 2002 |
|
196 |
16 |
Teqing, Jasmine85, Tetep, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
| RM202 |
176 |
2 |
Tetep, Taducan |
QRlh11 |
Teqing/Lemont |
Xie et al. 2008 |
|
204 |
7 |
Teqing, Azucena, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11 |
qSBR11-3 |
Tetep/HP2218 |
Channamalikarjuna et al. 2010 |
|
216 |
1 |
Jasmine85 |
|
|
|
|
220 |
7 |
MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
| RM224 |
126 |
2 |
Tetep, Taducan |
qSBR11-1 |
Tetep/HP2218 |
Channamalikarjuna et al. 2010 |
|
142 |
6 |
MR219-4, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
|
|
|
|
150 |
9 |
Teqing, Jasmine85, Azucena, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219 |
|
|
|
| RM215 |
156 |
1 |
MR219-4 |
qShB9-2 |
Jasmine85/Lemont |
Liu et al. 2009 |
|
168 |
14 |
Teqing, Azucena, Taducan, MR264, UKMRC2, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-9, IR65582-4-136-2-2, BR11, BRRIdhan28, BRRIdhan29 |
qSB9-2 |
Teqing/Lemont |
Wang et al. 2011 |
|
174 |
2 |
Jasmine85, Tetep |
|
|
|
| RM426 |
156 |
1 |
Azucena |
qShB3-2 |
Jasmine85/Lemont |
Liu et al. 2009 |
|
175 |
4 |
Taducan, MR264, BRRIdhan28, BRRIdhan29 |
|
|
|
|
190 |
12 |
Teqing, Jasmine85, Tetep, Azucena, UKMRC3, UKMRC9, UKMRC11, MR219, MR219-4, MR219-9, IR65582-4-136-2-2, BR11 |
|
|
|
|
200 |
1 |
UKMRC2 |
|
|
|
Fifty mL of 8% polyacrylamide gel was prepared according to the procedures described by Sambrook et al. (1989). CBS mini vertical gel electrophoresis system (CBS Scientific, USA) was used to run polyacrylamide gel electrophoresis (PAGE). After completion of the electrophoresis, the gel was removed from the glass plates very carefully to avoid any tears in the gel. DNA was stained by soaking the gel in Sybr-safe DNA gel-stain solution (10 uL/100 mL H2O) for 15 to 20 min. Subsequently, gels were visualized under UV transilluminator and images were documented using the Alphaimager gel documentation system (Alpha Innotech, USA).
Allele scoring of 17 SSR genotypes
Alleles of 17 SSR markers were genotyped based on 2 scoring systems using the presence or absence of respective alleles. PCR bands correspond to the size of the base-pair (bp). Bands’ sizes were estimated using AlphaEase software (Alpha Inotech, USA). The band sizes were categorized into different groups with 5 bp intervals. The bands which differed at least 5 bp from each other were taken into separate groups. Then the bands in the different groups were scored as ‘A/A’, ‘B/B’, ‘C/C’ or ‘D/D’ across the whole entries. Missing bands were scored as ‘0/0’. The markers’ alleles were also scored as ‘1’ or ‘0’ based on the presence or absence of the respective allele, and missing bands were scored as ‘9’.
Statistical analysis
Disease evaluation data recorded from 9 individual plants (tillers) were averaged as a single replication from mist-chamber screening. Morphological data were measured from 5 individual plants for each replication. RLL and VRT scores were recorded from 9 individuals and averaged into a single replication. Data of each of 3 replications were subjected to statistical analysis. Analysis of variance (ANOVA) and least significance differences (LSD) were calculated using the ANOVA procedure using SAS version 9.1 for Windows (SAS, Cary, NC). Simple Pearson’s correlation coefficients were estimated between morphological traits and sheath blight disease scores. Standardized data of morphological traits were used to calculate the Euclidean distance coefficient (Supplemental Table 1) using SIMINT program (NTSYS pc version 2.21q, Rohlf 2008). For SSR markers, each genotype was scored manually as ‘A/A’, ‘B/B’, ‘C/C’, ‘D/D’, or ‘0/0’for missing data. Genetic distance (Supplemental Table 2) was estimated based on Nei and Chesser (1983) using PowerMarker v4.3 (Liu 2003). SIMQUAL program of NTSYS-pc version2.21q was used to calculate allelic distances among the genotypes. Cluster analysis was performed separately for morphological and molecular data. UPGMA clustering was performed based on Euclidean distance and the Sequential Agglomerative Hierarchical and Nested (SAHN) method (Sneath and Sokal 1973). Principal component analysis (PCA) and scattered plot diagrams were also performed for both morphological and molecular data using Multivariate Statistical Software Package (MVSP) version 3.1 (Kovach 2007). To understand the association between SSR marker alleles and morphological traits, Mantel test (Mantel 1967) was performed using both morphological and molecular distance matrices (Supplemental Tables 1, 2) at 250 permutations followed by the determination of a significant level of correlation (MXCOMP program of NTSYS-pc, Rohlf 2000).
Results
Analysis of variance and mean performances
Analysis of variance (mean sum of squares) and related statistics for 11 morphological traits of 17 rice genotypes are presented in Table 3. Results indicate that all the genotypes showed highly significant differences from each other (p < F, 0.0214 to 0.0001). The mean performance of the morphological traits (Table 4) revealed that UKMRC9 was the latest maturing genotype. IR65482-4-136-2-2 had the lowest growth duration, and ‘Tetep’ possessed the highest plant height followed by ‘Azucena’. ‘Jasmine 85’ was the shortest cultivar. The longest flag leaf was recorded in MR219-4, whereas BR11 had the shortest one. The widest flag leaf was observed in BRRIdhan29 followed by MR219-9, whereas the narrowest was in ‘Tetep’ and ‘Taducan’. ‘Teqing’ had the most erect flag leaf, but ‘Azucena’ produced the droopiest flag leaves followed by UKMRC2. MR219-9 had the longest second leaves, whereas the shortest second leaves were found in MR264 next to BR11. The widest second leaves were possessed by MR219-9, but the narrowest by ‘Tetep’. ‘Teqing’, MR264, and UKMRC9 produced the most erect second leaves, whereas the droopiest second leaves were in BR11 next to BRRIdhan28. The most compact tillering patterns were observed in UKMRC11 followed by MR219-4 and BR11. The most highly spreading type plants were recorded from IR65482-4-136-2-2. The lowest relative lesion height (RLH) was observed in ‘Tetep’ followed by IR65482-4-136-2-2, MR264, MR219-4, Teqing, and UKMRC11. The highest RLH was recorded from UKMRC2. The most resistant cultivars, Teqing (Fig. 2A) and Tetep (Fig. 2B), had the lowest VRT (3.5). UKMRC2 (Fig. 2C) was the most susceptible cultivar among the 17 tested genotypes.
Table 3
Analysis of variance (ANOVA) for 11 morphological traits of 17 rice genotypes
| Source of variation |
DTHa |
PHTb |
FLLc |
FLWd |
FLAe |
SLLf |
SLWg |
SLAh |
PCPi |
RLHj |
VRTk |
| Genotype |
336.00 |
651.08 |
151.79 |
0.282 |
1355.24 |
504.69 |
0.146 |
515.10 |
31.15 |
0.884 |
2.70 |
| Error |
155.6 |
14.78 |
8.837 |
0.0018 |
8.83 |
3.41 |
0.0013 |
15.39 |
5.03 |
0.18 |
0.65 |
| F |
2.16 |
44.04 |
17.18 |
150.27 |
153.48 |
147.89 |
105.9 |
33.48 |
6.19 |
4.89 |
4.16 |
| p < F |
0.020 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
| LSD |
20.62 |
6.36 |
4.91 |
0.072 |
4.91 |
3.05 |
0.06 |
6.48 |
3.71 |
0.70 |
1.33 |
| Mean |
99.57 |
112.77 |
40.34 |
1.655 |
41.76 |
52.52 |
1.42 |
46.72 |
6.92 |
4.10 |
5.03 |
| % CV |
17.25 |
7.23 |
0.41 |
22.22 |
12.74 |
0.25 |
13.67 |
2.48 |
0.68 |
1.04 |
10.29 |
j Relative lesion height,
Table 4
Mean performances and standard deviations from the mean of 11 morphological traits of 17 rice genotypes
| Genotype |
DTHa |
PHTb |
FLLc |
FLWd |
FLAe |
SLLf |
SLWg |
SLAh |
PCPi |
RLHj |
VRTk |
| Teqing |
79.0 ± 1.0 cd |
110.6 ± 2.9 de |
47.3 ± 0.9 b |
1.7 ± 0.0 d |
15.3 ± 2.3 h |
50.2 ± 3.0 e |
1.4 ± 0.0 g |
30.0 ± 5.3 h |
7.1 ± 3.2 ef |
3.97 ± 0.0 cde |
3.5 ± 1.0 d |
| Jasmine85 |
97.7 ± 1.5 abc |
96.8 ± 1.1 k |
38.5 ± 0.8 efg |
1.6 ± 0.0 g |
57.3 ± 2.3 c |
50.1 ± 0.3 e |
1.4 ± 0.0 fg |
59.3 ± 2.3 b |
4.9 ± 2.1 de |
4.34 ± 0.6 bcde |
4.7 ± 1.0 bcd |
| Tetep |
96.7 ± 2.5 abcd |
146.0 ± 1.1 a |
42.2 ± 8.3 bcde |
1.0 ± 0.0 i |
34.7 ± 1.2 e |
63.7 ± 2.8 b |
1.0 ± 0.0 i |
55.3 ± 3.1 bc |
7.5 ± 0.7 b |
3.03 ± 0.0 f |
3.5 ± 1.2 d |
| Azucena |
107.7 ± 1.5 ab |
140.6 ± 3.0 b |
42.8 ± 1.7 cdef |
1.9 ± 0.1 c |
85.7 ± 3.8 a |
57.7 ± 2.3 cd |
1.8 ± 0.0 a |
42.7 ± 6.1 ef |
10.2 ± 1.9 cd |
4.47 ± 0.6 bc |
6.5 ± 0.6 a |
| Taducan |
111.3 ± 1.5 ab |
145.8 ± 3.8 a |
44.2 ± 2.9 bcd |
1.0 ± 0.0 i |
38.0 ± 1.7 e |
58.4 ± 1.4 cd |
1.0 ± 0.1 i |
55.3 ± 2.3 bc |
4.3 ± 1.9 fg |
4.39 ± 0.6 bcd |
5.2 ± 0.6 abc |
| MR264 |
107.3 ± 1.0 ab |
99.6 ± 1.0 ijk |
40.1 ± 4.1 defg |
1.6 ± 0.0 ef |
16.7 ± 1.2 h |
31.1 ± 1.0 i |
1.5 ± 0.1 e |
30.0 ± 2.3 bc |
11.5 ± 5.3 c |
3.63 ± 0.6 ef |
4.7 ± 0.6 bcd |
| UKMRC2 |
105.0 ± 1.0 ab |
107.1 ± 7.9 efg |
37.7 ± 1.2 efg |
1.5 ± 0.0 h |
84.7 ± 1.2 a |
42.7 ± 0.7 gf |
1.4 ± 0.0 ef |
44.7 ± 2.0 h |
11.0 ± 2.1 cd |
5.60.1 ± 0.6 a |
6.3 ± 1.5 a |
| UKMRC3 |
96.0 ± 1.0 abcd |
105.5 ± 2.0 fgh |
37.3 ± 1.3 fg |
1.5 ± 0.0 h |
28.0 ± 3.1 fg |
38.4 ± 0.8 h |
1.3 ± 0.0 g |
49.3 ± 1.2 ef |
5.8 ± 2.1 efg |
3.86 ± 0.6 cde |
5.3 ± 1.0 abc |
| UKMRC9 |
115.0 ± 1.2 a |
109.0 ± 0.7 |
def 35.2 ± 5.9 gh |
1.6 ± 0.0 e |
27.3 ± 2.3 g |
39.8 ± 0.5 gh |
1.6 ± 0.1 bc |
30.0 ± 0.0 de |
6.1 ± 0.4 efg |
4.20 ± 0.6 cde |
5.2 ± 1.0 abc |
| UKMRC11 |
111.0 ± 2.0 ab |
117.9 ± 6.4 c |
40.4 ± 2.4 def |
1.7 ± 0.0 d |
33.2 ± 2.0 ef |
59.5 ± 2.3 cd |
1.5 ± 0.0 d |
45.3 ± 5.0 h |
2.6 ± 0.7 g |
3.98 ± 0.6 cde |
5.7 ± 1.5 ab |
| MR219 |
111.0 ± 1.0 ab |
116.9 ± 1.2 c |
31.7 ± 1.9 h |
1.7 ± 0.0 d |
28.7 ± 1.4 fg |
43.7 ± 0.9 e |
1.6 ± 0.0 c |
53.3 ± 5.0 cd |
4.7 ± 0.6 g |
4.92 ± 0.6 ab |
5.7 ± 0.0 ab |
| MR219-4 |
98.7 ± 2.1 abc |
113.9 ± 1.3 cd |
59.0 ± 0.9 a |
1.9 ± 0.0 c |
29.3 ± 3.1 fg |
73.3 ± 1.6 a |
1.5 ± 0.1 e |
57.3 ± 2.3 bc |
3.2 ± 0.4 g |
3.67 ± 1.0 def |
4.3 ± 0.6 bcd |
| MR219-9 |
107.7 ± 1.5 ab |
104.2 ± 2.9 fghi |
46.8 ± 2.4 bc |
2.1 ± 0.1 b |
16.7 ± 1.2 h |
74.3 ± 0.5 a |
1.7 ± 0.0 b |
34.7 ± 5.0 gh |
4.7 ± 1.3 g |
4.01 ± 0.6 ef |
4.3 ± 1.2 bcd |
| IR65482-4-136-2-2 |
74.3 ± 0.6 d |
113.7 ± 2.2 cd |
40.1 ± 1.7 defg |
1.5 ± 0.0 h |
35.7 ± 6.8 e |
56.9 ± 3.6 d |
1.6 ± 0.0 bc |
36.0 ± 3.5 g |
14.0 ± 2.5 a |
3.31 ± 1.2 ef |
4.3 ± 0.6 bcd |
| BR11 |
91.3 ± 3.1 bcd |
97.9 ± 2.2 jk |
23.9 ± 1.3 i |
1.8 ± 0.0 e |
62.0 ± 2.0 c |
32.0 ± 1.0 i |
1.3 ± 0.0 g |
71.3 ± 4.9 a |
3.2 ± 1.6 fg |
4.11 ± 1.0 cde |
6.2 ± 0.6 a |
| BRRIdhan28 |
97.7 ± 1.5 abc |
101.9 ± 1. hij |
31.5 ± 1.1 h |
1.4 ± 0.0 j |
68.0 ± 3.5 b |
39.5 ± 1.9 h |
1.2 ± 0.0 h |
70.0 ± 0.0 a |
5.9 ± 1.9 cd |
4.26 ± 0.0 bcde |
4.3 ± 0.6 cd |
| BRRIdhan29 |
102.7 ± 1.5 ab |
103.1 ± 1.3 ghi |
41.2 ± 0.7 def |
2.3 ± 0.1 a |
48.0 ± 6.0 d |
60.2 ± 0.3 e |
1.7 ± 0.0 b |
34.7 ± 3.1 |
8.7 ± 2.6 cd |
4.14 ± 0.0 cde |
4.2 ± 0.6 cd |
| Grand Mean |
99.6 |
112.8 |
40.3 |
1.7 |
41.8 |
52.5 |
1.4 |
46.7 |
6.9 |
4.10.6 |
5.0 |
| SD (±) |
10.6 |
14.7 |
7.1 |
0.3 |
21.3 |
13.0 |
0.2 |
13.1 |
3.2 |
0.60.6 |
0.9 |
j Relative lesion height,
Numbers followed by the same letter/s are not significantly different at p < 0.05 level of probability.

Pearson’s correlation coefficients were estimated among all the 11 morphological traits (Table 5). Only the trait FLA was found to be significantly correlated with both SB scores, RLH (0.550) and VRT (0.553). Signification correlation (0.651) was also detected between RLH and VRT. However, among the morphological traits, significant correlations were also detected between PHT and FLW (−0.492), FLL and SLL (0.769), FLW and SLW (0.862), SLA and SLW (−0.573), and SLA and PCP (−0.521). Days to heading showed a positive correlation with both disease indices.
Table 5
Simple Pearson’s correlation coefficient (r) of 11 morphological traits of 17 rice genotypes
| Character |
DTHa |
PHTb |
FLLc |
FLWd |
FLAe |
SLLf |
SLWg |
SLAh |
PCPi |
RLHj |
VRTk |
| DTH |
1.00 |
0.189 |
−0.043 |
0.064 |
0.058 |
0.014 |
0.148 |
−0.046 |
−0.314 |
0.434 |
0.455 |
| PHT |
|
1.00 |
0.309 |
−0.492* |
0.064 |
0.438 |
−0.299 |
0.055 |
0.030 |
−0.136 |
0.041 |
| FLL |
|
|
1.00 |
0.145 |
−0.353 |
0.769** |
0.139 |
−0.345 |
0.010 |
−0.306 |
−0.456 |
| FLW |
|
|
|
1.00 |
−0.004 |
0.209 |
0.826** |
−0.321 |
−0.050 |
0.130 |
0.087 |
| FLA |
|
|
|
|
1.00 |
−0.178 |
−0.011 |
0.438 |
0.200 |
0.550* |
0.553* |
| SLL |
|
|
|
|
|
1.00 |
0.170 |
−0.128 |
−0.150 |
−0.285 |
−0.378 |
| SLW |
|
|
|
|
|
|
1.00 |
−0.573* |
0.269 |
0.163 |
0.210 |
| SLA |
|
|
|
|
|
|
|
1.00 |
−0.521 |
0.107 |
0.197 |
| PCP |
|
|
|
|
|
|
|
|
1.00 |
−0.054 |
−0.070 |
| RLH |
|
|
|
|
|
|
|
|
|
1.00 |
0.651* |
| VRT |
|
|
|
|
|
|
|
|
|
|
1.00 |
* and ** indicate a significant difference at p < 0.1 and p < 0.05 level of probability, respectively.
j Relative lesion height,
Cluster analysis of 17 genotypes using 11 morphological traits revealed 2 major groups, A-I and B-I. The largest cluster, A-I, was composed of 15 genotypes of diverse geographic origin. The cluster further differentiated into 4 sub-clusters, namely A-I-I, A-I-II, A-I-III, and A-I-IV (Fig. 3). The largest sub-group, A-I-III, had a total of 8 genotypes; most of them are susceptible, including BR11 and MR219. Resistant genotypes, such as Tetep and Teqing, were clustered into 2 different sub-groups, B-I and A-I-IV, respectively. UPGMA clustering produced a low cophenetic correlation score (0.717). The mantel test also showed an insignificant and low relationship (r = 0.253) between morphological and molecular data sets.

However, UPGMA clustering based on 47 marker alleles of 17 SSR loci produced 2 broad sub-groups, A-I and B-I (Fig. 4). Only the genotype Azucena was represented into sub-group A-I. The largest sub-group B-I contained the remaining 16 genotypes. The sub-group B-I has further been discriminated into 3 major sub-sub-groups, namely B-I-I, B-I-II, and B-I-III. The dendrogram indicated that the sub-sub-group B-I-I was composed of 8 genotypes with different levels of SB resistance including the resistant genotypes Teqing and Jasmine85 (sub-branch) and the susceptible genotypes UKMRC2 and MR219. Sub-sub-group B-I-II has been confined with six genotypes of variant environmental origins. Sub-sub-group B-I-III had 2 resistant genotypes, ‘Tetep’ and ‘Taducan’, which were clearly distinguished from the other groups comprising susceptible genotypes.
Principal component analysis (PCA)
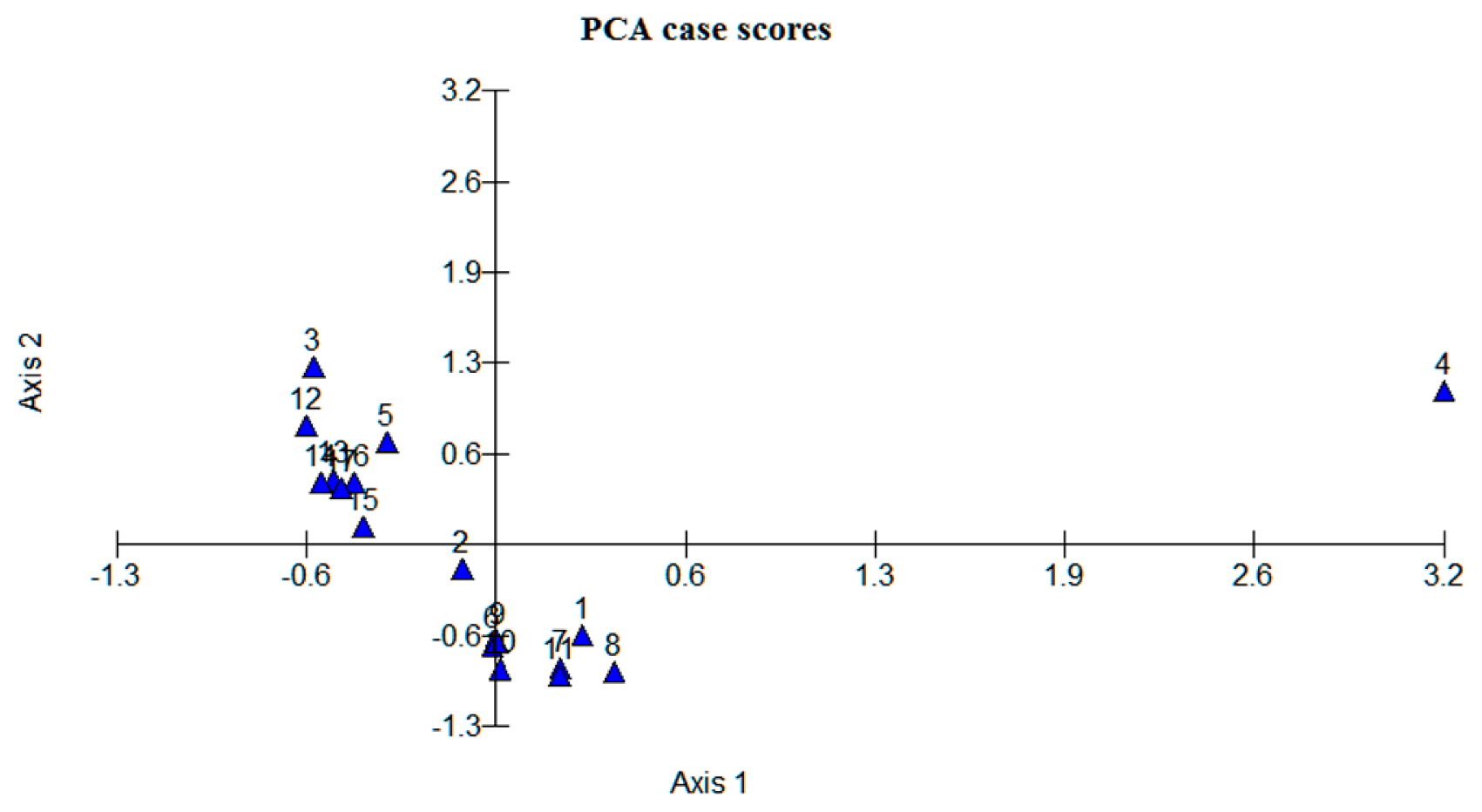
Principal component analysis (PCA), performed using 11 morphological traits, is presented in Table 6. The table showed that all 4 of the principal components (PCs) accounted for the maximum variations (78.73%) explained. The first two PCs accounted for 47.28% of the total variations. PCA variable loadings indicated that FLL (0.328) and PCP (0.332) contributed the maximum in the total variation explained by PC1 (24.58%). In the case of PC2, FLW (0.341), SLW (0.455), VRT (0.404) and RLH (0.356) added the highest values and interpreted 22.70% to the total variations. Similarly, FLL and SLL induced 18.38% and FLW contributed 13.07% to the total variations in PC3 and PC4, respectively. PCA scattered plot diagram (Fig. 5) showed a good separation of the genotypes which was high in agreement with the UPGMA clustering. Scattered plot diagram (Fig. 5) showed significant discrimination of the genotypes into quadrangles. The upper right quadrangle was occupied with 4 genotypes which showed moderate susceptibility. UKMRC2 was located on axis 1, whereas the upper left quadrangles were composed of 3 genotypes, including MR219, susceptible check. BR11 was found on axis 2. The lower left quadrangles contained 4 genotypes UKMRC11, BRRIdhan28, Jasmine85, and Taducan. On the other hand, the lower right quadrangle contained most of the resistant cultivars including ‘Tetep’, ‘Teqing’, IR65482-4-136-2-2, and MR219-4.
Table 6
Principal component analysis of 11 morphological traits of 17 rice genotypes
| Factors |
Axis-1 |
Axis-2 |
Axis-3 |
Axis-4 |
| Eigenvalue |
3.195 |
2.950 |
2.389 |
1.700 |
| Percentage |
24.58 |
22.696 |
18.378 |
13.074 |
| Cum. of percentage |
24.58 |
47.276 |
65.654 |
78.728 |
| PCA variable loadings |
| Days to heading |
−0.171 |
0.238 |
0.290 |
−0.275 |
| Plant height |
0.014 |
−0.195 |
0.037 |
−0.661 |
| Flag leaf length |
0.328 |
−0.134 |
0.331 |
−0.276 |
| Flag leaf width |
0.166 |
0.341 |
0.292 |
0.297 |
| Flag leaf angle |
−0.264 |
0.185 |
−0.278 |
−0.249 |
| Second leaf length |
0.236 |
−0.173 |
0.374 |
−0.303 |
| Second leaf width |
0.228 |
0.455 |
0.110 |
0.080 |
| Second leaf angle |
−0.449 |
−0.236 |
−0.029 |
0.026 |
| Plant compactness |
0.332 |
0.143 |
−0.472 |
−0.094 |
| Visual ratings |
−0.313 |
0.404 |
−0.160 |
0.164 |
| Relative lesion height |
−0.259 |
0.356 |
−0.039 |
−0.145 |
| PCA case scores |
| ’Teqing’ |
0.514 |
−0.216 |
0.029 |
0.214 |
| Jasmine 85 |
−0.368 |
−0.142 |
−0.019 |
0.224 |
| ’Tetep’ |
0.124 |
−1.082 |
−0.244 |
−0.362 |
| ’Azucena’ |
0.130 |
0.604 |
−0.111 |
−0.633 |
| ’Taducan’ |
−0.396 |
−0.421 |
0.198 |
−0.643 |
| MR264 |
−0.266 |
0.671 |
−0.486 |
−0.385 |
| UKMRC2 |
0.006 |
0.437 |
0.099 |
0.063 |
| UKMRC3 |
−0.345 |
0.355 |
0.169 |
0.016 |
| UKMRC9 |
0.468 |
0.172 |
−0.310 |
0.240 |
| UKMRC11 |
−0.095 |
−0.032 |
−0.084 |
0.211 |
| MR219 |
−0.303 |
0.071 |
0.500 |
−0.054 |
| MR219-4 |
0.144 |
−0.295 |
0.708 |
−0.064 |
| MR219-9 |
0.531 |
0.060 |
0.692 |
0.199 |
| IR65482-4-136-2-2 |
0.775 |
−0.177 |
−0.697 |
0.057 |
| BR11 |
−0.866 |
−0.001 |
−0.178 |
0.524 |
| BRRIdhan 28 |
−0.498 |
−0.498 |
−0.353 |
0.234 |
| BRRIdhan 29 |
0.443 |
0.349 |
0.349 |
0.156 |

Another PCA, performed using 47 alleles at 17 SSR marker loci, constructed the principal components (PCs) with 4 major axes based on Eigenvalues which interpreted about 80% of the total variation explained. The results indicated that PC axis 1 possessed the highest Eigenvalues (13.145), followed by 9.515, 6.724, and 4.488 which accounted for 27.97%, 20.25%, 14.31%, and 9.55%, respectively, of the total genetic variations explained (Supplemental Table 3). However, based on the value of PCA variable loadings from the 4 major axes, the following SSR marker alleles: RM209 (116, 130), RM257 (156), RM202 (176), RM224 (126), RM426 (156, 175), RM6971 (196), RM1339 (122, 148), RM439 (282), RM205 (102), RM245 (125), and RM434 (144) were identified as the major contributors to create the observable discrimination among genotypes into different quadrangles. However, a scattered plot diagram (Fig. 6) showed that only one genotype, Azucena, constituted the upper right quadrangle, whereas the upper left quadrangle was constituted of 8 genotypes, including both resistant (Tetep) and susceptible (BR11) cultivars. The lower left quadrangle contained only 1 resistant genotype, Jasmine85. On the other hand, 3 genotypes, MR264, UKMRC9 and UKMRC11, were located on axis 1. The lower right quadrangle constituted 4 genotypes, including both resistant (Teqing) and susceptible (UKMRC2) cultivars (Fig. 6).
Together, the 17 SSR markers produced a total of 47 alleles with different sizes across the 17 rice genotypes investigated in this study (Table 2). Major findings of the allelic variations among the rice genotypes have been described in this section. Marker RM209 produced 4 alleles ranging from 110 to 130 bp in size. Of these 4 alleles, allele 110 was detected in 8 entries. The resistant cultivars Teqing and Jasmine85 produced allele 125, whereas allele 130 was observed in Tetep only for the marker RM209. For the marker RM245, allele 110 was detected in the resistant cultivar Tetep only. Most of the entries produced allele 246 except Azucena (282) for marker 439. For marker RM1339, Tetep and Taducan possessed allele 122 while allele 134 was found in the rest of the cultivars used. In respect of marker RM478, most of the resistant entries, including Teqing and Tetep, possessed allele 234. In the case of marker RM205, allele 114 was observed in most of the cultivars in the resistant group, including Teqing, Jasmine85, Tetep, and Taducan. On the other hand, allele 176 was detected in all the cultivars genotyped for marker RM6971, except Azucena (196). For marker RM201, allele 176 was found only in Azucena, whereas the remaining 16 cultivars harbored allele 196. Four alleles were observed for marker RM202, which ranged from 176 to 220 bp. Resistant cultivars, such as Tetep and Taducan, had allele 176, whereas allele 216 was found in Jasmine85 only. Similarly, allele 126 was detected in Tetep and Taducan only for marker RM224. Marker RM215 produced allele 156 and 174, which were observed in only MR219-4, and in Jasmine85 and Tetep, respectively. Allele 156 and 200 for marker RM426 were only found in Azucena and UKMRC2, respectively.
Discussion
Association between morphological traits and SB resistance
A total of 11 morphological traits of 17 rice genotypes were used to investigate the association between morphological traits and SB resistance. SB resistance in rice may be enhanced due to suitable morphological and ecological parameters (Groth and Nowick 1992) for pathogens, such as reduced solar radiation, high humidity and high temperature (Rush and Lee 1992), and soil compositions (Rodrigues et al. 2003, Rush and Lee 1992), which influence plant growth and indirectly affect disease infestation. Evidence shows that morphological traits, such as plant height, days to heading, tiller angle, plant compactness, flag leaf length and leaf width, impact on SB resistance in rice (Pinson et al. 2005, Zou et al. 2000). However, from our observations, we found that flag leaf angle significantly correlated with SB resistance. The flag leaf in rice plants contributes greatly to rice grain yield. The influences of the flag leaves are controlled by leaf angle. For example, erect leaf morphology captures more light radiation and produces less shade in the plant canopy, especially at the lower part of the plant, than that of droopy phenotypes. Rice SB pathogens favor hot, humid and shady environments for its pathogenesity. This is why SB is generally more predominant at the lower part of the plant than the upper part. Since erect leaf morphology minimizes the shading effect at the lower part of the plant, this is how rice plants with erect flag leaves evade attacks by sheath blight and show low SB infestation. Similar results were also observed by Loan et al. (2004) and Channamallikarjuna et al. (2010). Teqing is a proven resistant cultivar with the lowest flag leaf angle (15.3 ± 2.3), and has been distinguished into a separate cluster (Figs. 3, 4). Principal component-1 accounted for the highest proportion (24.58%) of the total morphological variation which was contributed dominantly by flag leaf length (0.328) and plant compactness to create observable discrimination between resistant and susceptible genotypes. For example, MR219-4 with the highest leaf length (59.0) was distinguished into the lower right quadrangle (Fig. 5), and showed an improved resistance potential under greenhouse screening (Table 4). Thus, an explainable association between morphological traits and SB resistance was proved by our above findings. Morphological traits associated with SB resistance have been studied frequently. Plant height, days to heading, tiller angle, plant compactness, flag leaf length and flag leaf width have been reported as factors influencing resistance in rice plants (Pinson et al. 2005, Zou et al. 2000). Plant height, days to heading, and tiller angle were reported as independent to SB resistance (Srinivasachary et al. 2011). In our study, positive but insignificant correlations between plant height and SB resistance, and between days to heading and SB resistance were detected. However, Li et al. (1995b) mapped 3 SB resistance QTLs, QSbr3a, QSbr8a and QSbr9a, which were at approximately the same locations where the major QTLs, QPh3a, QPh8a and QPh9a, for plant height were mapped (Zou et al. 2000). The QTLs, QHd3a, QHd8a, and QHd9a, controlling days to heading corresponded to QTLs for plant height and SB resistance (Zou et al. 2000). Flag leaf angle was identified to be significantly correlated with SB resistance in our study. Principal component analysis revealed a good association of flag leaf length with SB resistance in our study. The tiller angle gene TAC1 has been identified at the region of qSB-9TQ associated with SB resistance (Zuo et al. 2014).
Marker alleles associated with sheath blight resistance
A total of 47 alleles were detected from 17 SSR markers across the 17 rice genotypes. It has been reported that all the markers were found to have linkage with 19 different putative SB resistance QTLs located on 7 different chromosomes of rice (Table 2). However, our findings indicate that allele 116 of marker RM209 was detected in 3 resistant cultivars, Teqing, Jasmine85, and Azucena (Table 2). Marker RM209 has been reported to be associated with the SB resistance QTL QRlh11, and mapped on chromosome 11 using the Teqing/Lemont population (Xie et al. 2008). The marker also yielded the unique allele 130 which was observed in another resistant cultivar, Tetep. Four resistant cultivars, Tetep, Taducan, MR219-4, and MR219-9, produced allele 136 of marker RM257 which corroborated the finding of Channamallikarjuna et al. (2010), a putative QTL qSBR9-1 identified and mapped on chromosome 9 using the Tetep/HP2219 mapping population. Marker RM478 produced the potential allele 234 which was identified in 8 cultivars. All these cultivars, including Teqing and Tetep, showed resistant reactions under greenhouse screening. Liu et al. (2009) showed that marker RM478 had significant linkage with QTL QRlh7b and was mapped on chromosome 7 using the Teqing/Lemont population. In our study, allele 122 of RM1339 was detected in only 2 resistant cultivars, Tetep and Taducan. Sharma et al. (2009) reported that marker RM1339, linked to SB resistance QTLs, was mapped on chromosome 1 using the Pecos/Rosemont population. On the other hand, alleles 176 and 126 of markers RM202 and RM224, respectively, were detected in Tetep and Taducan in our study. RM202 has been reported as linked to QTL qSBR11-3 (Channamallikarjuna et al. 2010) and QRlh11 (Xie et al. 2008). Similarly, RM224 was also reported by Channamallikarjuna et al. (2010) as a linked marker to the QTL qSBR11-1 which was fine mapped on chromosome 11 in the 0.85 Mb region using the Tetep/HP2219 mapping population. On the other hand, Han et al. (2002) mapped the QTL qSB-9 on chromosome 9 using the Teqing/Lemont population, and reported that marker RM6971 was linked to the QTL. The alleles discussed above were detected mainly in resistant cultivars and not observed in susceptible high-yielding cultivars. These reports clearly indicate that the described alleles are associated with potential resistance effects in the respective cultivars serving as valuable genetic resources to improve SB resistance in high-yielding susceptible genotypes by marker-assisted selection breeding.
Association between morphological traits and SSR marker alleles
Principal component analysis for the 17 morphological traits and 47 alleles of the 17 SSR markers revealed measurable discrimination among the genotypes used in this study. The PCA variable loadings for morphological traits indicated that FLL and PCP contributed the maximum value in the total variation explained by PC1. The longest flag leaf was recorded in MR219-4 which was resistant in our greenhouse screening. On the other hand, morphological traits FLW, SLW, VRT and RLH added the highest values to the total variations explained by PC2. The widest second leaf was possessed by MR219-9 and the narrowest by ‘Tetep’, ‘Teqing’, MR264 and UKMRC9. The most resistant cultivars, Teqing (Fig. 2A) and Tetep (Fig. 2B), had the lowest VRT (3.5). The lowest relative lesion height (RLH) was observed in ‘Tetep’ followed by IR65482-4-136-2-2, MR264, MR219-4, Teqing, and UKMRC11, whereas, the highest RLH was recorded in UKMRC2 (Fig. 2C) the most susceptible cultivar among the 17 tested genotypes. FLL and FLW, and SLL provided considerable contributions to the total variations in PC3 and PC4, respectively. The 47 alleles of the 17 SSR markers indicated a noteworthy partition among the genotypes with the 4 major axes. The following SSR marker alleles: RM209 (116, 130), RM257 (156), RM202 (176), RM224 (126), RM426 (156, 175), RM6971 (196), RM1339 (122, 148), RM439 (282), RM205 (102), RM245 (125), and RM434 (144) were identified as the major contributors to create the observable discrimination among genotypes into the different quadrangles. It can be concluded that the SSR markers that were reported in resistant cultivars were also reported with the same alleles in some improved resistant cultivars from our investigation, implying that the associated morphological traits and SSR markers might generate the potential variations among cultivars.
Association between QTLs and sheath blight resistance
In our study, we analyzed 17 SSR loci linked to 19 QTLs conferring resistance to SB located on different chromosomes (Table 2). More than 60 QTLs (Zeng et al. 2011) were mapped on different genetic backgrounds. To understand the association between QTLs and SB resistance in rice, UPGMA-based cluster analysis was performed using 47 alleles at 17 SSR marker loci. The dendrogram showed a clear-cut separation between resistant genotypes (Tetep and Taducan) and susceptible genotypes (UKMRC2 and MR219). The sub-group A-I contained the resistant genotypes, Azucena (A-I-I). Taducan and Tetep (B-I-III), and Teqing (B-I-I) were significantly separated from susceptible genotypes. The following marker alleles: RM209 (116), RM257 (136), RM478 (234), RM202 (176), RM224 (126), RM 426 (175), and RM6971 (196) possessed the higher values of Eigenvectors (Supplemental Table 3) and these alleles were also detected in resistant cultivars. In addition, it has been reported that the above mentioned SSR markers were found to have linkage with the following SB QTLs: QRlh11 (Xie et al. 2008), qSBR9-1 (Channamallikarjuna et al. 2010), QRlh7b (Liu et al. 2009), qSBR11-3 (Channamallikarjuna et al. 2010), QRlh11 (Xie et al. 2008), qSBR11-1 (Channamallikarjuna et al. 2010), qShB3-2 (Liu et al. 2009), and qSB-9 (Han et al. 2002), suggesting that they contributed significantly to create this measurable variation observed in our study. These results could authenticate that there is some extent of association between genotypes of SSR markers and SB resistance in rice.
Association between morphological traits and QTLs toward SB resistance
The correlation between morphological traits and SB resistance has been frequently observed in many findings (Channamalikajjuna et al. 2010, Groth and Nowick 1992, Li et al. 1995a). More than 60 QTLs including major and minor effects conferring SB resistance (Zeng et al. 2011) have been identified in rice (Table 2). Ten of these 60 QTLs co-localize with QTLs for morphological traits especially, PHT and DTH (Srinivasachary et al. 2011). Three QTLs (qPH-3, qPH-4, and qPH-11) controlling plant height and four QTLs (qHD-2, qHD-3, qHD-5, and qHD-7) for days to heading were mapped (Zou et al. 2000) on different chromosomes in the same mapping population. It has also been reported that the major QTLs QHd3a, QHd8a, and QHd9a controlling days to heading and QTLs QPh3a, QPh8a and QPh9a (Li et al. 1995a) controlling plant height were mapped to approximately the same locations which corresponded to the QTLs QSbr3a, QSbr8a and QSbr9a associated with SB resistance. The tiller angle gene TAC1 has been identified in the region qSB-9TQ associated with SB resistance (Zuo et al. 2014). For both the cases, findings from the analysis of morphological data and molecular data supported each other strongly. Cluster analysis showed the most common pattern of grouping which indicated that the resistant and susceptible genotypes have been grouped in a similar manner such as the ‘Tetep’ and ‘Teqing’ resistant genotypes were grouped into a separate cluster for both morphological data (Fig. 3) and molecular data (Fig. 4). However, not only cluster analysis, but principal component analysis also produced a similar discrimination (Figs. 5, 6) into components (data not shown).
Association analysis using the Mantel (1967) test indicated a weak relationship between the observed morphological traits and allelic variation pattern and detected low congruence between the two datasets produced in this study. Low correlation between morphological and molecular markers has been reported in many crops (Bushehri et al. 2005, Ferriol et al. 2004, Koehler-Santos et al. 2003) and these authors suggested that it could be as a result of the independent nature of morphological and molecular variations.
This suggests that genotypes like Tetep and Teqing might be selected as donors for improvement of SB resistance in rice. IR65482-4-136-2-2, MR219-4, and MR264 were evaluated and QTLs responsible for SB resistance were mapped. Moreover, QTLs controlling plant height (qPH-3, qPH-4 and qPH-11), QTLs controlling days to heading (qHD-2, qHD-3, QHd3a, qHD-5, qHD-7, QHd8a, and QHd9a), and those controlling morphological traits, such as FLA, FLL, and PCP, could be good choices with the following SB QTLs: qSB-9, qSBR9-1, QRlh6, and qShB3-2, to enhance SB resistance through QTL pyramiding or pyramiding QTLs together with any of these morphological traits. Successful pyramiding of the tiller angle gene TAC1 and qSB-9TQ has been reported by Zuo et al. (2014).
Acknowledgements
This work was supported by the Ministry of Higher Education, Malaysia, under the Long-term Research Grant Scheme for Food Security (Project code LRGS/TD/2011/UPM-UKM/KM/01).
Literature Cited
- Almasia, N.I., A.A. Bazzini, H.E. Hopp and C. Vazquez-Rovere (2008) Overexpression of snakin-1 gene enhances resistance to Rhizoctonia solani and Erwinia carotovora in transgenic potato plants. Mol. Plant Pathol. 9: 329–338.
- Bushehri, A., S. Torabi, M. Omidi and M. Ghannadha (2005) Comparison of genetic and morphological distance with heterosis with RAPD markers in hybrids of barley. Int. J. Agric. Biol. 7: 592–595.
- Channamallikarjuna, V., H. Sonah, M. Prasad, G.J.N. Rao, S. Chand, H.C. Upreti, N.K. Singh and T.R. Sharma (2010) Identification of major quantitative trait loci qSBR11-1 for sheath blight resistance in rice. Mol. Breed. 25: 155–166.
- Chen, Y., A.F. Zhang, W.X. Wang, Y. Zhang and T.C. Gao (2012) Baseline sensitivity and efficacy of thifluzamide in Rhizoctonia solani. Ann. Appl. Biol. 161: 247–254.
- Ferriol, M., B. Picó, P. Fernández de Córdova and F. Nuez (2004) Molecular diversity of a germplasm collection of squash (Cucurbita moschata) determined by SRAP and AFLP markers. Crop Sci. 44: 653–664.
- Groth, D.E. and E.M. Nowick (1992) Selection for resistance to rice sheath blight through number of infection cushions and lesion type. Plant Dis. 76: 721–723.
- Han, Y.P., Y.Z. Xing, Z.X. Chen, S.L. Gu, X.B. Pan, X.L. Chen and Q.F. Zhang (2002) Mapping QTLs for horizontal resistance to sheath blight in an elite rice restorer line, Minghui 63. Acta Genet. Sin. 29: 622–626.
- IRRI (2002) Standard Evaluation Systems for Rice (November 2002). International Rice Institute Research, Manila, Philippines, p. 19.
- Jia, Y., F. Correa-Victoria, A. McClung, L. Zhu, G. Liu, Y. Wamishe, J. Xie, M.A. Marchetti, S.R.M. Pinson, J.N. Rutger et al. (2007) Rapid determination of rice cultivar responses to the sheath blight pathogen Rhizoctonia solani using a micro-chamber screening method. Plant Dis. 91: 485–489.
- JPPA (2013) Epidemic and controlling area in 2012. In: Japan Plant Protection Association (eds.) Catalogue of agricultural chemicals, Tokyo, pp. 572–574.
- Koehler-Santos, P., A.L. Dornelles and L.B. Freitas (2003) Characterization of mandarin citrus germplasm from southern Brazil by morphological and molecular analyses. Pesqui. Agropecu. Bras. 38: 797–806.
- Kovach, W.L. (2007) A MultiVariate Statistical Package (MVSP) for Windows, ver. 3.1. Kovach Computing Services, Pentraeth, Wales, UK.
- Li, Z.K., S.R.M. Pinson, J.W. Stansel and W.D. Park (1995a) Identification of quantitative trait loci (QTLs) for heading date and plant height in cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 91: 374–381.
- Li, Z.K., S.R.M. Pinson, M.A. Marchetti, J.W. Stansel and W.D. Park (1995b) Characterization of quantitative trait loci (QTLs) in cultivated rice contributing to field resistance to sheath blight (Rhizoctonia solani). Theor. Appl. Genet. 91: 382–388.
- Liu, G., Y. Jia, F.J. Correa-Victoria, G.A. Prado, K.M. Yeater, A. McClung and J.C. Correll (2009) Mapping quantitative trait loci responsible for resistance to sheath blight in rice. Phytopathology 99: 1078–1084.
- Liu, J. (2003) PowerMarker: New Genetic Data Analysis Software, Version 4.0. http://www.powermarker.net
- Loan, L.C., P.V. Du and Z.K. Li (2004) Molecular dissection of quantitative resistance of sheath blight in rice (Oryza sativa L.). Omonrice 12: 1–12.
- Mantel, N. (1967) The detection of disease clustering and a generalized regression approach. Cancer Res. 27: 209–220.
- Marchetti, M.A. and C.N. Bollich (1991) Quantification of the relationship between SB severity and yield loss in rice. Plant Dis. 75: 773–775.
- Mew, T.W., H. Leung, S. Savary, C.M. Vera-Cruz and J.E. Leach (2004) Looking ahead in rice disease research and management. CRC Crit. Rev. Plant Sci. 23: 103–127.
- Nei, M. and R.K. Chesser (1983) Estimation of fixation indices and gene diversities. Ann. Hum. Genet. 47: 253–259.
- Ou, S.H. (1985) Rice Diseases, 2nd edn. Commonwealth Agricultural Bureaux, Kew, England, pp. 263–272.
- Pan, X.B., J.H. Zou, Z.X. Chen, J.F. Lu, H.X. Yu, H.T. Li, Z.B. Wang, X.Y. Pan, M.C. Rush and L.H. Zhu (1999) Tagging major quantitative trait loci for sheath blight resistance in a rice variety, Jasmine 85. Chin. Sci. Bull. 44: 1783–1789.
- Pinson, S.R.M., F.M. Capdevielle and J.H. Oard (2005) Confirming QTLs and finding additional loci conditioning sheath blight resistance in rice using recombinant inbred lines. Crop Sci. 45: 503–510.
- Prasad, B. and G.C. Eizenga (2008) Rice sheath blight disease resistance identified in Oryza spp. accessions. Plant Dis. 92: 1503–1509.
- Rodrigues, F.Á., F.X.R. Vale, L.E. Datnoff, A.S. Prabhu and G.H. Korndörfer (2003) Effect of rice growth stages and silicon on sheath blight development. Phytopathology 93: 256–261.
- Rohlf, F.J. (2000) NTSYS-pc: Numerical Taxonomy System.Ver. 2.1. Exeter Software, Setauket, NY, USA, pp. 29–34.
- Rohlf, F.J. (2008) NTSYS-pc. Numerical taxonomy and multivariate analysis system, Version 2.2. Exeter Software, Setauker, New York.
- Rush, M.C. and F.N. Lee (1992) Sheath blight. In: Webster, R.K. and P.S. Gunnell (eds.) Compendium of Rice Diseases, The American Phytopathological Society, St. Paul, MN, pp. 22–23.
- Sambrook, J., E.F. Frtsch and T. Maniatis (1989) Molecular Cloning: A Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory Press, NY, USA.
- Sato, H., O. Ideta, I. Ando, Y. Kunihiro, H. Hirabayashi, M. Iwano, A. Miyasaka, H. Nemoto and T. Imbe (2004) Mapping QTLs for sheath blight resistance in the rice line WSS2. Breed. Sci. 54: 265–271.
- Shahjahan, A.K.M., N.R. Sharma, H.U. Ahmad and S.A. Miah (1986) Yield loss in modern rice varieties due to SB in Bangladesh. J. Agril. Res. 11: 82–90.
- Sharma, N.R., P.S. Teng and F.M. Olivares (1990) Comparison of assessment methods for rice sheath blight disease. Philipp. Phytopathol. 26: 20–24.
- Sharma, A., A.M. McClung, S.R.M. Pinson, J.L. Kepiro, A.R. Shank, R.E. Tabien and R. Fjellstrom (2009) Genetic mapping of sheath blight resistance QTLs within 2 tropical japonica rice cultivars. Crop Sci. 49: 256–264.
- Sneath, P.H.A. and R.R. Sokal (1973) Numerical Taxonomy: The Principals and Practice of Numerical Classification, W.H. Freeman and Co., San Francisco, Calif, USA. Teng, P.S., C.Q. Torries, F.L. Nuque, S.B. Calvero (eds.) 1990. Current knowledge on crop losses in tropical rice. In: IRRI Crop loss assessment in rice. International Rice Research Institute, Los Banos, pp. 39–54.
- Srinivasachary, L. Willocquet and S. Savary (2011) Resistance to rice sheath blight (Rhizoctonia solani Kühn) [(teleomorph: Thanatephorus cucumeris (A.B. Frank) Donk.] disease: current status and perspectives. Euphytica. 178: 1–22.
- Taguchi-Shiobara, F., H. Ozaki, H. Sato, H. Maeda, Y. Kojima, T. Ebitani and M. Yano (2013) Mapping and validation of QTLs for rice sheath blight resistance. Breed. Sci. 63: 301–308.
- Tang, J.B., B.T. Ma, L.Z. Wang, P. Li, A.P. Zheng and H. Chen (2002) Biological control of rice sheath blight with Trichoderma. Chinese J. Rice Sci. 16: 63–66.
- Xie, X.W., M.R. Xu, J.P. Zang, Y. Sun, L.H. Zhu, J.L. Xu, Y.L. Zhou and Z.K. Li (2008) Genetic background and environmental effects on QTLs for sheath blight resistance revealed by reciprocal introgression lines in rice. Acta Agron. Sin. 34: 1885–1893.
- Zeng, Y.X., Z.J. Ji, L.Y. Ma, X.M. Li and C.D. Yang (2011) Advances in mapping loci conferring resistance to rice sheath blight and mining Rhizoctonia solani resistant resources. Rice Sci. 18: 56–66.
- Zou, J.H., X.B. Pan, Z.X. Chen, J.Y. Xu, J.F. Lu, W.X. Zhai and L.H. Zhu (2000) Mapping quantitative trait loci controlling sheath blight resistance in two rice cultivars (Oryza sativa L.). Theor. Appl. Genet. 101: 569–573.
- Zuo, S., Y. Zhang, Y. Yin, G. Li, G. Zhang, H. Wang, Z. Chen and X. Pan (2014) Fine-mapping of qSB-9TQ, a gene conferring major quantitative resistance to rice sheath blight. Mol. Breed. 34: 2191–2203.


