Abstract
Low molecular weight glutenin subunits are important components of wheat storage proteins, which play an important role in determining end-use quality of common wheat. A newly established matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) procedure was used to analyze 478 landraces of bread wheat collected from the Yangtze-River region in China. Results indicated that 17 alleles at three loci: Glu-A3, Glu-B3 and Glu-D3 were identified, resulting in 87 different allele combinations. Of the 17 alleles detected at all the Glu-3 loci, five belonged to Glu-A3, seven to Glu-B3 and five to Glu-D3 locus. MALDI-TOF-MS indicated Glu-A3a/c was present in 72.8%, Glu-A3b in 8.4%, Glu-A3d in 8.4%, Glu-A3f in 5.2% and Glu-A3e in 3.6% lines. Seven types of alleles were identified at the Glu-B3 locus: Glu-B3d/i (25.5%), Glu-B3b (21.3%), Glu-B3c (16.9%), Glu-B3h (13.8%), Glu-B3f (8.4%), Glu-B3a (8.2%), and Glu-B3g (5.2%). Five types of Glu-D3 alleles were detected: Glu-D3a (58.4%), Glu-D3c (22.6%), Glu-D3d (15.5%), Glu-D3b (3.3%) and Glu-D3f (0.2%). Four new alleles that showed abnormal MALDI-TOF spectrum patterns were identified at the Glu-A3 and Glu-B3 loci. A detailed study is needed to further characterize these alleles and their potential usage for wheat improvement.
Introduction
Gluten proteins, responsible for the functional characteristics of wheat flour products, are conventionally divided into monomeric gliadins and polymeric glutenins, which determine the rheological characteristics (strength and extensibility) of flour dough (Békés et al. 2001, Butow et al. 2003, Fu and Kovacs 1999, Ma et al. 2005, Maucher et al. 2009). The glutenins are divided into high molecular weight glutenin subunits (HMW-GSs) and low molecular weight glutenin subunits (LMW-GSs) based on their mobility during sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE).
The LMW-GSs, which are further subdivided into B, C and D groups on the basis of their mobility in SDS-PAGE and their isoelectric points (Jackson et al. 1983), ranging in molecular mass from 25 to 43 kDa (Liu et al. 2010). They represent about one-third of the total seed protein, account for approximately 60% of total glutenins (Bietz and Wall 1973), and are essential contributors that determine dough properties in wheat, such as dough extensibility (Cornish et al. 2001) and gluten strength (Gianibelli et al. 2001). Different allelic forms of LMW-GSs play different roles in determining different quality parameters (Appelbee 2007, He et al. 2005, Luo et al. 2001). Therefore, it is essential to identify the allelic composition of LMW-GSs in different wheat varieties in order to efficiently use the allele in wheat breeding.
LMW-GSs exhibit high polymorphic protein complexes encoded by a multigene family (D’Ovidio and Masci 2004). Generally, the genes encoding LMW-GSs are located at the Glu-A3, Glu-B3 and Glu-D3 loci on the short arms of chromosomes 1A, 1B and 1D, respectively (Pogna et al. 1990, Singh and Shepherd 1988). The LMW-GSs are not as well assessed as the HMW-GSs by SDS-PAGE because of their large numbers of subunits and the complexity of the LMW-GS patterns (Huang and Cloutier 2008, Zhang et al. 2011). Multiple analytical procedures have been established to accurately analyze the HMW-GS in bread wheat, including one-dimensional SDS-PAGE (Branlard et al. 2003), 2-DE (An et al. 2005), PCR (Ma et al. 2003) and MALDI-TOF (Liu et al. 2009). Due to the complexity, accurate identification of LMW-GS alleles has been proven to be a difficult task. The SDS-PAGE procedure often results in errors. The 2-DE analytical process can display much more information than SDS-PAGE, but is not generally recommended in breeding programs due to its time consuming and high cost nature. Recently, PCR approach has become a powerful tool for characterizing LMW-GS composition in common wheat (Appelbee et al. 2009, Ram et al. 2011, Wang et al. 2009, 2010, Zhang et al. 2011). However, allele specific PCR markers are only available for a proportion of LMW-GS alleles, which limited its usage in wheat breeding. Recently, MALDI-TOF-MS has been put into use for analyzing LMW-GS alleles (Liu 2008, Wang 2008). It is the most efficient method to characterize wheat gluten proteins and requires only a few minutes per sample to perform the analysis (Liu et al. 2009, Wang 2008, Zheng et al. 2011). The standard allele specific MALDI-TOF spectrum patterns have been established for most known LMW-GS alleles by Wang et al. (2015), which enables fast and reliable identification of LMW-GS alleles.
China is considered as a secondary centre of origin for common wheat (Dong and Zheng 2000) according to its history of wheat production and the extensive genetic variation that is identified in the middle and low branches of the Yellow river (i.e. in Henan and Shandong provinces). There are a mass of wheat landraces that have been accumulated. The Chinese National Germplasm Bank stores more than ten thousand Chinese wheat landrace accessions collected from various wheat production regions. Some novel HMW-GS alleles have been found in the Chinese wheat landraces (Fang et al. 2009, Guo et al. 2010, Liu et al. 2007, Zheng et al. 2011). Unfortunately, little is known about the LMW-GS allelic compositions due to lack of efficient analytical procedures. This has hampered the usage of Chinese landraces LMW-GS variations in wheat breeding. The objective of this study was to detect the LMW-GS allelic compositions of Chinese wheat landraces from the Yangtze-River region.
Materials and Methods
Plant material
Four hundred and seventy-eight accessions of bread wheat (Triticum aestivum L.) landraces were collected from the Yangtze-River region (389 landraces from Hubei province, and 89 landraces from Tibet Autonomous Region) in China, including most accessions collected by Huazhong Agricultural University Wuhan, Hubei province in the past fifty years (Supplemental Table 1).
Protein extraction
Proteins were extracted from whole meal based on the procedure reported by Wang et al. (2015). Whole meal (100 mg) was suspended in 1.0 ml of 50% propanol-1-ol (v/v) for 5 min continuous vortexing, followed by incubation at 65°C for 20 min, vortexing for 5 min, and centrifugation at 10,000 × g for 5 min. This step was repeated three times to remove the majority of the gliadins. The glutenin in the pellet was reduced with 50 mM Tris-HCl buffer containing 50% propanol-1-ol and 1% w/v dithiothreitol, then 1.4% v/v of 4-vinylpyridine was added, and alkylation was carried out overnight at room temperature.
MALDI-TOF-MS protocol and nomenclature
MALDI-TOF-MS was carried out at the State Agriculture Biotechnology Center, Murdoch University, Australia. Acetone (80%) was used to precipitate the glutenin fraction, and the resulting pellets were dissolved in 60 μL acetonitrile/H2O (50:50 v/v) with 0.05% v/v trifluoroacetic acid (60 min at room temperature). Sample preparation was performed on the basis of the dried droplet method (Kussmann et al. 1997), employing sinapinic acid as matrix. The sinapinic acid was dissolved in acetonitrile/H2O (50:50 v/v) with 0.05% v/v trifluoroacetic acid at a concentration of 10 mg/ml.
A sandwich matrix/sample/matrix 1:1:1 (0.7 μL) was placed to a 100-sample MALDI probe tip, and dried at room temperature.
MALDI-TOF-MS was carried out on a Voyager DE-PRO TOF mass spectrometer (Applied Biosystems, Foster City, CA, USA) equipped with nitrogen laser (337 nm) and delayed extraction. Analyses were carried out on a positive linear ion mode with the following parameters: mass range 10000–50000 Da, acceleration voltage 25 kV, and delay time 900 ns. Ten thousands Da was selected as the low mass gate value for analysis to avoid saturation of the detector. The allele specific MALDI-TOF spectrum patterns followed the newly established system (Wang et al. 2015). The LMW-GS nomenclature system of Ikeda et al. (2008) was used.
Result
By following the new method of Wang et al. (2015), the mass spectra of the LMW-GSs displayed distinct, well-separated spectrum peaks for all wheat lines, with the molecular masses ranging from 25,000 to 44,000 Da in the spectra. Representative spectra are shown in Fig. 1 to Fig. 8. The allele combinations and variants at the Glu-3 loci identified by MALDI-TOF-MS are shown in Table 1. All scored alleles matched well with the reported allele specific spectrum peak patterns (Wang et al. 2015). Low level of peak distortion or position shifting in a specific pattern was observed.


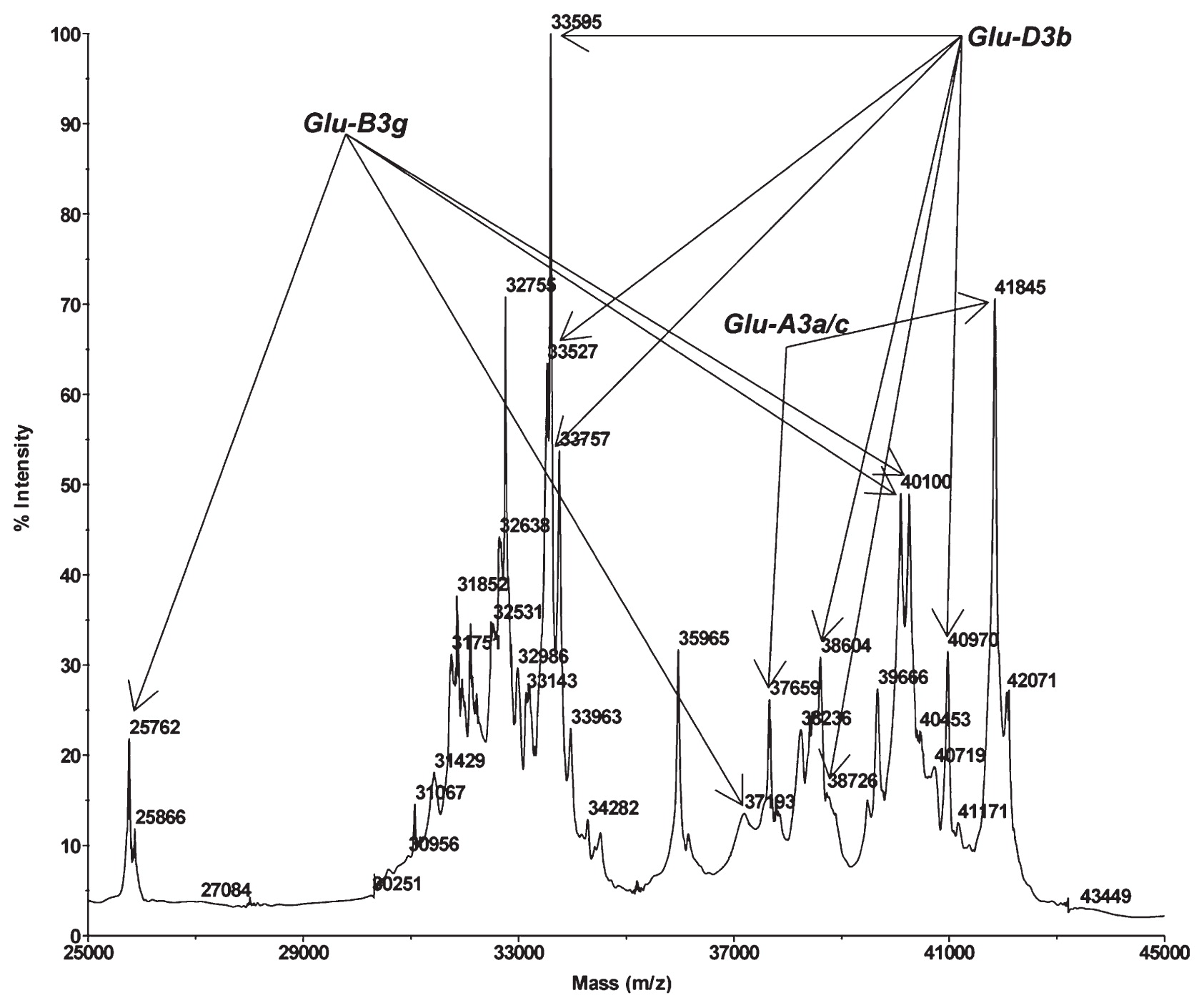





Table 1
Allele combinations and variants at the
Glu-3 loci in Chinese wheat landraces of the Yangtze-River region
|
Glu-A3 |
Glu-B3 |
Glu-D3 |
Varieties |
Frequency (%) |
|
Glu-A3 |
Glu-B3 |
Glu-D3 |
Varieties |
Frequency (%) |
| 1 |
43268Da |
a |
a |
1 |
0.2 |
45 |
b |
c |
d |
6 |
1.3 |
| 2 |
43338Da |
d/i |
a |
1 |
0.2 |
46 |
b |
d/i |
a |
1 |
0.2 |
| 3 |
43354Da |
a |
a |
1 |
0.2 |
47 |
b |
d/i |
c |
1 |
0.2 |
| 4 |
43384Da |
h |
a |
1 |
0.2 |
48 |
b |
d/i |
d |
2 |
0.4 |
| 5 |
43385Da |
h |
d |
1 |
0.2 |
49 |
b |
f |
d |
1 |
0.2 |
| 6 |
43754Da |
b |
b |
1 |
0.2 |
50 |
b |
g |
a |
3 |
0.6 |
| 7 |
43770Da |
a |
a |
1 |
0.2 |
51 |
b |
h |
a |
3 |
0.6 |
| 8 |
43781Da |
b |
a |
1 |
0.2 |
52 |
b |
h |
c |
1 |
0.2 |
| 9 |
a/c |
40563Da |
d |
1 |
0.2 |
53 |
b |
h |
d |
4 |
0.8 |
| 10 |
a/c |
40576Da |
b |
1 |
0.2 |
54 |
d |
b |
a |
6 |
1.3 |
| 11 |
a/c |
40642Da |
a |
1 |
0.2 |
55 |
d |
b |
c |
3 |
0.6 |
| 12 |
a/c |
a |
a |
19 |
4.0 |
56 |
d |
b |
d |
2 |
0.4 |
| 13 |
a/c |
a |
b |
1 |
0.2 |
57 |
d |
c |
a |
3 |
0.6 |
| 14 |
a/c |
a |
c |
4 |
0.8 |
58 |
d |
c |
d |
2 |
0.4 |
| 15 |
a/c |
a |
d |
4 |
0.8 |
59 |
d |
f |
a |
6 |
1.3 |
| 16 |
a/c |
b |
a |
45 |
9.4 |
60 |
d |
f |
c |
2 |
0.4 |
| 17 |
a/c |
b |
b |
1 |
0.2 |
61 |
d |
f |
d |
2 |
0.4 |
| 18 |
a/c |
b |
c |
19 |
4.0 |
62 |
d |
g |
b |
1 |
0.2 |
| 19 |
a/c |
b |
d |
11 |
2.3 |
63 |
d |
g |
c |
3 |
0.6 |
| 20 |
a/c |
c |
a |
30 |
6.3 |
64 |
d |
g |
d |
1 |
0.2 |
| 21 |
a/c |
c |
b |
2 |
0.4 |
65 |
d |
h |
a |
3 |
0.6 |
| 22 |
a/c |
c |
c |
12 |
2.5 |
66 |
d |
h |
b |
2 |
0.4 |
| 23 |
a/c |
c |
d |
10 |
2.1 |
67 |
d |
h |
c |
2 |
0.4 |
| 24 |
a/c |
c |
f |
1 |
0.2 |
68 |
d |
h |
d |
2 |
0.4 |
| 25 |
a/c |
d/i |
a |
66 |
13.8 |
69 |
e |
a |
a |
1 |
0.2 |
| 26 |
a/c |
d/i |
b |
3 |
0.6 |
70 |
e |
a |
c |
1 |
0.2 |
| 27 |
a/c |
d/i |
c |
21 |
4.4 |
71 |
e |
b |
a |
2 |
0.4 |
| 28 |
a/c |
d/i |
d |
10 |
2.1 |
72 |
e |
c |
a |
3 |
0.6 |
| 29 |
a/c |
f |
a |
17 |
3.6 |
73 |
e |
c |
c |
1 |
0.2 |
| 30 |
a/c |
f |
c |
7 |
1.5 |
74 |
e |
d/i |
a |
3 |
0.6 |
| 31 |
a/c |
f |
d |
2 |
0.4 |
75 |
e |
d/i |
c |
1 |
0.2 |
| 32 |
a/c |
g |
a |
6 |
1.3 |
76 |
e |
f |
a |
2 |
0.4 |
| 33 |
a/c |
g |
b |
1 |
0.2 |
77 |
e |
g |
a |
1 |
0.2 |
| 34 |
a/c |
g |
c |
9 |
1.9 |
78 |
e |
h |
a |
2 |
0.4 |
| 35 |
a/c |
h |
a |
30 |
6.3 |
79 |
f |
a |
a |
2 |
0.4 |
| 36 |
a/c |
h |
b |
1 |
0.2 |
80 |
f |
b |
c |
4 |
0.8 |
| 37 |
a/c |
h |
c |
5 |
1.0 |
81 |
f |
b |
d |
3 |
0.6 |
| 38 |
a/c |
h |
d |
8 |
1.7 |
82 |
f |
c |
a |
1 |
0.2 |
| 39 |
b |
a |
a |
3 |
0.6 |
83 |
f |
d/i |
a |
9 |
1.9 |
| 40 |
b |
a |
b |
1 |
0.2 |
84 |
f |
d/i |
c |
2 |
0.4 |
| 41 |
b |
b |
b |
1 |
0.2 |
85 |
f |
d/i |
d |
2 |
0.4 |
| 42 |
b |
b |
c |
3 |
0.6 |
86 |
f |
f |
a |
1 |
0.2 |
| 43 |
b |
c |
a |
4 |
0.8 |
87 |
f |
h |
a |
1 |
0.2 |
| 44 |
b |
c |
c |
6 |
1.3 |
|
|
|
|
|
|
At the Glu-A3 locus, five alleles were identified, with the most frequent one being Glu-A3a/c present in 72.8% of the lines. Glu-A3b (8.4%) and Glu-A3d (8.4%) were less common. The less frequent alleles were the Glu-A3f (5.2%) and Glu-A3e (3.6%). Three novel alleles were detected which were represented by three unrecognized peak patterns. A new subunit with molecular weight of 43,754 Da was found in three lines (66, 778, 793). Line 657 contained a new subunit of 43,268 Da, and three lines (158, 170, 693) had a new Glu-A3 type subunit of 43,350 Da. These new Glu-A3 subunits are located around the Glu-A3d position in the spectra.
Seven alleles were identified at the Glu-B3 locus. Overall, the Glu-B3 locus had four predominant allele compositions including Glu-B3d/i (25.5%), Glu-B3b (21.3%), Glu-B3c (16.9%) and Glu-B3h (13.8%). The rest three alleles, namely Glu-B3f, Glu-B3a and Glu-B3g, existed in 8.4%, 8.2% and 5.2% of the tested landraces, respectively. Only one new LMW-GS was identified at the Glu-B3 locus which was present in three lines (241, 628, 649), with the molecular weight of 40,600 Da.
Glu-D3 allele usually consists of high number of spectrum peaks. However, accurate identification of all known Glu-D3 alleles was achieved. Five alleles were observed at the Glu-D3 locus. The frequencies of Glu-D3a and Glu-D3c were 58.4% and 22.6%, respectively. Glu-D3d was found in 15.5% of the landraces and Glu-D3b was present in 3.3% of the landraces. Glu-D3f was observed only in one landrace (line155).
Discussion
Allele identification of glutenins is important for promoting wheat quality. Many favorable alleles with positive effects on dough characteristics and bread-making quality are encoded by Glu-1 and Glu-3 loci. Accurate identification of these alleles is essential for selecting parents in crossing and accumulating them by pyramidal breeding. Meanwhile, discovery of new allele will further aid the wheat quality improvement efforts. Chinese wheat landraces are well known to harbor novel genes. Characterization of glutenin compositions of Chinese landraces will make it possible to utilize these old wheat lines in modern wheat breeding.
In this study, the allelic compositions of LMW-GSs in 478 wheat landraces collected from the Yangtze-River region of China were identified using the newly established MALDI-TOF procedure (Wang et al. 2015). The landraces were collected from Hubei province (Huazhong1–Huazhong700), and Tibet Autonomous Region (Huazhong740–Huazhong833) (Supplemental Table 1). Concerning the landraces from Hubei province, the most frequent alleles at Glu-3 were Glu-A3a/c (74.6%), Glu-A3b (7.7%), Glu-A3d (7.2%); Glu-B3d/i (30.6%), Glu-B3c (20.3%), Glu-B3b (15.9%), Glu-B3h (13.4%); Glu-D3a (59.4%), Glu-D3c (20.1%), Glu-D3d (17.0%). As for the landraces from Tibet Autonomous Region, the most frequent alleles at Glu-3 were Glu-A3a/c (65.2%), Glu-A3d (13.5%), Glu-A3b (11.2%); Glu-B3b (44.9%), Glu-B3f (19.1%), Glu-B3h (15.7%), Glu-B3g (12.4%); Glu-D3a (53.9%), Glu-D3c (33.7%). Some landraces with the same name were collected from different regions of the Yangtze-River; however, they are phenotypical different in many agronomic traits, and published results identified that they are different landraces (Zheng et al. 2011), but farmers called them the same name.
For the Glu-A3 locus, the most frequent alleles were Glu-A3a and Glu-A3c. Bellil et al. (2010, 2012), Bradová and Štočková (2010), Branlard et al. (2003) and Igrejas et al. (1999) also reported a similar conclusion in that Glu-A3a was the predominant allele in wheat cultivars with the frequency of 49.3% among 69 cultivars grown in France, 60.0% among 40 cultivars of Saharan wheats originating from Algerian oases, 47.0% in a collection of 86 Czech registered winter wheat varieties, 44.5% among a set of 200 hexaploid wheat cultivars grown commonly in France and 57.1% among 63 bread wheats primarily grown in Portugal, respectively. The frequency of Glu-A3c allele among 65 accessions representing a historical trend in the cultivars released or introduced in Iran from the year 1940 to 1990 was 40.3% (Izadi-Darbandi et al. 2010), and a higher frequency (40.8%) was also identified in a diverse set of 103 cultivars of common wheat collected from 12 countries including 21 cultivars from China, 19 from Argentina, 15 from Australia, 14 from France, 10 from Japan, 8 from Mexico, 7 from Canada, 3 from the USA, two from the Netherlands, two from Italy, one from Germany and one from Finland (Liu et al. 2010). Thus, these results indicated that the alleles Glu-A3a and Glu-A3c are worldwide predominance among bread wheat. It is worthy of noting that the Glu-A3d is a desirable allele for gluten quality and pan bread quality (He et al. 2005) and was present in 40 landraces. Moreover, the four newly identified Glu-A3 alleles are all located around the Glu-A3d location in the spectra, suggesting that it is novel glu-A3d sub-alleles. The previously discovered unfavorable Glu-A3e allele that reduced the maximum resistance and extensibility of dough (Appelbee 2007) was found to be the least frequent allele (3.5%) in the studied Chinese wheat landraces, suggesting the high value of Chinese wheat landraces for modern wheat quality improvement.
The number of alleles identified at the Glu-B3 (7 alleles) was the same to that reported by Bradová and Štočková (2010) in the Czech winter wheat cultivars. The predominant alleles in our study were Glu-B3d/i and Glu-B3b. The same result was observed in Saharan bread wheat cultivars and French cultivars for Glu-B3d allele, but Glu-B3b was rare in both collections (Bellil et al. 2010, 2012). On the contrary, Bradová and Štočková (2010), Branlard et al. (2003), and Shan et al. (2007) reported that Glu-B3g was the most frequent allele in their collections.
In China, Glu-D3 alleles were typically classified as Glu-D3a, Glu-D3b, Glu-D3c, Glu-D3d and Glu-D3f (Liu 2008). The frequency of Glu-D3 alleles from our study was primarily agreeable with previous report (Liu 2008). In comparison with Glu-D3 allele frequencies previously found in 233 Chinese bread wheats (Liu 2008), both Glu-D3a and Glu-D3c accounted for a high proportion, while the rare allele Glu-D3f was observed only in six lines in Liu’s study (Liu 2008) and one line in our study. Some research discovered only minor Glu-D3 effects on end-use quality traits (Branlard et al. 2003, Eagles et al. 2002, Gupta et al. 1994, He et al. 2005), while other researchers found more important effects (Appelbee 2007, Dong et al. 2010, Flaete and Uhlen 2003, Jin et al. 2013, Luo et al. 2001, Maucher et al. 2009, Park et al. 2011). For example, Glu-D3h was found to display a significantly positive effect on dough rheological quality. Unfortunately, this allele was not discovered in the Chinese landraces used here.
Based on the bread wheat nomenclature reported by Ikeda et al. (2008) for LMW-GSs, we were able to identify the LMW-GS allele compositions of 467 out of the 478 landraces. The other 11 lines contained four novel alleles with each expressing a subunit with molecular weight of about 40,600 Da, 43,268 Da, 43,385 Da, or 43,754 Da. The four novel subunits may play a positive role in determining the viscoelastic properties of wheat, and meeting new end-product requirement. A more detailed study is required to characterize the four novel alleles. Recently, numerous LMW-GS genes have been cloned and characterized (Jiang et al. 2008, Li et al. 2008, Zhang et al. 2010, Zhao et al. 2006, 2008). Based on the available knowledge, the newly identified Glu-3 alleles should be readily cloned and analyzed.
In conclusion, Seventeen known LMW-GS alleles and four novel alleles were found in a collection of 478 landraces from the Yangtze-River region of China. The information obtained in this study is useful for wheat breeders to make decisions on crossing and selection strategies to improve wheat quality, especially to breed new cultivars to meet specific end-product requirements. The results also add to our understanding of genetic effects of LMW-GSs.
Acknowledgments
This research is financially supported by China National Key Project Grant No. 2016YFD0100102.
Literature Cited
- An, X.L., Q.Y. Li, Y.M. Yan, Y.H. Xiao, S.L.K. Hsam and F.J. Zeller (2005) Genetic diversity of European spelt wheat (Triticum aestivum ssp. spelta L. em. Thell.) revealed by glutenin subunit variations at the Glu-1 and Glu-3 loci. Euphytica 146: 193–201.
- Appelbee, M.J. (2007) Quality potential of gluten proteins in hexaploid wheat and related species. PhD thesis, University of Adelaide.
- Appelbee, M.J., G.T. Mekuria, V. Nagasandra, J.P. Bonneau, H.A. Eagles, R.F. Eastwood and D.E. Mather (2009) Novel allelic variants encoded at the Glu-D3 locus in bread wheat. J. Cereal Sci. 49: 254–261.
- Békés, F., P.W. Gras, R.S. Anderssen and R. Appels (2001) Quality traits of wheat determined by small-scale dough testing methods. Crop Pasture Sci. 52: 1325–1338.
- Bellil, I., A. Bouguennec and D. Khelifi (2010) Diversity of seven glutenin and secalin loci within triticale cultivars grown in France. Not. Bot. Horti. Agrobot. Cluj Napoca 38: 48–55.
- Bellil, I., C.M. Bouziani and D. Khelifi (2012) Genetic diversity of high and low molecular weight glutenin subunits in Saharan bread and durum wheats from Algerian oases. Czech J. Genet. Plant Breed. 48: 23–32.
- Bietz, J.A. and J.S. Wall (1973) Isolation and characterization of gliadin-like subunits from glutelin. Cereal Chem. 50: 537–547.
- Bradová, J. and L. Štočková (2010) Evaluation of winter wheat collection in terms of HMW- and LMW-glutenin subunits. Czech J. Genet. Plant Breed. 46 (Special Issue): 96–99.
- Branlard, G., M. Dardevet, N. Amiour and G. Igrejas (2003) Allelic diversity of HMW and LMW glutenin subunits and omega-gliadins in French bread wheat (Triticum aestivum L.). Genet. Resour. Crop Evol. 50: 669–679.
- Butow, B.J., W. Ma, K.R. Gale, G.B. Cornish, L. Rampling, O. Larroque, M.K. Morell and F. Békés (2003) Molecular discrimination of Bx7 alleles demonstrates that a highly expressed high-molecular-weight glutenin allele has a major impact on wheat flour dough strength. Theor. Appl. Genet. 107: 1524–1532.
- Cornish, G.B., F. Békés, H.M. Allen and D.J. Martin (2001) Flour proteins linked to quality traits in an Australian doubled haploid wheat population. Crop and Pasture Sci. 52: 1339–1348.
- Dong, L.L., X.F. Zhang, D.C. Liu, H.J. Fan, J.Z. Sun, Z.J. Zhang, H.J. Qin, B. Li, S.T. Hao, Z.S. Li et al. (2010) New insights into the organization, recombination, expression and functional mechanism of low molecular weight glutenin subunit genes in bread wheat. PLoS ONE 5: e13548.
- Dong, Y.S. and D.S. Zheng (2000) Wheat genetic resources in China, 1st edn. China Agricultural Press, Beijing, pp. 16–30.
- D’Ovidio, R. and S. Masci (2004) The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci. 39: 321–339.
- Eagles, H.A., G.J. Hollamby, N.N. Gororo and R.F. Eastwood (2002) Estimation and utilisation of glutenin gene effects from the analysis of unbalanced data from wheat breeding programs. Crop Pasture Sci. 53: 367–377.
- Fang, J.Y., Y. Liu, J. Luo, Y.S. Wang, P.R. Shewry and G.Y. He (2009) Allelic variation and genetic diversity of high molecular weight glutenin subunit in Chinese endemic wheats (Triticum aestivum L.). Euphytica 166: 177–182.
- Flaete, N.E.S. and A.K. Uhlen (2003) Association between allelic variation at the combined Gli-1, Glu-3 loci and protein quality in common wheat (Triticum aestivum L.). J. Cereal Sci. 37: 129–137.
- Fu, B.X. and M.I.P. Kovacs (1999) Rapid single-step procedure for isolating total glutenin proteins of wheat flour. J. Cereal Sci. 29: 113–116.
- Gianibelli, M.C., O.R. Larroque, F. MacRitchie and C.W. Wrigley (2001) Biochemical, genetic, and molecular characterization of wheat glutenin and its component subunits. Cereal Chem. 78: 635–646.
- Guo, X.M., J.X. Guo, X.Q. Li, X.M. Yang and L.H. Li (2010) Molecular characterization of two novel Glu-D1-encoded subunits from Chinese wheat (Triticum aestivum L.) landrace and functional properties of flours possessing the two novel subunits. Genet. Resour. Crop Evol. 57: 1217–1225.
- Gupta, R.B., J.G. Paul, G.B. Cornish, G.A. Palmer, F. Bekes and A.J. Rathjen (1994) Allelic variation at glutenin subunit and gliadin loci, Glu-1, Glu-3 and Gli-1, of common wheats. I. Its additive and interaction effects on dough properties. J. Cereal Sci. 19: 9–17.
- He, Z.H., L. Liu, X.C. Xia, J.J. Liu and R.J. Peña (2005) Composition of HMW and LMW glutenin subunits and their effects on dough properties, pan bread, and noodle quality of Chinese bread wheats. Cereal Chem. 82: 345–350.
- Huang, X.Q. and S. Cloutier (2008) Molecular characterization and genomic organization of low molecular weight glutenin subunit genes at the Glu-3 loci in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 116: 953–966.
- Igrejas, G., H. Guedes-Pinto, V. Carnide and G. Branlard (1999) The high and low molecular weight glutenin subunits and ω-gliadin composition of bread and durum wheats commonly grown in Portugal. Plant Breed. 118: 297–302.
- Ikeda, T.M., G. Branlard, R.G. Peña, K. Takata, L. Liu, Z.H. He, S.E. Lerner, M.A. Kolman, H. Yoshida and W.J. Rogers (2008) International collaboration for unifying Glu-3 nomenclature system in common wheats. Proc 11th Int Wheat Genet Symp, Sydney University Press, Sydney, Australia 042.
- Izadi-Darbandi, A., B. Yazdi-Samadi, A. Shanejat-Boushehri and M. Mohammadi (2010) Allelic variations in Glu-1 and Glu-3 loci of historical and modern Iranian bread wheat (Triticum aestivum L.) cultivars. J. Genet. 89: 193–199.
- Jackson, E.A., L.M. Holt and P.I. Payne (1983) Characterisation of high molecular weight gliadin and low-molecular-weight glutenin subunits of wheat endosperm by two-dimensional electrophoresis and the chromosomal localisation of their controlling genes. Theor. Appl. Genet. 66: 29–37.
- Jiang, C.X., Y.H. Pei, Y.Z. Zhang, X.H. Li, D.N. Yao, Y.M. Yan, W.J. Ma, S.L.K. Hsam and F.J. Zeller (2008) Molecular cloning and characterization of four novel LMW glutenin subunit genes from Aegilops longissima, Triticum dicoccoides and T. zhukovskyi. Hereditas 145: 92–98.
- Jin, H., Y. Zhang, G.Y. Li, P.Y. Mu, Z.R. Fan, X.C. Xia and Z.H. He (2013) Effects of allelic variation of HMW-GS and LMW-GS on mixograph properties and Chinese noodle and steamed bread qualities in a set of Aroona near-isogenic wheat lines. J. Cereal Sci. 57: 146–152.
- Kussmann, M., E. Nordhoff, H. Rahbek-Nielsen, S. Haebel, M. Rossel-Larsen, L. Jakobsen, J. Gobom, E. Mirgorodskaya, A. Kroll-Kristensen, L. Palm et al. (1997) Matrix-assisted laser desorption/ionization mass spectrometry sample preparation techniques designed for various peptide and protein analytes. J. Mass Spectrom. 32: 593–601.
- Li, X.H., A.L. Wang, Y.H. Xiao, Y.M. Yan, Z.H. He, R. Appels, W.J. Ma, S.L.K. Hsam and F.J. Zeller (2008) Cloning and characterization of a novel low molecular weight glutenin subunit gene at the Glu-A3 locus from wild emmer wheat (Triticum turgidum L. var. dicoccoides). Euphytica 159: 181–190.
- Liu, L. (2008) Identification of glutenin composition in common wheat by proteomics technology. PhD thesis, Chinese academy of agricultural sciences.
- Liu, L., A.L. Wang, R. Appels, J.H. Ma, X.C. Xia, P. Lan, Z.H. He, F. Bekes, Y. Yan and W. Ma (2009) A MALDI-TOF based analysis of high molecular weight glutenin subunits for wheat breeding. J. Cereal Sci. 50: 295–301.
- Liu, L., T.M. Ikeda, G. Branlard, R.J. Peña, W.J. Rogers, S.E. Lerner, M.A. Kolman, X.C. Xia, L.H. Wang, W.J. Ma et al. (2010) Comparison of low molecular weight glutenin subunits identified by SDS-PAGE, 2-DE, MALDI-TOF-MS and PCR in common wheat. BMC Plant Biol. 10: 124.
- Liu, Y., Z.Y. Xiong, Y.G. He, P.R. Shewry and G.Y. He (2007) Genetic diversity of HMW glutenin subunit in Chinese common wheat (Triticum aestivum L.) landraces from Hubei province. Genet. Resour. Crop Evol. 54: 865–874.
- Luo, C., W.B. Griffin, G. Branlard and D.L. McNeil (2001) Comparison of low- and high molecular-weight wheat glutenin allele effects on flour quality. Theor. Appl. Genet. 102: 1088–1098.
- Ma, W.J., W. Zhang and K.R. Gale (2003) Multiplex-PCR typing of high molecular weight glutenin alleles in wheat. Euphytica 134: 51–60.
- Ma, W.J., R. Appels, F. Bekes, O. Larroque, M.K. Morell and K.R. Gale (2005) Genetic characterisation of dough rheological properties in a wheat doubled haploid population: additive genetic effects and epistatic interactions. Theor. Appl. Genet. 111: 410–422.
- Maucher, T., J.D.C. Figueroa, W. Reule and R.J. Peña (2009) Influence of low molecular weight glutenins on viscoelastic properties of intact wheat kernels and their relation to functional properties of wheat dough. Cereal Chem. 86: 372–375.
- Park, C.S., C.S. Kang, J.U. Jeung and S.H. Woo (2011) Influence of allelic variations in glutenin on the quality of pan bread and white salted noodles made from Korean wheat cultivars. Euphytica 180: 235–250.
- Pogna, N.E., J.C. Autran, F. Mellini, D. Lafiandra and P. Feillet (1990) Chromosome 1B-encoded gliadins and glutenin subunits in durum wheat: genetics and relationship to gluten strength. J. Cereal Sci. 11: 15–34.
- Ram, S., S. Sharma, A. Verma, B.S. Tyagi and R.J. Peña (2011) Comparative analyses of LMW glutenin alleles in bread wheat using allele-specific PCR and SDS-PAGE. J. Cereal Sci. 54: 488–493.
- Shan, X., S.R. Clayshulte, S.D. Haley and P.F. Byrne (2007) Variation for glutenin and waxy alleles in the US hard winter wheat germplasm. J. Cereal Sci. 45: 199–208.
- Singh, N.K. and K.W. Shepherd (1988) Linkage mapping of genes controlling endosperm storage proteins in wheat. 1. Genes on the short arms of group 1 chromosomes. Theor. Appl. Genet. 75: 628–641.
- Wang, A.L. (2008) Development of proteomic approaches for the characterization of wheat major seed proteins and its application in genetics and breeding. PhD thesis, Capital Normal University.
- Wang, A.L., L. Liu, Y.C. Peng, S. Islam, M. Applebee, R. Appels, Y.M. Yan and W.J. Ma (2015) Identification of low molecular weight glutenin alleles by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) in common wheat (Triticum aestivum L.). PLoS ONE 10: e0138981.
- Wang, L.H., X.L. Zhao, Z.H. He, W.J. Ma, R. Appels, R.J. Peña and X.C. Xia (2009) Characterization of low-molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (Triticum aestivum L.). Theor. Appl. Genet. 118: 525–539.
- Wang, L.H., G.Y. Li, R.J. Peña, X.C. Xia and Z.H. He (2010) Development of STS markers and establishment of multiplex PCR for Glu-A3 alleles in common wheat (Triticum aestivum L.). J. Cereal Sci. 51: 305–312.
- Zhang, M.Y., K. Wang, S.L. Wang, X.H. Li, F.J. Zeller, S.L.K. Hsam and Y.M. Yan (2010) Molecular cloning, function prediction and phylogenetic analysis of LMW glutenin subunit genes in Triticum timopheevii (Zhuk.). Plant Breed. 129: 622–629.
- Zhang, X.F., D.C. Liu, W. Jiang, X.L. Guo, W.L. Yang, J.Z. Sun, H.Q. Ling and A.M. Zhang (2011) PCR-based isolation and identification of full-length low-molecular-weight glutenin subunit genes in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 123: 1293–1305.
- Zhao, X.L., X.C. Xia, Z.H. He, K.R. Gale, Z.S. Lei, R. Appels and W.J. Ma (2006) Characterization of three low-molecular-weight Glu-D3 subunit genes in common wheat. Theor. Appl. Genet. 113: 1247–1259.
- Zhao, X.L., Y. Yang, Z.H. He, Z.S. Lei, W.J. Ma, Q.X. Sun and X.C. Xia (2008) Characterization of novel LMW-GS genes at Glu-D3 locus on chromosome 1D in Aegilops tauschii. Hereditas 145: 238–250.
- Zheng, W., Y.C. Peng, J.H. Ma, R. Appels, D.F. Sun and W.J. Ma (2011) High frequency of abnormal high molecular weight glutenin alleles in Chinese wheat landraces of the Yangtze-River region. J. Cereal Sci. 54: 401–408.
