Strictly aerobic acetic acid bacteria (AAB) are classified as a sub-group of the Acetobacteraceae family in the class Alphaproteobacteria. AAB strongly oxidize various sugars, alcohols, and sugar alcohols. Vinegar is industrially produced by oxidative fermentation using AAB, particularly those in Acetobacter and Komagataeibacter (37). In ethanol-containing medium, the bacterial strains of these two genera exhibit three growth phases; an ethanol oxidation phase, acetic acid resistance phase, and acetate over oxidation phase (14, 19). In the ethanol oxidation phase, cells oxidize ethanol to acetic acid via acetoaldehyde with membrane-bound alcohol and aldehyde dehydrogenases linked to the respiratory chain (20). In the acetic acid resistance phase, cells resist autoproduced acetic acid using several mechanisms without assimilating it. In the overoxidation phase, cells assimilate acetate by oxidizing it to CO2 via the TCA cycle (14, 19).
Materials and Methods
Genomic DNA and library preparation
Genomic DNA from SKU1108 for genome sequencing was prepared as previously reported (23). Genomic DNA (~20 μg) was purified with the AMPure Xp Kit (Beckman Coulter, Beverly, MA, USA). DNA was sheared using a Covaris g-TUBE (Covaris, Woburn, MA, USA) following the manufacturer’s recommendations for 20-kb fragments. A sequencing library was prepared using the SMRTbell Template Prep Kit 1.0 (Pacific Biosciences of California, Menlo Park, CA, USA). Libraries for 2 SMRT cells (~10 μg) were constructed using two different methods. Small library fragments were removed using BluePippin from one SMRT cell (Sage Science, Beverly, MA, USA) with a 10-kb cut-off. In the other cell, small library fragments were not removed.
DNA sequencing and assembly
Genome sequencing was performed by Takara Bio using PacBio RSII single-molecule real-time (SMRT) sequencing technology (Pacific Biosciences of California, Menlo Park, CA, USA). The sequence data obtained from 2 SMRT cells were used for the subsequent sequence assembly. Sequencing reads were assembled using Hierarchical Genome Assembly Process 3 (HGAP3) in PacBio SMRT portal version 2.3.0. Three large contigs were assembled with a mean coverage of 387-fold. The assembly was corrected with the Quiver consensus algorithm to obtain a high-accuracy genome assembly (7). The overlap sequences of each contig end were manually edited. Previously reported Illumina sequence reads of the SKU1108 strain were mapped onto these three contigs using Bowtie 2 (18, 22). Unmapped reads were collected and a de novo assembly was performed using SPAdes 3.0.0 (3). The resulting assembly revealed two additional small plasmids: plasmids 3 and 4.
Genome annotation
The gene detection and genome annotation of the chromosome and four plasmid sequences were performed using the auto-annotation package Prokka (29). Protein-coding sequences (CDS) of the complete genome sequence were predicted using Prodigal 2.62 (11, 12). ARAGORN 1.26 and Barrnap 0.4 were used to predict tRNA and rRNA regions, respectively (16). The functional assignments of the predicted CDSs were based on a BLASTP homology search against the previously reported Acetobacter pasteurianus genome and the NCBI non-redundant (NR) database (1). The positions of the start codon were manually checked and edited using the commercial genome sequence editing software, in silico Molecular Cloning v5.3.75 (In Silico Biology, Kanagawa, Japan). All signal peptide genes encoded by the SKU1108 genome were predicted by SignalP 4.1 (26). Clustered-regularly interspaced short palindromic repeats (CRISPRs) in the genome sequence were revealed by MinCED 0.16 (4).
Whole genome alignment and genome map
The genomic sequences of A. pasteurianus 386B (HF677570–HF677572) and A. pasteurianus IFO 3283-01 (=NBRC 3283) (AP011121–AP011127) were downloaded from the NCBI FTP website at ftp.ncbi.nlm.nih.gov. The chromosome sequences of the two A. pasteurianus strains, 386B and NBRC 3283, were independently aligned against that of the strain SKU1108 using NUCmer (15). Unaligned regions were manually assigned as specific regions 1 to 10. The genes located at these specific regions were identified and listed. We generated graphic illustrations of genome alignments using CGView (33). Graphical alignments were generated using the progressiveMauve program with default parameters (8).
Identification of genes associated with thermotolerance
The previously reported mutated sites in the genomes of the TI and TH-3 strains associated with thermotolerance were re-confirmed using a previously reported method (22). The 24 thermotolerant genes conserved in A. tropicalis NBRC 101654 (=SKU1100) and their homologous sequences were also searched for in the three A. pasteurianus complete genomes by BLASTP with an E-value cut-off of 10−10 and sequence overlap (query and subject) ≥70% (1, 30).
Phylogenetic tree construction
A BLASTP search against all proteins encoded by 13 Acetobacter and 1 Gluconacetobacter diazotrophicus Pal 5 genomes was performed using the amino acid sequences of AarC and AarC1 in A. pasteurianus SKU1108 as a query. The resulting hits were aligned using MUSCLE v.3.8.31 at the amino acid sequence level (9, 10). Poorly aligned regions were removed using GBLOCKS version 0.91b (6, 34). A phylogenetic tree was constructed using the PROTGAMMAWAG model in RAxML 8.0.14 with 1,000 bootstrap replicates and visualized with the MEGA 6.1 package (31, 32, 35).
Sequence data deposition
The A. pasteurianus SKU1108 (=NBRC 101655) genome sequence was deposited in DDBJ/EMBL/GenBank under the accession numbers AP014881 to AP014885. The versions described here are the first versions. The BioProject ID is PRJDA65545.
Results and Discussion
General genome features
The genomic DNA of A. pasteurianus SKU1108 consisted of a 2,902,389-bp circular chromosome and four plasmids: plasmid 1 (187,193 bp), plasmid 2 (6,331 bp), plasmid 3 (2,799 bp), and plasmid 4 (2,278 bp), with a GC content of 52.75% (Table 1). In total, 2,662 and 214 CDSs were identified in the chromosome and plasmids, respectively. Putative functions were assigned to 2,002 genes. Signal sequences were searched against all CDSs, and detected in 252 cases (data not shown). In total, 56 tRNAs, 1 transfer-messenger RNA (tmRNA), and 5 sets of ribosomal RNA operons (rrn) were predicted in the chromosome sequence (Table 1).
Table 1
Summary of the complete genome sequence of
Acetobacter pasteurianus SKU1108
| Genome information |
Nucleotide sequence length (bp) |
GC content (%) |
Number of ORFs |
Number of tRNAs |
Number of tmRNAs |
Number of rRNAs |
GenBank accession number |
| chromosome |
2,902,389 |
52.79 |
2,662 |
56 |
1 |
15 |
AP014881 |
| plasmid 1 |
187,193 |
52.02 |
201 |
— |
— |
— |
AP014882 |
| plasmid 2 |
6,331 |
52.88 |
8 |
— |
— |
— |
AP014883 |
| plasmid 3 |
2,799 |
58.20 |
3 |
— |
— |
— |
AP014884 |
| plasmid 4 |
2,278 |
55.71 |
2 |
— |
— |
— |
AP014885 |
| Total |
3,135,082 |
52.75 |
2,876 |
56 |
1 |
15 (5 sets of the rRNA operon) |
— |
The genomes of A. pasteurianus are known to contain numerous genes encoding membrane-bound oxidoreductases. In the SKU1108 genome, we revealed genes that code for the membrane-bound PQQ-dependent alcohol dehydrogenase (ADH), adhAB operon (locus_tag: APT_00084–APT_00083) and its subunit III, adhS (APT_00654), and other PQQ enzymes, such as the membrane-bound glucose dehydrogenase (APT_00581), PQQ-dependent dehydrogenase 1 (APT_02219), and PQQ-dependent dehydrogenase 4 (APT_01465). In addition, genes encoding two membrane-bound aldehyde dehydrogenase operons (APT_00973–APT_00975 and APT_00426–APT_00428) were identified. These genes were also conserved in complete genomes of the strains 386B and NBRC 3283. The respiratory chains of Acetobacter species are known to play crucial roles in energy metabolism (24, 27). Therefore, the gene repertoires of respiratory chains were investigated. The cytochrome ba3 ubiquinol oxidase cyaBACD operon (APT_00087–APT_00090), cytochrome bd ubiquinol oxidase cydBACD operon (APT_01543–APT_01546), cyanide-insensitive ubiquinol oxidase (CIO) cioBA operon (APT_02213–APT_02214), type I NADH-quinone oxidoreductase, nuoA-nuoN operon (APT_01737–APT_01725) and nuoM (APT_00690), two type II NADH dehydrogenase ndh (APT_00547 and APT_02111), heme A synthase ctaA (APT_00075), heme O synthase ctaB (APT_01774), and cytochrome c oxidase subunit ctaD (APT_01775) were conserved in the complete genomes of the strains SKU1108, 386B, and NBRC 3283. In contrast, the cytochrome b subunit petB (APT_01924) of the ubiquinol-cytochrome c reductase (bc1 complex) operon (APT_01924-APT_01926) was disrupted with a stop codon insertion around the start codon site in SKU1108, suggesting that this bc1 complex is not functional in SKU1108. However, a paralog sequence of this gene was also found in the SKU1108 genome (APT_00754). Therefore, a combination of these remaining subunits may construct an active bc1 complex.
Whole genome level comparison in complete genome sequences of three closely related strains
Illeghems and co-workers elucidated the complete genome sequence of the thermotolerant A. pasteurianus 386B (13). They isolated this strain from cocoa bean heap fermentation with heating up to approximately 42–43°C (5). They also showed that the strain 386B rapidly produced acetic acid at a high concentration (17). We previously reported that SKU1108 is more thermotolerant than NBRC 3283 and performs acetic acid fermentation even at 38.5°C (22). Therefore, SKU1108 and 386B were defined as thermotolerant acetic acid fermentation strains. On the other hand, NBRC 3283 was defined as a mesophilic strain, which has the ability to perform fermentation at temperatures up to 37°C, but not at 38.5°C. Thus, in order to elucidate the mechanism underlying thermotolerance, a comparative genome analysis was performed using the complete genome sequences of one mesophilic and two thermotolerant strains. Whole genome level synteny was compared in these three complete genomes using Mauve aligner (Fig. S1). A genome synteny analysis revealed that the synteny of these three genomes was highly conserved.
In order to define the uniquely conserved genomic regions in the SKU1108 genome sequence, three complete genome sequences were mapped and aligned against the SKU1108 chromosome sequence using NUCmer, which is a part of the MUMmer genome alignment package. A circular map of the SKU1108 chromosome sequence with alignments is shown in Fig. 1. Ten strain-specific regions were identified (Table S1). Genes located in these regions are also shown in Table S2. Of these, regions 1, 4, and 6 were only conserved between the thermotolerant strains SKU1108 and 386B. Therefore, genes from these regions may be responsible for the thermotolerance phenotype of these strains and may be classified as thermotolerance-conferring conserved regions. Of the ten specific regions, regions 1, 2, 3, and 9 encode prophage-related proteins such as phage integrase and phage terminase. Therefore, these regions may have been acquired by prophage-insertion.
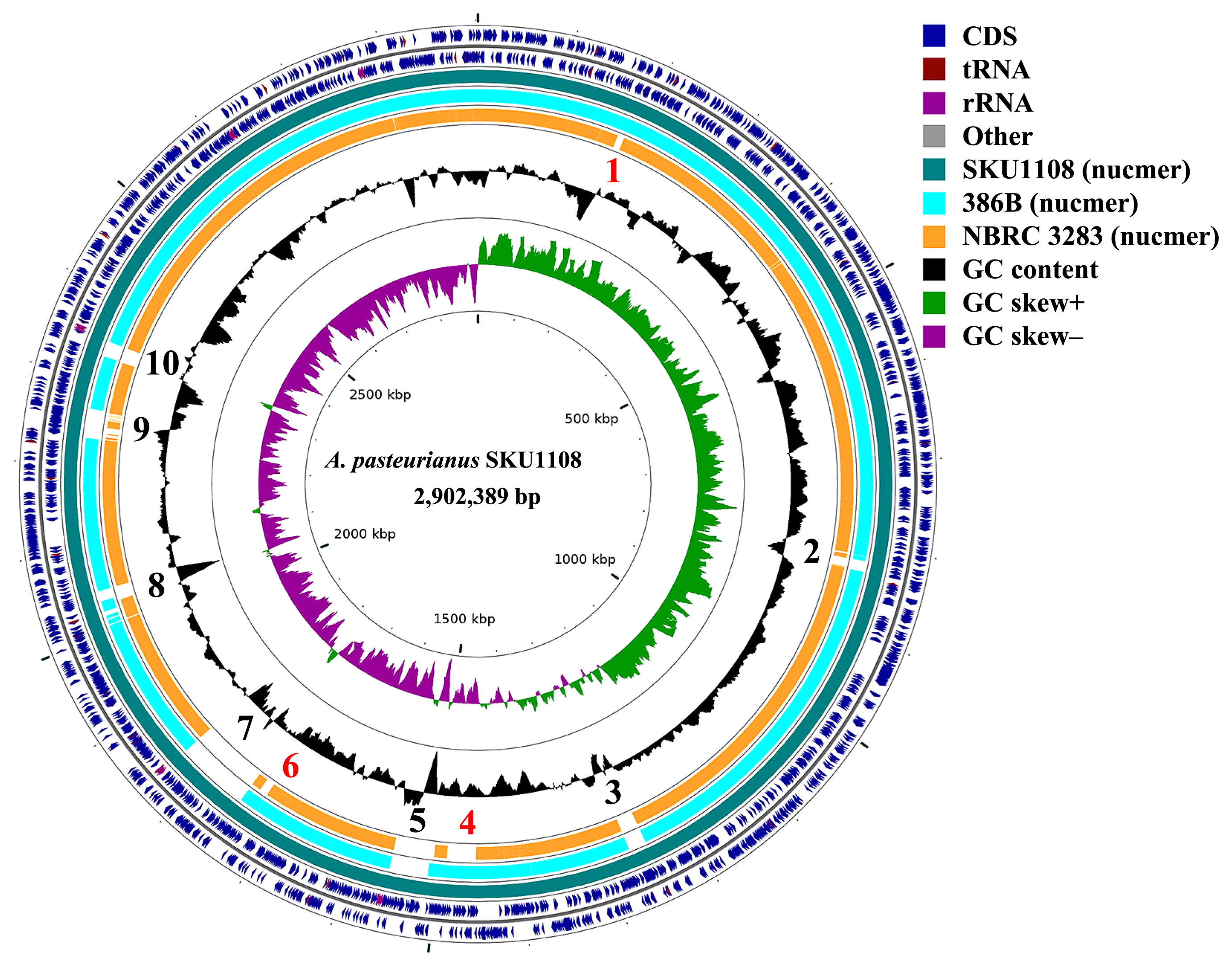
We also compared the repertoire of plasmid sequences among the three studied strains. SKU1108 had one large 187-kb plasmid 1 and three small plasmids (Table 1). On the other hand, 386B and NBRC 3283 had one (194 kb) and two (191 and 182 kb) large plasmids, respectively (2, 13). In order to clarify differences between the large plasmids, we aligned their sequences against the SKU1108 plasmid 1 sequence using NUCmer. Large plasmid 1 of SKU1108 and that of 386B, Apa386Bp1 (HF677571) were highly conserved (Fig. S2). Whole plasmid sequence comparisons also revealed that the synteny of these two plasmids was highly conserved (data not shown). In contrast, the two large plasmids of the strain NBRC 3283 were distinct from that of SKU1108 and not conserved. The results of the whole genome-based phylogenetic analysis suggested that SKU1108 and 386B are more closely related to each other than to NBRC 3283 (Fig. S3). Therefore, since the large plasmids were conserved in the two thermotolerant strains, they may have existed in the ancestor strain, i.e. before the divergence of SKU1108 and 386B into two separate thermotolerant strains.
Genes associated with thermotolerance
In a previous study, we performed the thermal adaptation of SKU1108 by its repeated cultivation under acetic acid fermentation conditions. Consequently, two individually adapted strains, TI and TH-3, were obtained, and their mutational sites were identified using the draft SKU1108 genome assembly as a reference (22). In order to identify additional mutational sites, the previously reported Illumina sequence reads of two adapted strains were mapped onto the complete genomic DNA sequence of SKU1108 that also included the DNAs of four plasmids. No additional mutational sites were detected, indicating that the mapping analysis against the draft genome assembly constructed from Illumina reads was sufficient for a mutation site analysis of the adapted strains. All the mutational sites were detected on the chromosome sequence. Therefore, we have now changed the locus tag number based on the complete genome sequence (Table S3).
In order to identify the genes whose expression is required for growth at higher temperatures, we previously mutated the A. tropicalis SKU1100, a thermotolerant AAB that grows even at 42°C, by transposon mutagenesis, thereby inducing random insertions in the Tn10 transposon using a conjugation method (30). We revealed that 24 “thermotolerant” genes (see Table 2 or Table S4) were associated with growth at higher temperatures. Therefore, in the present study, we clarified whether these 24 genes and their corresponding nucleotide and amino acid mutations were present in complete genomes. A phylogenetic analysis performed for each gene demonstrated that the thermotolerant genes from SKU1100 and three complete A. pasteurianus genomes were clustered in the same clade (data not shown). Therefore, together with the result that all thermotolerant genes were encoded in the conserved regions of all three complete genomes, we concluded that all known thermotolerant genes are conserved in the three genomes, even in the mesophilic strain NBRC 3283 (Table S4). The number of nucleotide/amino acid mutations in the 24 thermotolerant genes and additional 6 genes composed of operons with four genes (DNA methyltransferase [APT_00916], MinC [APT_00954], RpoE [APT_01242], and XdhA [APT_01267]) of the thermotolerant genes, rhnB (APT_00195), minE (APT_00952), minD (APT_00953), rpoE (APT_01243), xdhC (APT_01265), and xdhB (APT_01266) (30 genes in total) are shown in Table 2. The nucleotide sequences of the nine genes were identical in the three strains. There was some variation in nucleotide mutations in 21 other genes, 16 of which mainly accumulated nucleotide mutations in NBRC 3283 (Table 2 and Fig. S3). In contrast, the gene encoding asparagine synthetase (APT_01828) accumulated nucleotide mutations in SKU1108 only, and the gene for DNA methyltransferase in 386B only. In the remaining three genes, flavodoxin/nitric oxide synthase (APT_01927), amiA (APT_02041), and cysG (APT_02252), nucleotide mutations accumulated in all three strains. Non-synonymous mutations in these thermotolerant genes were also investigated and amino acid mutations were detected in 15 (Table 2). Of these, seven genes had amino acid mutations (10 mutations in total) that conferred substitutions by dissimilar amino acids. Amino acid mutations with dissimilar residues only occurred in NBRC 3283, suggesting that they are responsible for the less thermotolerant phenotype of NBRC 3283. Their mutation sites were APT_00246 (A115D), APT_00603 (S95R), APT_01265 (G106E), APT_01266 (T394R and E413G), APT_01267 (R161C, C162R, and T220I), APT_01828 (R471S), and APT_02252 (E185A). In particular, genes encoding the xdhABC operon (APT_01265–APT_01267) accumulated 6 amino acid mutations. Of these, the T220 residue (T206 in Rhodobacter capsulatus) of xdhA (APT_01267), shown to be in the active site in the crystal structure of xdhAB from R. capsulatus, is particularly important because it mediates hydrogen bonding with the co-factor FAD (36). Since mutated T220I cannot form hydrogen bonds with FAD, this mutation may affect the xanthine dehydrogenase activity of NBRC 3283.
Table 2
Number of nucleotide and amino acid mutations of 24 thermotolerant genes among three analyzed strains
| Gene |
Product |
Locus_tag |
Nucleotide mutation |
Amino acid mutation |
|
|
|
|
|
|
|
|
|
| SKU1108 |
SKU1108 |
SKU1108 |
NBRC 3283 |
386B |
SKU1108 |
NBRC 3283 |
386B |
SKU1108 |
| — |
hypothetical protein |
APT_00053 |
0 |
0 |
0 |
0 |
0 |
0 |
| hpnB |
glycosyl transferase |
APT_00180 |
0 |
0 |
0 |
0 |
0 |
0 |
| poxA |
lysyl-tRNA synthetase |
APT_00246 |
13 |
0 |
0 |
2 (1) |
0 |
0 |
| lepA |
GTP-binding protein LepA |
APT_00584 |
9 |
0 |
0 |
1 (0) |
0 |
0 |
| — |
hypothetical protein |
APT_00590 |
4 |
0 |
0 |
0 |
0 |
0 |
| mmpA |
putative metalloprotease MmpA |
APT_00603 |
8 |
0 |
0 |
1 (1) |
0 |
0 |
| hsp20 |
heat shock protein Hsp20 |
APT_00623 |
0 |
0 |
0 |
0 |
0 |
0 |
| — |
hypothetical protein |
APT_00856 |
5 |
0 |
0 |
0 |
0 |
0 |
| glnE |
glutamate-ammonia-ligase adenylyltransferase |
APT_00857 |
0 |
0 |
0 |
0 |
0 |
0 |
| rnhB |
ribonuclease HII |
APT_00915 |
0 |
0 |
0 |
0 |
0 |
0 |
| yhdJ |
DNA methyltransferase |
APT_00916 |
0 |
1 |
0 |
0 |
0 |
0 |
| minE |
cell division topological specificity factor MinE |
APT_00952 |
3 |
0 |
0 |
2 (0) |
0 |
0 |
| minD |
cell division inhibitor MinD |
APT_00953 |
9 |
0 |
0 |
0 |
0 |
0 |
| minC |
cell division inhibitor MinC |
APT_00954 |
4 |
0 |
0 |
1 (0) |
0 |
0 |
| — |
hypothetical protein |
APT_00961 |
16 |
0 |
0 |
0 |
0 |
0 |
| nhaK2 |
Na+/H+ antiporter |
APT_00984 |
69 |
0 |
0 |
1 (0) |
0 |
0 |
| smc |
chromosome segregation protein SMC |
APT_01022 |
44 |
0 |
0 |
9 (0) |
0 |
0 |
| ttg2C |
toluene ABC transporter periplasmic protein |
APT_01083 |
0 |
0 |
0 |
0 |
0 |
0 |
| serA |
d-3-phosphoglycerate dehydrogenase |
APT_01136 |
6 |
0 |
0 |
0 |
0 |
0 |
| rpoE |
RNA polymerase sigma-E factor (sigma-24) protein 2 |
APT_01242 |
0 |
0 |
0 |
0 |
0 |
0 |
| rpoE |
RNA polymerase sigma-E factor (sigma-24) protein 1 |
APT_01243 |
0 |
0 |
0 |
0 |
0 |
0 |
| xdhC |
xanthine dehydrogenase accessory protein XdhC |
APT_01265 |
7 |
1 |
0 |
1 (1) |
1 (0) |
0 |
| xdhB |
xanthine dehydrogenase molybdopterin binding subunit XdhB |
APT_01266 |
48 |
0 |
0 |
9 (2) |
0 |
0 |
| xdhA |
xanthine dehydrogenase XdhA |
APT_01267 |
28 |
0 |
0 |
11 (3) |
0 |
0 |
| — |
asparagine synthetase |
APT_01828 |
1 |
1 |
30 |
1 (1) |
1 (0) |
5 (0) |
| — |
flavodoxin/nitric oxide synthase |
APT_01927 |
5 |
5 |
4 |
2 (0) |
1 (0) |
1 (0) |
| amiA |
N-acetylmuramoyl-l-alanine amidase |
APT_02041 |
11 |
8 |
9 |
2 (0) |
0 |
1 (0) |
| cysG |
siroheme synthase |
APT_02252 |
16 |
5 |
8 |
3 (1) |
0 |
2 (0) |
| — |
hypothetical protein |
APT_02411 |
0 |
0 |
0 |
0 |
0 |
0 |
| htrA |
endopeptidase DegP/Do |
APT_02556 |
21 |
0 |
0 |
3 (0) |
0 |
0 |
Numbers in parentheses indicate residues substituted with dissimilar amino acids. An additional six genes: APT_00915, APT_00952, APT_00953, APT_01243, APT_01265, and APT_01266, which are clustered with 24 thermotolerant genes, are also included in the mutational analysis.
In order to identify other thermotolerant genes only conserved in the two thermotolerant strains, we performed a BLASTP search against all proteins encoded in the three complete genomes using 24 genes of SKU1108 as the query. BLASTP hits (Table 3) showed that four thermotolerant genes, lepA, ttg2C, xdhA, and degP, had paralogs in the chromosome. However, a paralog of xdhA (APT_01390) encoding xanthine dehydrogenase XdhA (APT_01267) was only conserved in the two thermotolerant strains. This is a part of the xdhAB operon (APT_01390–APT_01389) that is encoded in specific region 4 found in the genomes of both thermotolerant strains (Fig. 1 and Table S2). Xanthine dehydrogenase is known to regulate heat shock and stress responses (30). This xdhAB paralog may partly contribute to the thermotolerance of SKU1108 and 386B. Thus, amino acid mutations accumulated in the thermotolerant genes and their paralogs encoded in the conserved thermotolerance-conferring region may be responsible for the thermotolerance of the two thermotolerant strains. As described above, two thermotolerant strains have the same large plasmid 1 (Fig. S2). Since these plasmids are only conserved in two thermotolerant strains, we speculated that the genes responsible for thermotolerance are encoded in these two large plasmids. Therefore, we also searched the paralog sequences encoded in these plasmid sequences by a BLASTP search. In addition, a BLASTP search was performed against all other plasmid sequences of the three strains. However, we were unable to identify any paralog sequences corresponding to the 24 thermotolerant genes in these plasmids.
Table 3
List of paralogous sequences of 24 thermotolerant genes conserved in three
A. pasteurianus strains
Gene
SKU1108 |
Blast query |
Blast hits |
Gene (hits) |
Product (hits) |
E-value |
Identity (%) |
| lepA |
APT_00584 |
APT_00570 |
typA |
GTP-binding protein TypA |
5.00E-32 |
26 |
| APA01_06200 |
bipA |
GTP-binding protein TypA |
5.00E-32 |
26 |
| APA386B_2113 |
typA |
GTP-binding protein TypA/BipA |
5.00E-32 |
26 |
|
| ttg2C |
APT_01083 |
APA386B_1720 |
ttg2C |
toluene transporter substrate-binding periplasmic protein Ttg2C |
4.00E-14 |
35 |
| APT_00211 |
ttg2C |
toluene ABC transporter periplasmic protein |
4.00E-14 |
35 |
| APA01_02300 |
ttg2C |
toluene ABC transporter periplasmic protein |
4.00E-14 |
35 |
|
| xdhA |
APT_01267 |
APA386B_251 |
xdhA |
xanthine dehydrogenase small subunit |
4.00E-53 |
32 |
| APT_01390 |
xdhA |
xanthine dehydrogenase XdhA |
4.00E-53 |
32 |
|
| htrA |
APT_02556 |
APA386B_1296 |
degP |
endopeptidase DegP/Do |
1.00E-94 |
42 |
| APT_02481 |
degP |
endopeptidase DegP/Do |
1.00E-94 |
42 |
| APA01_24860 |
degP |
endopeptidase DegP/Do |
2.00E-94 |
42 |
Incidentally, related to the acetic acid resistance or acetic acid assimilation ability of Acetobacter species, we found an aarC paralog (aarC1) sequence in strain-specific region 7. Acetobacter species have a specialized citric acid cycle in which the aarC gene encoding acetate CoA-transferase plays a crucial role by regulating acetate assimilation (25). A phylogenetic analysis of the AarC protein, conserved in published Acetobacter genome sequences, produced the phylogenetic tree that was divided into two large clades: genes belonging to the aarC clade are conserved in all Acetobacter species, whereas those of the aarC1 clade are only conserved in genomes of the three A. pasteurianus strains studied including SKU1108 (Fig. S4). Thus, it will be interesting to clarify the role of the aarC1 gene in acetate assimilation.
