Abstract
Nitrogen fixing symbiosis between rhizobia and legumes contributes significant amounts of N to agricultural and natural environments. In natural soils, rhizobia compete with indigenous bacterial communities to colonize legume roots, which leads to symbiotic interactions. However, limited information is currently available on the effects of the rhizobial symbiont on the resident microbial community in the legume rhizosphere, rhizoplane, and endosphere, which is partly due to the presence of native nodulating rhizobial strains. In the present study, we used a symbiotic system comprised of Paraburkholderia phymatum and Mimosa pudica to examine the interaction of an inoculant strain with indigenous soil bacteria. The effects of a symbiont inoculation on the native bacterial community was investigated using high throughput sequencing and an analysis of 16S rRNA gene amplicons. The results obtained revealed that the inoculation induced significant alterations in the microbial community present in the rhizoplane+endosphere of the roots, with 13 different taxa showing significant changes in abundance. No significant changes were observed in the rhizospheric soil. The relative abundance of P. phymatum significantly increased in the rhizoplane+endosphere of the root, but significant decreased in the rhizospheric soil. While the rhizosphere, rhizoplane, and root endosphere contained a wide diversity of bacteria, the nodules were predominantly colonized by P. phymatum. A network analysis revealed that the operational taxonomic units of Streptomyces and Phycicoccus were positively associated with P. phymatum as potential keystone taxa. Collectively, these results suggest that the success of an inoculated symbiont depends on its ability to colonize the roots in the face of competition by other soil bacteria. A more detailed understanding of the mechanisms by which an inoculated strain colonizes its plant host is crucial for realizing the full potential of microbial inoculants in sustainable agriculture.
Nitrogen-fixing symbiosis between legumes and rhizobia is a major contributor to the global nitrogen cycle (Sprent and James, 2007). We herein use the word rhizobia to collectively refer to all bacteria forming nitrogen-fixing nodules on legumes, regardless of taxonomy. In agricultural systems, these symbiotic relationships have been estimated to contribute more than 80% of fixed nitrogen (O’Hara, 1998). Rhizobial inoculants are extensively utilized to enhance the growth of crop legumes (Deaker et al., 2004; Narożna et al., 2015; Parnell et al., 2016). However, in many cases, these inoculants fail to increase crop productivity due to competition by indigenous rhizobia that are more adapted to the local soil environment (Triplett and Sadowsky, 1992). Rhizobial interactions with host legumes under axenic laboratory conditions have been characterized in detail (Oldroyd et al., 2011; Oldroyd, 2013; Venkateshwaran et al., 2013) and typically start with the colonization of and attachment to plant roots (and sometimes stems) (Poole et al., 2018; Wheatley and Poole, 2018). However, limited information is currently available on the genetic and physiologic traits that are important for inoculated rhizobia that need to survive and compete with other soil microbes to reach and colonize legume roots (Poole et al., 2018). Moreover, soil edaphic factors, the number of indigenous rhizobia, biotic-biotic interactions, and climate all play critical roles in the success of added legume inoculants (Triplett and Sadowsky, 1992). A more detailed understanding of this process is crucial for realizing the full potential of nitrogen-fixing and plant growth-promoting microbial inoculants in commercial agriculture.
The soil immediately in contact with plant roots (the rhizosphere) is rich in microbial numbers and diversity, in large part due to the secretion of substantial amounts of carbon, nitrogenous compounds, and other nutrients into the rhizosphere (Walker et al., 2003; Jones et al., 2009). The majority of the thus far characterized legume-nodulating rhizobia belonging to Alphaproteobacteria are metabolically diverse and function well as soil saprophytes in the absence of legumes (Poole et al., 2018). In addition to bulk and rhizosphere soil, other plant compartments and microhabitats impacted by soil bacteria and inoculants include the rhizoplane and endosphere (Clúa et al., 2018; Zhang et al., 2019).
Due to the presence of resident indigenous rhizobia in soil, difficulties are associated with introducing superior inoculant strains (Yates et al., 2011; Parnell et al., 2016; Chibeba et al., 2017) and elucidating the fate of the inoculated rhizobia using the 16S rRNA sequences typically employed in microbial community analyses. Similarly, the majority of other molecular methods cannot differentiate between added inoculant strains and the rhizobia already present in the resident population.
In the last few decades, some strains of Betaproteobacteria within the genera Burkholderia (Paraburkholderia) and Cupriavidus have been identified as symbionts of legumes, such as Mimosa pudica. While M. pudica and its symbiont (Paraburkholderia phymatum) are prevalent in South America and Asia, they have not yet been reported in the Midwestern United States (Gyaneshwar et al., 2011). Soils that lack specific rhizobia provide an opportunity to examine the interactions of an inoculant strain with indigenous bacteria and investigate root colonization in situ. A more detailed understanding of these interactions and the mechanisms involved may provide a means to enhance nodulation and subsequently N2 fixation by legumes in their non-native environments.
In the present study, we used a soil devoid of indigenous M. pudica-nodulating rhizobia to examine host-bacterial interactions in the absence of competition from nodulating strains. We further utilized this symbiosis to assess changes in the bacterial community of M. pudica in the rhizosphere, rhizoplane, and endosphere upon an inoculation with P. phymatum. The results obtained showed that while the population of P. phymatum increased in the rhizoplane of M. pudica following the soil inoculation, no significant changes were observed in the bacterial community structure in the soil. Consequently, a plant inoculation with the betaproteobacterium resulted in changes to a limited number of microhabitats.
Materials and Methods
Bacterial strains and growth conditions
P. phymatum strain MP20 was marked with β-glucuronidase (GUS) as previously described (Gehlot et al., 2013). The strain was maintained on Yeast Extract Mannitol (YEM) agar (Vincent, 1970). In inoculation studies, these strains were grown in YM broth at 28°C with shaking.
Plant inoculation and growth conditions
The nodulation of M. pudica by P. phymatum was performed as previously described (Gehlot et al., 2013). Briefly, M. pudica seeds (obtained from Outsidepride Seed Source, USA) were treated with concentrated sulfuric acid for 5 min, washed five times with sterile water, and germinated on sterile paper towels at 30°C. The soil used in the present study, a Kewaunee mesic Typic Hapludalfs, was obtained from the Cedar Grove Hawk Research Station State Natural Area located in Cedar Grove, WI, (43°33′35.8″N 87°47′55.0″W). To examine plant nodulation in soil, seedlings free of contamination were planted in Styrofoam cups containing a 1:1 ratio of vermiculite and Kewaunee soil. Three seedlings were grown in each cup. On day 2 after transplanting seedlings, three of the cups were inoculated with P. phymatum MP20-GUS (~106 bacteria per cup), and another three were used as uninoculated controls. Plants were incubated in a plant growth chamber at 26°C with a 14/10-h light/dark cycle. Plants were watered with modified Jensen’s nitrogen-free medium as needed. In the bacterial community analysis, seedlings were planted in a 1:1 ratio of vermiculite and the same soil, but in plastic trays with drainage. Each tray contained forty seedlings, and three trays were inoculated with independently grown P. phymatum MP20-GUS (~108 bacteria per tray). Plants were incubated as described above.
Plant colonization and nodulation
Plant colonization and nodulation were examined as previously described (Gehlot et al., 2013). Briefly, plants were removed from the soil mix 14 days after inoculation (DAI), and the roots were visually observed for nodule formation. Roots were stained for GUS activity using X-gluc (5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid) to confirm the presence of inoculated P. phymatum MP20-GUS. Stained nodules were observed and counted using a dissecting microscope.
Effects of the inoculant on soil and the rhizosphere microbiome
To examine the effects of the P. phymatum inoculation on the bacterial community structure, the rhizosphere and rhizoplane-endosphere soil, roots, and nodules were collected 0, 3, 7, and 14 DAI. Ten plants were taken for each time point (Fig. S1). Briefly, on 0 and 3 DAI, plants were gently removed from the pot, and closely adhering rhizosphere soil was removed with the aid of a sterile spatula. The roots were then separated from rhizosphere soil and homogenized in liquid nitrogen using a mortar and pestle to obtain rhizoplane plus endosphere samples. Regarding samples collected on 7 and 14 DAI, plants were gently pulled from the pot, and the attached rhizosphere soil was collected by gentle shaking and the use of forceps. Tightly adherent soil and the roots were homogenized to obtain rhizoplane samples (this fraction may also contain endospheric microbiota). Nodules were collected on 14 DAI, washed with sterile water, and homogenized. Ground samples were used to isolate DNA with the Qiagen® DNeasy PowerLyzer® PowerSoil® kit (Qiagen) according to manufacturer’s instructions
16 rRNA gene amplification and sequencing
In the analysis of microbial communities, the V4 hypervariable region of the 16S rRNA gene was amplified by PCR as previously described (Zhang et al., 2019). PCR conditions were 95°C for 5 min followed by 25 cycles at 98°C for 20 s, 55°C for 15 s, 72°C for 1 min, and a final extension at 72°C for 5 min. Sequencing was performed using barcoded primers and the dual indexing method at the University of Minnesota Genomics Center (UMGC, Minneapolis, MN, USA) (Gohl et al., 2016). Libraries were sequenced using the Illumina MiSeq platform (Illumina).
Analysis of bacterial diversity
The 300-nt sequences obtained were analyzed using the QIIME and R programs (McMurdie and Holmes, 2013). Briefly, raw sequence reads were trimmed and processed using SHI7 software (Al-Ghalith et al., 2018). Greengenes ver. 13.8 was used to align the processed sequences using NINJA-OPS (Al-Ghalith et al., 2018). Chloroplast and mitochondrial DNA sequences were removed using QIIME, and filtered data were further rarefied to 15,000 reads per sample for subsequent statistical analyses. Raw sequencing data were deposited in the NCBI Sequence Read Archive (SRA) under Accession Number PRJNA649756.
Microbial diversity values were assessed using alpha-diversity indices (e.g., observed species, Chao1, and Shannon) and were calculated from rarefied sequence data using the phyloseq package in R program. Differences in beta diversity were calculated based on Bray-Curtis dissimilarity matrices (Bray and Curtis, 1957). Non-metric multidimensional scaling (nMDS) was used to ordinate samples using metaMDS. Comparisons of communities between rhizosphere soils and roots were performed using ANOSIM. The influence of inoculated P. phymatum on the rhizosphere microbiome was analyzed using the Envift function in the vegan package (Oksanen et al., 2020).
Network analysis to identify potential keystone taxa
The co-occurrence network was generated as described by Zhang et al. (2019). Briefly, rare operational taxonomic units (OTUs, those with relative taxonomic abundance values <0.01%) were removed from samples, and the WGCNA package (Langfelder and Horvath, 2008) was used to construct the network using Spearman’s correlation matrix. The random matrix theory (RMT) was used to establish whether 0.791 was the appropriate similarity threshold (Luo et al., 2006). Additionally, to avoid type I errors in the correlation matrix, P values were further adjusted using the Benjamini and Hochberg false discovery rate (FDR) of <0.05 (Luo et al., 2006). Network properties were calculated using igraph (Csárdi and Nepusz, 2006). Potential keystone taxa were identified as follows: OTUs with degree >5, closeness centrality >0.15, and centrality <0.035 (Berry and Widder, 2014). The network image was generated using Cytoscape (Shannon et al., 2003).
Results and Discussion
Nodulation of M. pudica by P. phymatum MP20-GUS in soil
To establish whether agricultural soil collected from a field in Cedarburg WI contained rhizobia that nodulate M. pudica, farm soil was used as an inoculum for M. pudica seedlings, and nodulation was examined after a four-week incubation. No nodules formed on M. pudica roots following the soil inoculation (Fig. 1B), indicating the absence of rhizobia that formed a symbiotic relationship with M. pudica. These results are consistent with the known native and non-native range of M. pudica that does not include the Midwestern USA (Gyaneshwar et al., 2011).
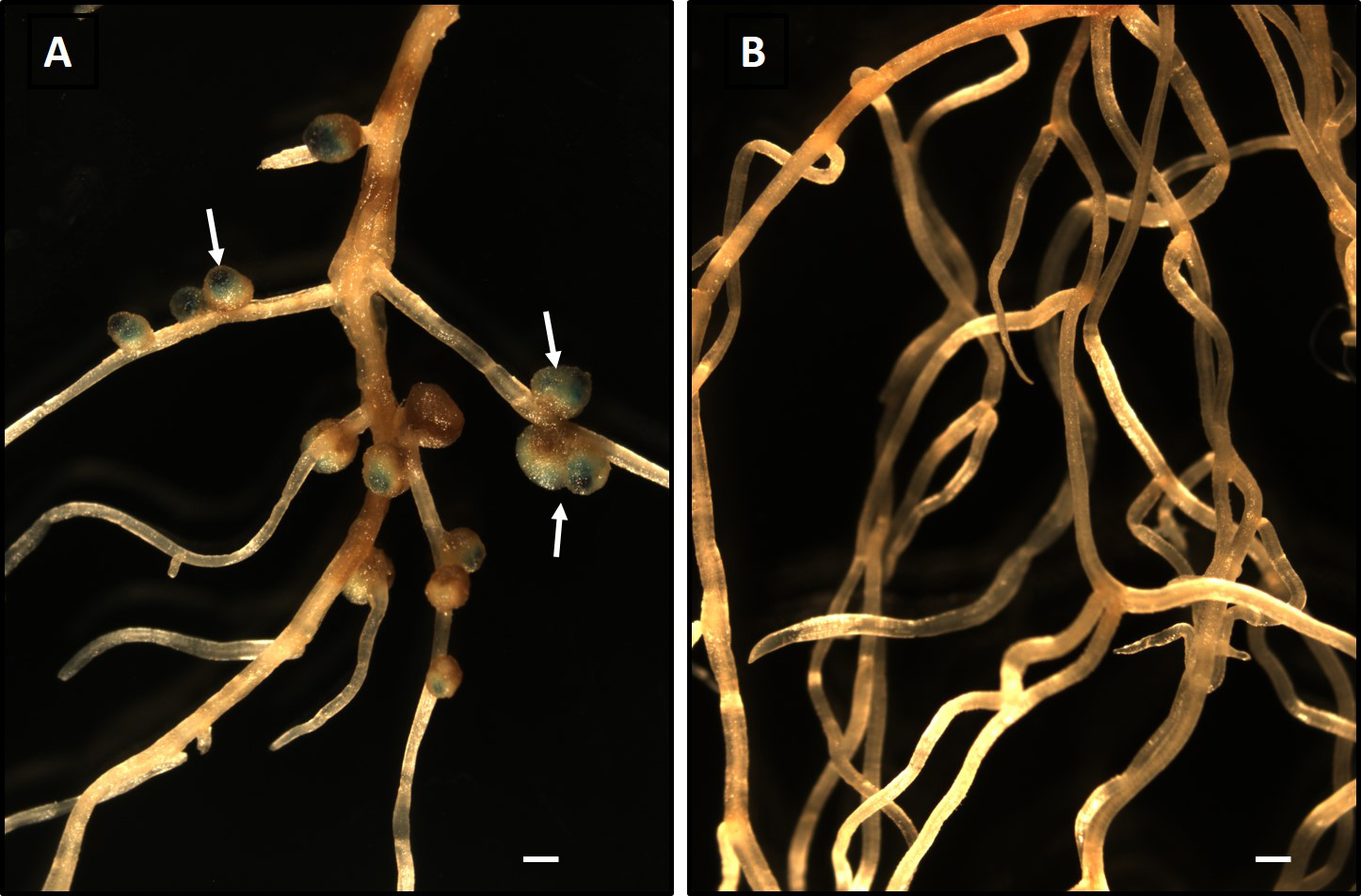
To investigate whether a known symbiont of M. pudica competes with microbes in non-native soil and successfully colonizes and nodulates a host legume, we inoculated soil-grown seedlings with approximately 1×103
P. phymatum strain MP20-GUS (Gehlot et al., 2013). This bacterium was previously shown to form nodules on M. pudica (Gehlot et al., 2013). The inoculation of M. pudica with P. phymatum MP20-GUS resulted in the formation of nodules throughout the root system (Fig. 1A). To confirm that nodules were formed by the inoculated strain, roots were stained for GUS activity using X-gluc. Blue staining due to GUS was observed in the nodules (Fig. 1A). These results indicated that P. phymatum MP20-GUS colonized and formed a symbiotic relationship with M. pudica in soil. The colonization of a host plant in non-sterile soils will have to compete with native microorganisms for resources that are present in the plant rhizosphere and recognize signaling molecules in the presence of other microorganisms. The ability of P. phymatum to efficiently nodulate M. pudica in non-native soil was expected because this bacterium was previously identified as a major symbiont of M. pudica in the Indian subcontinent in which M. pudica is highly invasive (Gehlot et al., 2013). These findings suggest that M. pudica-P. phymatum symbiosis in non-native soils may be developed further as a model system to obtain a more detailed understanding of plant-bacterial interactions in situ rather than under axenic artificial plant growth conditions.
Microbial community changes in M. pudica rhizosphere and rhizoplane soils following the inoculation.
The soils comprising the plant rhizosphere and rhizoplane are rich in nutrients and, thus, contain a great diversity of microorganisms. Previous studies estimated that 1 g of soil contained >10,000–50,000 bacterial species (Roesch et al., 2007; Berendsen et al., 2012; Turner et al., 2013). To establish whether an inoculation with P. phymatum leads to specific changes in the resident soil bacterial community surrounding M. pudica, we collected rhizosphere and rhizoplane+endosphere soils from the roots of soil-grown M. pudica plants with or without the inoculation with P. phymatum. The bacterial community structure and dynamics were examined using a 16S rRNA gene sequence analysis.
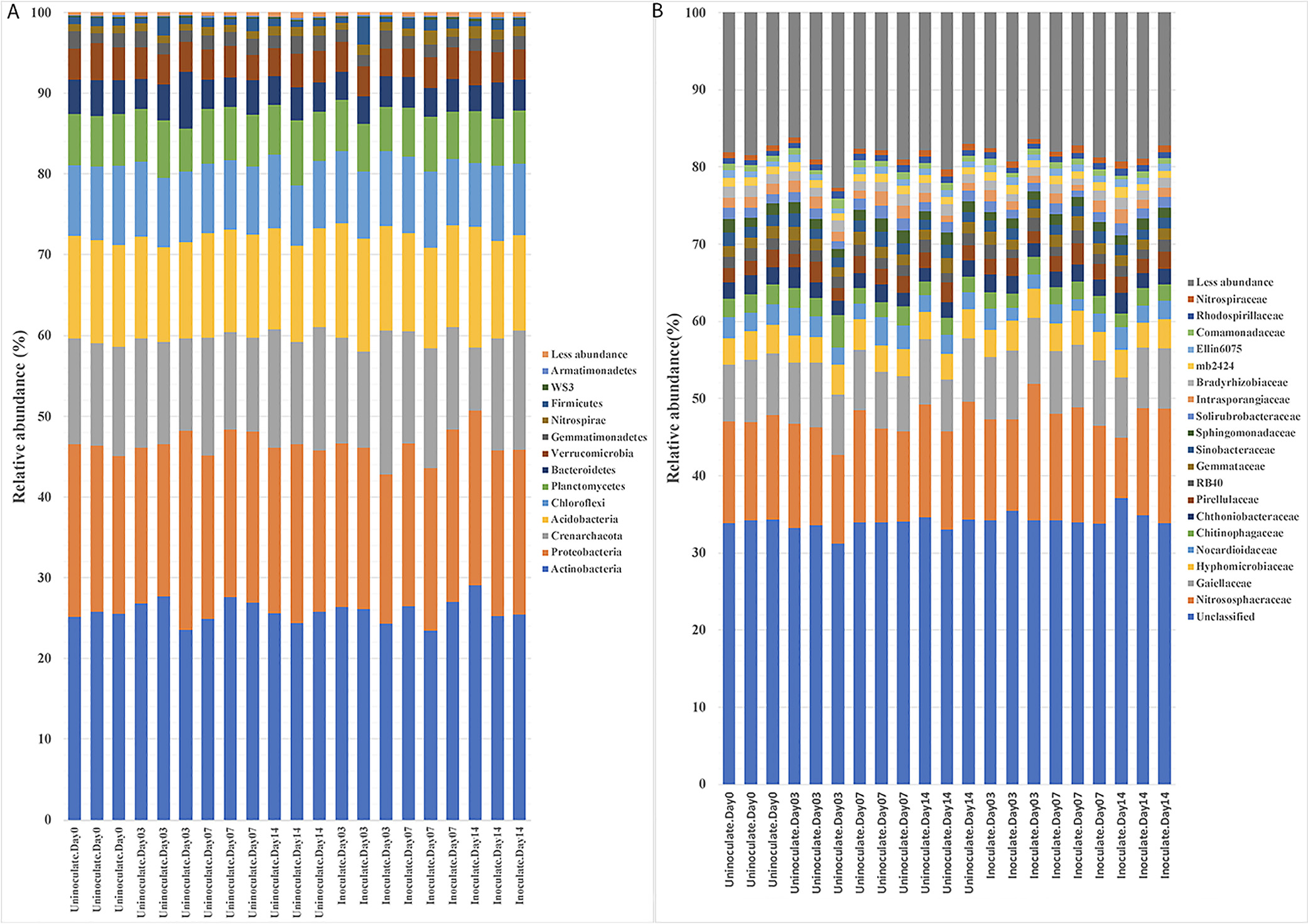
In uninoculated plants, the bacterial community associated with the rhizoplane showed changes on different days of growth. While Proteobacteria decreased from 70.2 to 48.2% during the two-week incubation, the amounts of Actinobacteria, Acidobacteria, Crenarchaeota, Chloroflexi, Verrucomicrobia, and Planctomycetes significantly increased (Fig. 2A). Previous studies reported that the root microbiome depends on the soil and plant species (Lundberg et al., 2012; Bulgarelli et al., 2013; Tkacz and Poole, 2015). Consequently, the changes observed in the bacterial community during two weeks of growth were consistent with previous findings showing that the plant exerts control over its microbiota via the secretion of specific metabolites from the roots, which change with the growth of the plant (Bais et al., 2006; Lundberg et al., 2012; Bulgarelli et al., 2013).

The inoculation with P. phymatum induced a significant shift in the OTUs present in rhizoplane+endosphere samples (Fig. 2B) from those in uninoculated plants. Moreover, OTUs representing 13 different taxa showed significant differences in relative abundance (Fig. 2B). Specifically, OTUs corresponding to Oxalobacteraceae, Enterobacteriaceae, Pseudomonadaceae, and Sphingobacteriaceae showed a significant decrease over the two-week period, whereas OTUs representing Rhizobiaceae, Nitrososphaeraceae, and Hyphomicrobiaceae increased (Fig. 2B).
In contrast to the changes observed in rhizoplane soil, the inoculation did not induce marked changes in the dynamics of OTUs in the rhizosphere soil (Fig. 3). The most abundant OTUs belonged to Actinobacteria (25.88±1.40%), Proteobacteria (20.56±1.29%), Acidobacteria (12.67±0.81%), and Crenarchaeota (13.28±1.92%), (Fig. 3A). Similarly, no major changes due to the P. phymatum inoculation were observed when OTUs were characterized at the family level. These diverse OTUs belonged to 19 major families and were dominated by Nitrosophaeraceae, Gaiellaceae, and Hyphomicrobiaceae at 13.3±1.9, 7.9±0.5, and 3.6±0.3%, respectively (Fig. 3B).
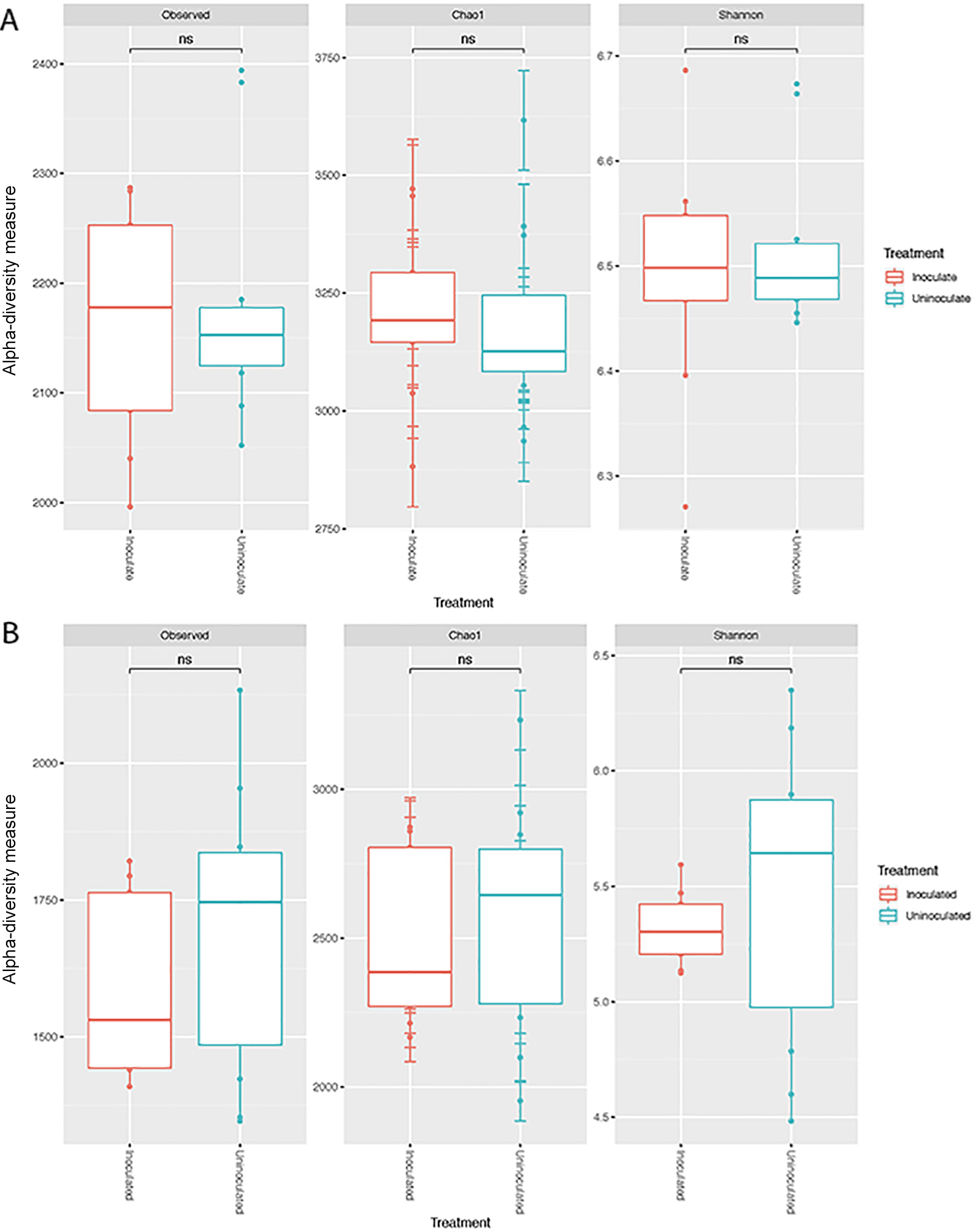
Although not significantly different (P>0.05), alpha-diversity increased in rhizosphere samples, but decreased in the rhizoplane as a result of the inoculation. This is consistent with previous findings showing that microbial communities were less diverse in the rhizoplane/roots than in bulk and rhizosphere soils. This is mainly considered to be due to the release of specific metabolites in root exudates (Berendsen et al., 2012; Lundberg et al., 2012). No significant differences were observed in alpha-diversity between uninoculated and inoculated soils and root samples (Fig. 4A and B).
Non-metric multidimensional scaling (NMDS), which represents pairwise dissimilarity between objects, revealed that the inoculation led to the significant separation of communities in rhizoplane+endosphere samples. In contrast, the inoculation did not induce any significant changes in microbial communities in the rhizospheric soil. This was further confirmed using ANOSIM based on an analysis of genera by treatment. The results of the analysis showed that microbial communities were not significantly affected by the P. phymatum inoculation (R2=0.18, P=0.17) in the rhizosphere soil (Fig. 5A), but were significantly separated in rhizoplane/endosphere samples (R2=0.86, P=0.001) (Fig. 5B).
Growth of P. phymatum in soil
The majority of microbial inoculants are generally assumed to multiply in the rhizosphere before colonizing the roots of the plant host. To examine this in more detail, we focused our attention on Burkholderia/Paraburkholderia OTUs. The results of this analysis (Fig. 6A) showed that Burkholderia OTUs significantly decreased over time in the rhizosphere soil from 0.4% at the time of the inoculation to 0.08% by 14 DAI. In contrast, Burkholderia OTUs significantly increased in rhizoplane+endosphere samples from 1.2 to 17.4% over the same period (Fig. 6B).
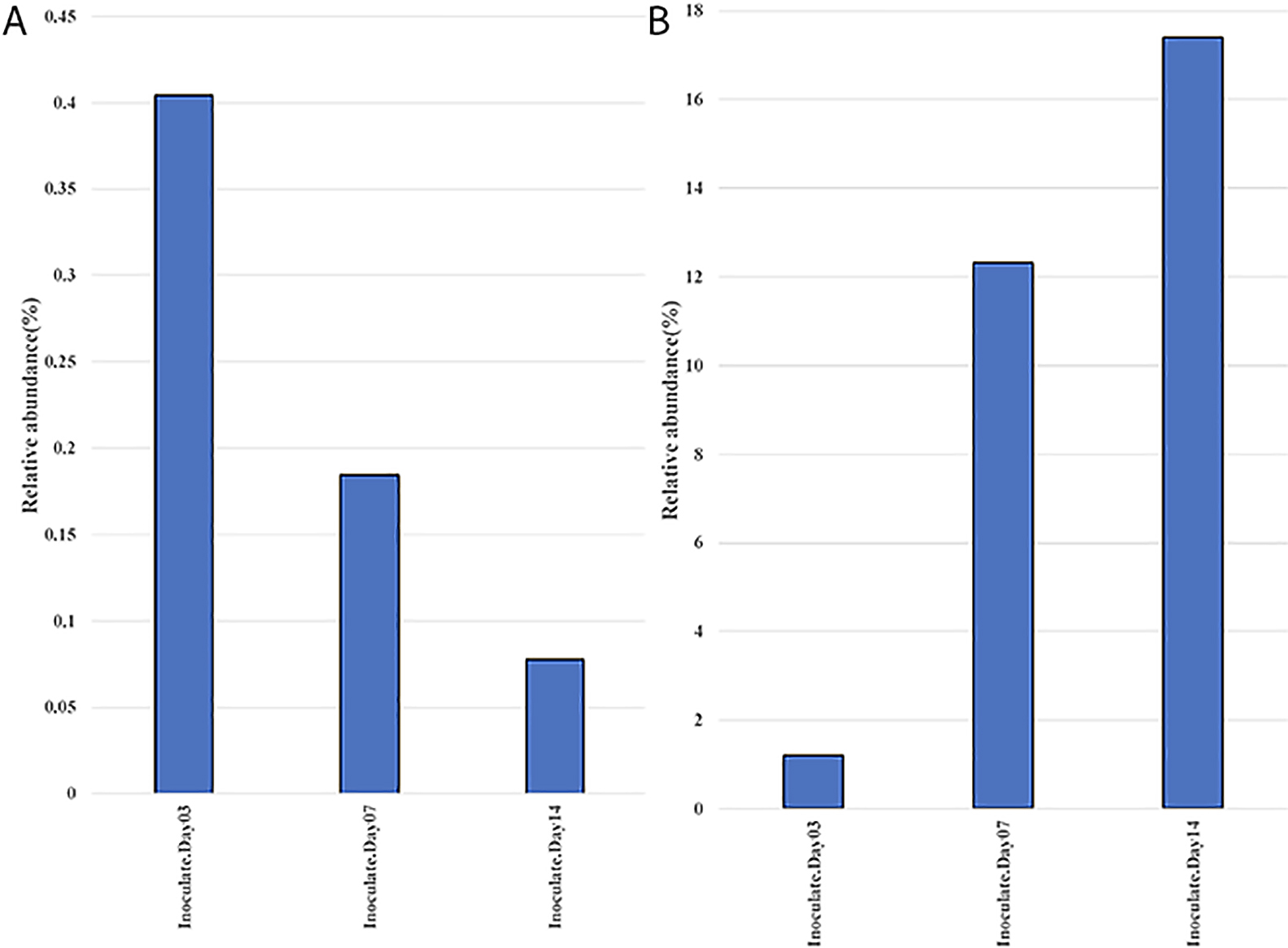
These results suggest that the colonization of the rhizoplane and roots is important for rhizobial competition and growth, more so than just growth in the rhizosphere, which may occur far from the root proper. Root attachment is considered to be most important factor for rhizobial proliferation (Wheatley and Poole, 2018). Root colonization requires the migration of bacteria towards root exudates and the P. phymatum genome contains 113 genes annotated as being involved in chemotaxis and motility (Moulin et al., 2014), which were differentially expressed in the presence of M. pudica root exudates (Klonowska et al., 2018). P. phymatum may utilize M. pudica root exudates as nutrient sources more effectively than resident microbes once it colonizes and attaches to the root. A recent study using transcriptomics showed that root exudates of M. pudica induced P. phymatum genes involved in transport and the utilization of aromatic compounds, inositol, and organic acids (Klonowska et al., 2018). The ability to metabolize myo-inositol was previously shown to be important for root colonization and the nodulation of pea and vetch by Rhizobium leguminosarum bv. viciae (Fry et al., 2001). Further studies are needed to elucidate the role of these catabolic pathways in the ability of P. phymatum to compete with native microorganisms and nodulate M. pudica in soil.
Identification of keystone bacteria affected by the inoculation of P. phymatum
Microbial communities possess different metabolic capabilities and are involved in beneficial and/or competitive interactions with each other. These diverse activities and microbial interactions form functional networks between the microbes and their biotic and abiotic environments. Within these functional networks, certain species and strains are considered to be keystone and have been postulated to play crucial roles in the functional ecology of the environment (Barberán et al., 2012; Bulgarelli et al., 2012; van der Heijden and Hartmann, 2016). Since the present results showed that the inoculation of M. pudica with P. phymatum significantly affected the microbial communities associated with root surfaces, we attempted to clarify whether keystone species were associated with these changes. Therefore, we analyzed and compared the bacterial network and keystone taxa associated with inoculated and uninoculated roots using co-occurrence network analyses (Berry and Widder, 2014; Banerjee et al., 2018). The network of co-occurring OTUs was comprised of 924 nodes and 1,330 edges. The keystone taxa within each network of positive co-occurrences were comprised of OTUs belonging to two different genera: Streptomyces and Phycicoccus (Fig. 7). Members of Streptomyces significantly increased from 0.079% in the uninoculated treatment to 0.113% in the inoculated treatment (Tukey’s HSD, P<0.05), while those of Phycicoccus significantly decreased from 0.101 to 0.061% (Fig. 7). The increase observed in Streptomyces may be of ecological significance for nodulation by P. phymatum because these bacteria synthesize and secrete diverse secondary metabolites that include many antibiotics (van Kulen and Dyson, 2014). In the plant rhizosphere, Streptomyces spp. have been shown to produce antifungal and plant growth-promoting metabolites and have potential as inoculants for enhancing agriculture production (Schrey and Tarkka, 2008; Rey and Dumas, 2017). Further studies are needed to clarify the direct/indirect effects of Streptomyces on legume nodulation in non-sterile soils.

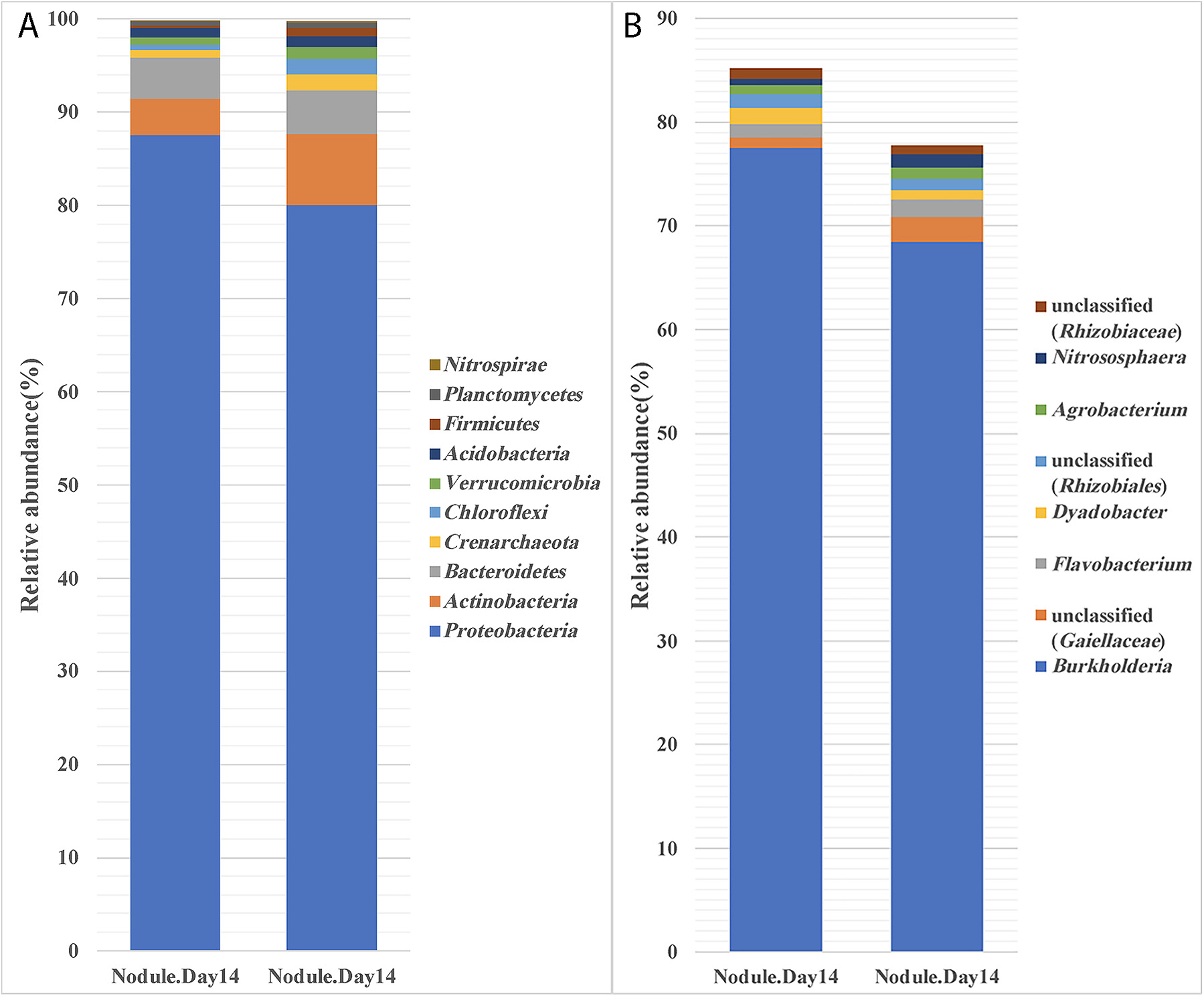
Previous studies showed that bacteria unrelated to legume-nodulating rhizobia may be isolated from surface-sterilized nodules, indicating that other bacteria also possess the ability to infect legume nodules (Deng et al., 2011; Aserse et al., 2013; Martinez-Hidalgo and Hirsch, 2017). Although these bacteria do not nodulate under axenic conditions, they may be important for symbiosis in soil environments. Studies conducted to date have relied on the isolation of culturable bacteria from nodules. Since the majority of bacteria are not cultured in the laboratory, the extent of the non-rhizobial occupants of nodules in soils remains unclear. In the present study, we analyzed the 16S rRNA diversity of M. pudica nodules that formed following an inoculation with P. phymatum. While the results obtained demonstrated that the majority (~75%) of nodule-occupant OTUs were from Burkholderia, which was expected, we also detected OTUs from Agrobacterium, Rhizobiales, Nitrososphaera, and Flavobacterium in the homogenates of surface-sterilized nodules (Fig. 8B). Since no nodules were detected on M. pudica in the absence of the P. phymatum inoculation, these bacteria do not have the ability to nodulate M. pudica. Therefore, they may have entered M. pudica nodules via a co-infection mechanism. This phenomenon is now well recognized (Muresu et al., 2008; Martinez-Hidalgo and Hirsch, 2017). However, further studies are needed to examine the role played by these co-infecting microbes.
Conclusion
We examined the effects of an inoculation of P. phymatum on changes in the microbial community in the rhizoplane, roots, and nodules in natural agricultural soil. An analysis using the 16S rRNA gene showed that P. phymatum did not significantly change the community of the rhizospheric soil. In contrast, the population of P. phymatum significantly increased in root samples. A network analysis identified Streptomyces spp. as keystone taxa that may be involved in nodulation by P. phymatum.
Citation
Welmillage, S. U., Zhang, Q., Sreevidya, V. S., Sadowsky, M. J., and Gyaneshwar, P. (2021) Inoculation of Mimosa Pudica with Paraburkholderia phymatum Results in Changes to the Rhizoplane Microbial Community Structure. Microbes Environ 36: ME20153.
https://doi.org/10.1264/jsme2.ME20153
Acknowledgements
This study was supported in part by funding from the RACAS grant of the University of Wisconsin, Milwaukee (to PG) and from the Minnesota Agricultural Experiment Station (to MJS)
References
- Al-Ghalith, G.A., Hillmann, B., Ang, K., Shields-Cutler, R., and Knights, D. (2018) SHI7 is a self-learning pipeline for multipurpose short-read DNA quality control. mSystems
3: e00202-17.
- Aserse, A.A., Räsänen, L.A., Aseffa, F., Hailemariam, A., and Lindström, K. (2013) Diversity of sporadic symbionts and nonsymbiotic endophytic bacteria isolated from nodules of woody, shrub, and food legumes in Ethiopia. Appl Microbiol Biotechnol
97: 10117–10134.
- Bais, H.P., Weir, T.L., Perry, L.G., Gilroy, S., and Vivanco, J.M. (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol
57: 233–266.
- Banerjee, S., Schlaeppi, K., and van der Heijden, M.G.A. (2018) Keystone taxa as drivers of microbiome structure and functioning. Nat Rev Microbiol
16: 567–576.
- Barberán, A., Bates, S.T., Casamayor, E.O., and Fierer, N. (2012) Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J
6: 343–351.
- Berendsen, R.L., Pieterse, C.M.J., and Bakker, P. (2012) The rhizosphere microbiome and plant health. Trends Plant Sci
17: 478–486.
- Berry, D., and Widder, S. (2014) Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front Microbiol
5: 219.
- Bray, J.R., and Curtis, J.T. (1957) An ordination of the upland forest communities of southern Wisconsin. Ecol Monog
27: 325–349.
- Bulgarelli, D., Rott, M., Schlaeppi, K., van Themaat, E.V.L., Ahmadinejad, N., Assenza, F., et al. (2012) Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature
488: 91–95.
- Bulgarelli, D., Schlaeppi, K., Spaepen, S., Ver Loren van Themaat, E., and Schulze-Lefert, P. (2013) Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol
64: 807–838.
- Chibeba, A.M., Kyei-Boahen, S., Guimarães, M.F., Nogueira, M.A., and Hungria, M. (2017) Isolation, characterization and selection of indigenous Bradyrhizobium strains with outstanding symbiotic performance to increase soybean yields in Mozambique. Agric, Ecosyst Environ
246: 291–305.
- Clúa, J., Roda, C., Zanetti, E.M., and Blanco, A.F. (2018) Compatibility between legumes and Rhizobia for the establishment of a successful nitrogen-fixing symbiosis. Genes
9: 125.
- Csárdi, G., and Nepusz, T. (2006) The igraph software package for complex network research. J Comp Appl
29: 2191–2193.
- Deaker, R., Roughley, R.J., and Kennedy, I.R. (2004) Legume seed inoculation technology—A review. Soil Biol Biochem
36: 1275–1288.
- Deng, Z.S., Zhao, L.F., Kang, Z.Y., Yang, W.Q., Lindström, K., Wang, E.T., and Wei, G.H. (2011) Diversity of endophytic bacteria within nodules of the Sphaerophysa salsula in different regions of Loess Plateau in China. FEMS Microbiol Ecol
76: 463–475.
- Fry, J., Wood, M., and Poole, P.S. (2001) Investigation of myo-inositol catabolism in Rhizobium leguminosarum bv. viciae and its effect on nodulation competitiveness. Mol Plant Microbe Interact
14: 1016–1025.
- Gehlot, H.S., Tak, N., Kaushik, M., Mitra, S., Chen, W.M., Poweleit, N., et al. (2013) An invasive Mimosa in India does not adopt the symbionts of its native relatives. Ann Bot
112: 179–196.
- Gohl, D., Vangay, P., Garbe, J., MacLean, A., Hauge, A., Becker, A., et al. (2016) Systematic improvement of amplicon marker gene methods for increased accuracy in microbiome studies. Nat Biotechnol
34: 942–949.
- Gyaneshwar, P., Hirsch, A.M., Moulin, L., Chen, W.M., Elliott, G.N., Bontemps, C., et al. (2011) Legume-nodulating betaproteobacteria: Diversity, host range and future prospects. Mol Plant-Microbe Interact
24: 1276–1288.
- Jones, D.L., Nguyen, C., and Finlay, R.D. (2009) Carbon flow in the rhizosphere: Carbon trading at the soil-root interface. Plant Soil
321: 5–33.
- Klonowska, A., Melkonian, R., Miché, L., Tisseyre, P., and Moulin, L. (2018) Transcriptomic profiling of Burkholderia phymatum STM815, Cupriavidus taiwanensis LMG19424 and Rhizobium mesoamericanum STM3625 in response to Mimosa pudica root exudates illuminates the molecular basis of their nodulation competitiveness and symbiotic evolutionary history. BMC Genomics
19: 105.
- Langfelder, P., and Horvath, S. (2008) WGCNA: An R package for weighted correlation network analysis. BMC Bioinformatics
9: 559.
- Lundberg, D.S., Lebeis, S.L., Paredes, S.H., Yourstone, S., Gehring, J., Malfatti, S., et al. (2012) Defining the core Arabidopsis thaliana root microbiome. Nature
488: 86–90.
- Luo, F., Zhong, J., Yang, Y., Scheuermann, R., and Zhou, J. (2006) Application of random matrix theory to biological networks. Phys Lett A
357: 420–423.
- Martinez-Hidalgo, P., and Hirsch, A.M. (2017) The nodule microbiome; N2-fixing rhizobia do not live alone. Phytobiomes J
1–52.
- McMurdie, P.J., and Holmes, S. (2013) phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One
8: e61217.
- Moulin, L., Klonowska, A., Caroline, B., Booth, K., Vriezen, J.A.C., Melkonian, R., et al. (2014) Complete Genome sequence of Burkholderia phymatum STM815T, a broad host range and efficient nitrogen-fixing symbiont of Mimosa species. Stand Genomic Sci
9: 763–774.
- Muresu, R., Polone, E., Sulas, L., Baldan, B., Tondello, A., Delogu, G. et al. (2008) Coexistence of predominantly nonculturable rhizobia with diverse, endophytic bacterial taxa within nodules of wild legumes. FEMS Microbiol Ecol
63: 383–400.
- Narożna, D., Pudełko, K., Króliczak, J., Goliñska, B., Sugawara, M., Mkadrzak, C.J., et al. (2015) Survival and competitiveness of Bradyrhizobium japonicum strains 20 years after introduction into field locations in Poland. Appl Environ Microbiol
81: 5552–5559.
- O’Hara, G.W. (1998) The role of nitrogen fixation in crop production. J Crop Prod
1: 115–138.
- Oksanen, A.J., Blanchet, F.G., Friendly, M., Kindt, R., Legendre, P., Mcglinn, D., et al. (2020) Package ‘vegan’ Community Ecology Package Version 2.5–7. URL https://CRAN.R-project.org/package=vegan
- Oldroyd, G.E.D., Murray, J.D., Poole, P.S., and Downie, J.A. (2011) The rules of engagement in the legume-rhizobial symbiosis. Annu Rev Genet
45: 119–144.
- Oldroyd, G.E.D. (2013) Speak, friend and enter: Signaling systems that promote beneficial symbiotic associations in plants. Nat Rev Microbiol
11: 252–263.
- Parnell, J.J., Berka, R., Young, H.A., Sturino, J.M., Kang, Y., Barnhart, D.M., and DiLeo, M.V. (2016) From the lab to the farm: An industrial perspective of plant beneficial microorganisms. Front Plant Sci
7: 1110.
- Poole, P., Ramachandran, V., and Terpolilli, J. (2018) Rhizobia: from saprophytes to endosymbionts. Nat Rev Microbiol
16: 291–303.
- Rey, T., and Dumas, B. (2017) Plenty is no plague: Streptomyces symbiosis with crops. Trends Plant Sci
22: 30–37.
- Roesch, L., Fulthorpe, R., Riva, A., Casella, G., Hadwin, A.K.M., Kent, A.D., et al. (2007) Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J
1: 283–290.
- Schrey, S.D., and Tarkka, M. (2008) Friends and Foes: streptomyces as modulators of plant disease and symbiosis. Antonie van Leeuwenhoek
94: 11–19.
- Shannon, P., Markiel, A., Ozier, O., Baliga, N.S., Wang, J.T., Ramage, D., et al. (2003) Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res
13: 2498–2504.
- Sprent, J.I., and James, E.K. (2007) Legume evolution: Where do nodules and mycorrhizas fit in?
Plant Physiol
144: 575–581.
- Tkacz, A., and Poole, P.S. (2015) The role of microbiota in plant productivity. J Exp Bot
66: 2167–2175.
- Triplett, E.W., and Sadowsky, M. (1992) Genetics of competition for nodulation of legumes. Annu Rev Microbiol
43: 399–428.
- Turner, T.R., James, E.K., and Poole, P.S. (2013) The plant microbiome. Genome Biol
14: 209.
- van der Heijden, M.G., and Hartmann, M. (2016) Networking in the plant microbiome. PLoS Biol
14: e1002378.
- van Keulen, G., and Dyson, P.J. (2014) Production of specialized metabolites by Streptomyces coelicolor A3(2). Adv Appl Microbiol
89: 217–266.
- Venkateshwaran, M., Volkening, J.D., Sussman, M.R., and Ane, J.M. (2013) Symbiosis and the social network of higher plants. Curr Opin Plant Biol
16: 118–127.
- Vincent, J.M. (1970) A Manual for the Practical Study of Root Nodule Bacteria. I.B.P Handbook No: 15. Oxford, UK: International Biological Programme, Blackwell Science.
- Walker, T.S., Bais, H.P., Grotewold, E., and Vivanco, J.M. (2003) Root exudation and rhizosphere biology. Plant Physiol
132: 44–51.
- Wheatley, R.M., and Poole, P.S. (2018) Mechanisms of bacterial attachment to roots. FEMS Microbiol Rev
42: 448–461.
- Yates, R.J., Howieson, J.G., Reeve, W.G., and O’Hara, G.W. (2011) A re-appraisal of the biology and terminology describing rhizobial strain success in nodule occupancy of legumes in agriculture. Plant Soil
348: 255–267.
- Zhang, Q., Acuña, J.J., Inostroza, N.G., Mora, M.L., Radic, S., Sadowsky, M.J., et al. (2019) Endophytic bacterial communities associated with roots and leaves of plants growing in Chilean extreme environments. Sci Rep
9: 4950.
