Abstract
Anammox bacteria produce N2 gas by oxidizing NH4+ with NO2–, and hydroxylamine (NH2OH) is a potential intermediate of the anammox process. N2 gas production occurs when anammox bacteria are incubated with NH2OH only, indicating their capacity for NH2OH disproportionation with NH2OH serving as both the electron donor and acceptor. Limited information is currently available on NH2OH disproportionation by anammox bacteria; therefore, the stoichiometry of anammox bacterial NH2OH disproportionation was examined in the present study using 15N-tracing techniques. The anammox bacteria, Brocadia sinica, Jettenia caeni, and Scalindua sp. were incubated with the addition of 15NH2OH, and the production of 15N-labeled nitrogenous compounds was assessed. The anammox bacteria tested performed NH2OH disproportionation and produced 15-15N2 gas and NH4+ as reaction products. The addition of acetylene, an inhibitor of the anammox process, reduced the activity of NH2OH disproportionation, but not completely. The growth of B. sinica by NH2OH disproportionation (–240.3 kJ mol NH2OH–1 under standard conditions) was also tested in 3 up-flow column anammox reactors fed with 1) 0.7 mM NH2OH only, 2) 0.7 mM NH2OH and 0.5 mM NH4+, and 3) 0.7 mM NH2OH and 0.5 mM NO2–. NH2OH consumption activities were markedly reduced after 7 d of operation, indicating that B. sinica was unable to maintain its activity or biomass by NH2OH disproportionation.
Anaerobic ammonium-oxidizing (anammox) bacteria were initially discovered in a denitrifying bioreactor in 1995 (Mulder et al., 1995), and are now recognized as the main players in the global nitrogen cycle (Kuypers et al., 2003; 2005; Amano et al., 2007; Yoshinaga et al., 2011). In the anammox process, NH4+ is oxidized to N2 gas using NO2– as an electron acceptor, and the stoichiometry of the anammox process has been described as follows (Lotti et al., 2014):
1 NH4++1.146 NO2–+0.071 HCO3–+0.057 H+
→ 0.986 N2+0.161 NO3–+0.071 CH1.74O0.31N0.20+2.002 H2O eq. 1
As an intermediate, hydrazine (N2H4) is synthesized from NH4+ and NO or NH2OH by hydrazine synthase, and its biosynthesis appears to be unique to metabolism by anammox bacteria (Strous et al., 2006; Kartal et al., 2011; Oshiki et al., 2016a). Anammox bacteria are monophyletically affiliated into the bacterial order Brocadiales in the phylum Planctomycetota (Strous et al., 1999), and five candidate genera have been proposed: Candidatus Brocadia, Kuenenia, Jettenia, Anammoxoglobus, and Scalindua (Egli et al., 2003; Kuypers et al., 2003; Quan et al., 2008). These anammox bacteria have been detected in various natural and man-made ecosystems and are significantly involved in nitrogen loss (Oshiki et al., 2016b).
Hydroxylamine (NH2OH), a well-known intermediate of the aerobic NH3 oxidation reaction (Madigan et al., 2019), is also a potential intermediate of the anammox process (van de Graaf et al., 1997). The anammox bacterium, Brocadia sinica reduced NO2– to NH2OH (Oshiki et al., 2016a) potentially by using reductive hydroxylamine dehydrogenase (rHao) (Ferousi et al., 2021), and synthesized N2H4 from the NH2OH and NH4+ using hydrazine synthase (Oshiki et al., 2016a). Kuenenia stuttgartiensis reduced NO2– to NO (Kartal et al., 2011), and NO was further reduced to NH2OH by hydrazine synthase to synthesize N2H4 (Dietl et al., 2015). Apart from the anammox process, the nitrogen transformation reaction, NH2OH disproportionation, has been reported in anammox bacteria (van der Star et al., 2008). A disproportionation reaction is a reaction in which a chemical compound serves as both an electron donor and accepter, and the disproportionation reaction of inorganic sulfur (Finster, 2008) is an example of a microbial disproportionation reaction. In NH2OH disproportionation, NH2OH is converted to N2 gas and NH4+ using the following stoichiometry (Pacheco et al., 2011):
3 NH2OH+H+ → NH4++N2+3 H2O eq. 2
NH2OH disproportionation involves the following 2 reactions, N2H4 production and consumption (Soler-Jofra et al., 2020):
NH4++NH2OH → N2H4+H2O+H+ eq. 3
2 NH2OH+N2H4+2 H+ → 2 NH4++N2+2 H2O eq. 4
Although NH2OH disproportionation by K. stuttgartiensis (van der Star et al., 2008; Soler-Jofra et al., 2020) and B. sinica (Oshiki et al., 2016a) has been described, further studies are required to obtain a more detailed understanding of anammox bacterial NH2OH disproportionation for the following reasons. Although the kinetics of NH2OH disproportionation have been investigated using K. stuttgartiensis (van der Star et al., 2008; Soler-Jofra et al., 2020), the amounts of N2 gas produced in NH2OH disproportionation (see eq. 2) were not measured in previous studies and the stoichiometry of NH2OH disproportionation was not established. Furthermore, we previously examined NH2OH disproportionation by B. sinica; however, we only reported the occurrence of NH2OH disproportionation (Oshiki et al., 2016a) and did not investigate the effects of NH2OH concentrations on NH2OH disproportionation by repeating batch incubations. In addition, although eq. 2 yields –240.3 kJ mol NH2OH–1 of free energy under standard conditions (Soler-Jofra et al., 2020), the growth of anammox bacteria with NH2OH disproportionation has not yet been examined.
Therefore, the present study investigated anammox bacterial NH2OH disproportionation. The phylogenetically different anammox bacteria, B. sinica, Jettenia caeni, and Scalindua sp. were incubated with 15N-labeled NH2OH, and the stoichiometry of NH2OH disproportionation was carefully assessed based on measurements of 15-15N2 and NH4+ concentrations. NH2OH disproportionation was also analyzed under acetylene inhibition conditions. Acetylene is a strong inhibitor of aerobic NH3 oxidation, N2O reduction to N2 (i.e., denitrification), and the anammox process (Jensen et al., 2007); however, its effects on NH2OH disproportionation currently remain unknown. B. sinica was cultured in up-flow column reactors with the addition of NH2OH to establish whether it grows on NH2OH, and the activity of NH2OH consumption and the abundance of the anammox bacterial 16S rRNA gene were evaluated.
Materials and Methods
Anammox bacterial cultures
Planktonic cells of B. sinica, J. caeni, and Scalindua sp. were cultivated in membrane bioreactors (MBRs) equipped with a hollow fiber membrane module (pore size of 0.1 μm, polyethylene) as previously described (Oshiki et al., 2013; Zhang and Okabe, 2020). Culture media fed into MBRs contained KH2PO4 (24.4 mg L–1), MgSO4·7H2O (60 mg L–1), CaCl2 (51 mg L–1), yeast extract (Becton, Dickinson and Company) (1.0 mg L–1), and 0.5 mL of trace element solutions I and II (van de Graaf et al., 1996). The artificial sea salt SEALIFE (Marine Tech) (Kindaichi et al., 2011) was supplemented into media for “Ca. Scalindua sp.” at a final concentration of 28 g L–1. Equimolar amounts of NH4(SO4)2 and NaNO2 were supplemented into media at 10 mM for B. sinica and Scalindua sp. and 5 mM for J. caeni, and the nitrogen loading rates of the MBRs for B. sinica, J. caeni, and Scalindua sp. were set at 0.55, 0.18, and 0.45 kg N m–3 d–1, respectively. MBRs were operated at 37°C for B. sinica and at 25°C for J. caeni and Scalindua sp. pH was not controlled in MBRs, but was in the range of pH 7.6–8.0. Anammox bacterial cells accounted for more than 90% of the total biomass in MBRs as measured by a fluorescence in-situ hybridization (FISH) analysis using the oligonucleotide probes AMX820 (Schmid et al., 2001) and EUBmix composed of equimolar EUB338, EUB338II, and EUB338III (Daims et al., 1999). Anammox bacterial species were routinely checked based on the partial anammox bacterial 16S rRNA gene sequence using Sanger sequencing (Oshiki et al., 2011).
Batch incubations of anammox bacteria
Standard anaerobic techniques were employed in an anaerobic chamber (Coy Laboratories Products) in which the concentration of oxygen was maintained at <1 ppm. Culture media and stock solutions were prepared by purging N2 gas for >30 min, and then repeatedly vacuuming and purging He gas. The 15N enrichment of 15NH2OH·HCl (Cambridge Isotope Laboratories) was >98%.
Anammox bacterial cells collected from MBRs were centrifuged at 13,420×g at 20°C for 10 min, washed, and then resuspended in the above culture media without NH4+ and NO2– at concentrations of 0.5 mg protein mL–1. Twenty-five milliliters of the cell suspension was dispensed into 70-mL serum glass vials (Nichiden-Rika glass), and the headspace was replaced with He gas (>99.99995%) after sealing with butyl rubber stoppers and aluminium caps. Vials were incubated after the addition of 15NH2OH (final concentration of 1.0 to 10 mM) and acetylene (30 μM) (Jensen et al., 2007) in the dark at 37°C for B. sinica and at 25°C for J. caeni and Scalindua sp.. Liquid samples were collected using a 1-mL plastic disposable syringe, immediately filtered using a 0.2-μm cellulose acetate filter, and subjected to measurements of NH4+, NO2–, NO3–, and NH2OH concentrations. Gas samples were collected using a gas-tight glass syringe and immediately injected into a gas chromatograph to assess 14-15N2 and 15-15N2 concentrations.
Up-flow column reactors fed with NH2OH
Three 255-mL up-flow column reactors were operated at 37°C in the dark with the continuous feeding of the above culture media containing 1) 0.7 mM 14NH2OH, 2) 0.7 mM 14NH2OH and 0.5 mM 14NH4+, or 3) 0.7 mM 14NH2OH and 0.5 mM 14NO2–. B. sinica cells immobilized on polyvinyl alcohol (PVA)-sodium alginate (SA) (6 and 2% [w/v], respectively) beads were inoculated into the column reactors at a packing ratio of 50% (v/v). The gel immobilization of B. sinica cells in PVA-SA gel beads was performed as previously described (Ali et al., 2015). Briefly, the planktonic cells of B. sinica collected from the above MBR were resuspended in culture media, and mixed with an equal volume of PVA-alginate (FUJIFILM Wako) solution (12 and 4% [w/v], respectively). The gel solution was dropped using a disposable 50-mL plastic syringe (Terumo) into a 4% (w/v) CaCl2 solution to form gel beads (diameter of ca. 2 mm). After an overnight incubation at 20°C, gel beads were washed with fresh inorganic medium. The gel beads obtained were inoculated into the up-flow column reactors.
Chemical analysis
NH4+, NO2–, and NO3– concentrations were measured using the ion chromatograph IC-2010 equipped with the TSKgel SuperIC-Anion HS or TSKgel SuperIC-Cation HS column (Tosoh). NH2OH concentrations were measured colorimetrically (Frear and Burrell, 1955). Briefly, liquid samples were mixed with 0.48% (w/v) trichloroacetic acid, 0.2% (w/v) 8-hydroxyquinoline, and 0.2 M Na2CO3, heated at 100°C for 1 min, and absorbance was then measured at a wavelength of 705 nm using the spectrophotometer V-630bio (Jasco). N2H4 concentrations were measured colorimetrically using p-dimethyl-aminobenzaldehyde (Watt and Chrisp, 1952). Briefly, liquid samples were mixed with 0.12 M p-dimethyl-aminobenzaldehyde, and absorbance was measured at a wavelength of 460 nm.
14-15N2 and 15-15N2 concentrations were measured by gas chromatography mass spectrometry (GC/MS) (Isobe et al., 2011a; 2011b). Fifty microliters of the headspace gas was collected using a 100-μL gas-tight glass syringe and immediately injected into the gas chromatograph GCMS-QP 2010 SE (Shimadzu) equipped with a fused silica capillary column (Agilent Technologies). Peaks at m/z=29 and 30 corresponding to 14-15N2 and 15-15N2 were monitored, and concentrations were calculated using a standard curve prepared using 15-15N2 gas (Cambridge Isotope Laboratories). The 14-15N2 and 15-15N2 concentrations of 15NH2OH were calculated by considering the 15N enrichment of 15NH2OH·HCl (15N, 98%) and the natural abundance of 14-15N2 and 15-15N2 in atmospheric N2 gas contaminated at the injection of the gas sample.
qPCR assay
The copy numbers of the anammox bacterial 16S rRNA gene were measured using a qPCR assay. Genomic DNA was extracted from gel beads collected from the up-flow column reactors using the FastDNA SPIN kit (Qiagen) according to the instruction manual supplied by the manufacturer. The qPCR assay was conducted using the ABI7500 fast Real-Time PCR System (Thermo Fisher Scientific) and Premix Ex Taq (Probe qPCR) (TakaraBio) under previously described thermal conditions (Zhang and Okabe, 2017a). The oligonucleotide primers and TaqMan probe used for the PCR amplification of the B. sinica 16S rRNA gene were BRS95F, BRS170R, and BRS130P. Standard curves (101 to 106 copies μL–1) were prepared using a dilution series of plasmid DNAs containing a partial B. sinica 16S rRNA gene sequence.
Results
NH2OH disproportionation by phylogenetically different anammox bacteria
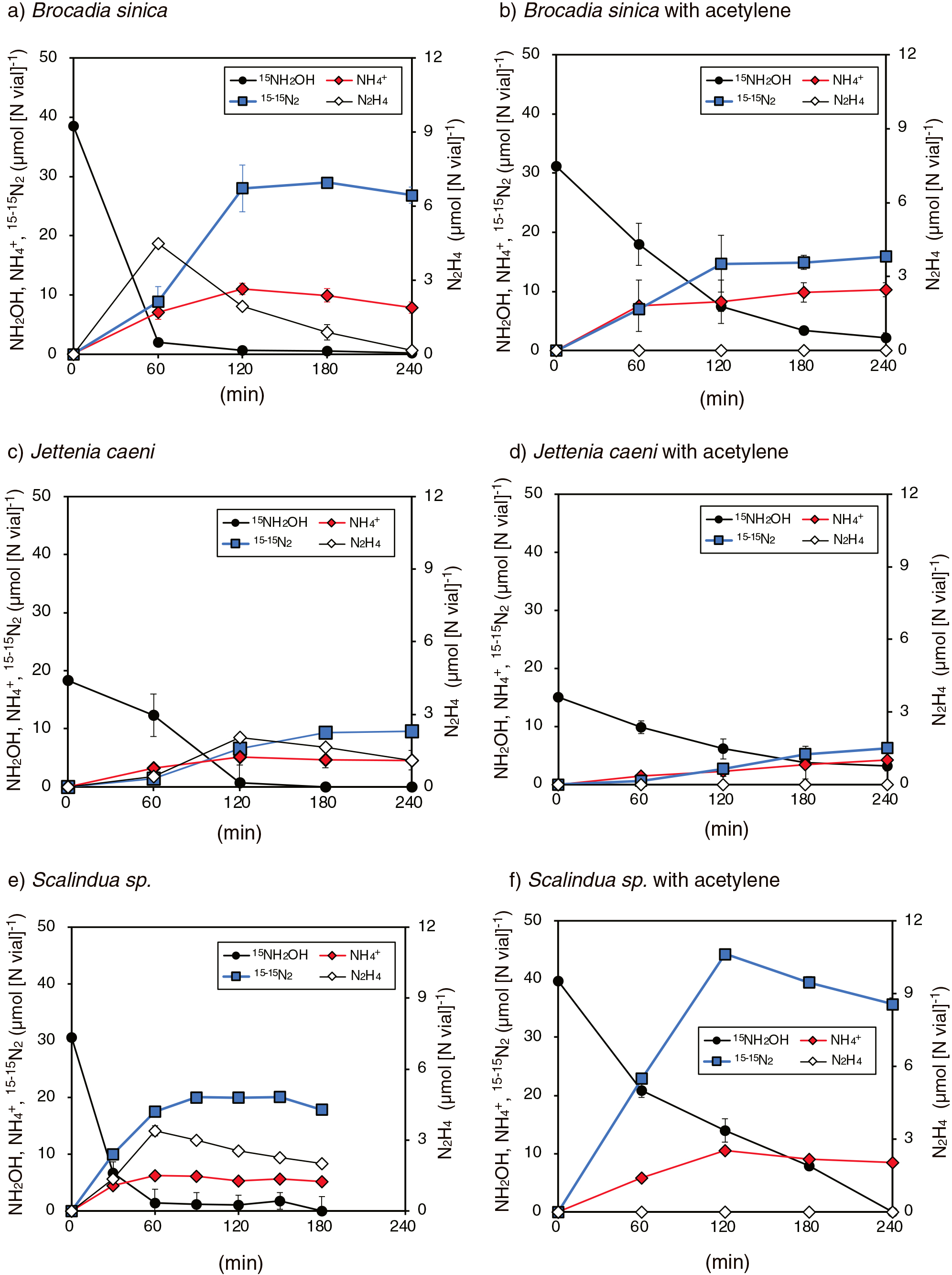
Anammox bacterial cells of B. sinica, J. caeni, and Scalindua sp. were incubated with the addition of 1.0 to 2.5 mM 15NH2OH. As shown in Fig. 1a, c, and e, 15NH2OH consumption occurred in the culture concurrently with the production of 15-15N2 gas and NH4+. N2H4 was also produced and markedly increased when NH2OH concentrations decreased below 0.1 mM (i.e., a 60-min incubation for B. sinica and Scalindua sp. and a 120-min incubation for J. caeni). 14-15N2, NO2–, and NO3– were not detected during any incubations, and 15NH2OH consumption was negligible in vials without anammox bacterial cells (i.e., abiotic control incubation).
Batch incubations were repeated with the addition of 30 μM acetylene. Anammox bacterial cells consumed 15NH2OH (Fig. 1b, d, and f), whereas consumption rates were >3-fold lower than those without acetylene. Although 15-15N2 gas and NH4+ were produced during the incubation, N2H4 was not produced in any anammox bacterial cultures.
The above batch incubation of B. sinica was repeated with an increase in the initial NH2OH concentration (from 5 to 10 mM) with/without 30 μM acetylene. As shown in Fig. 2, 15NH2OH consumption and the concomitant production of 15-15N2 and NH4+ occurred, similar to the batch incubation with the addition of 2.5 mM 15NH2OH (i.e., Fig. 1a and b). It is important to note that N2H4 production only occurred when NH2OH concentrations decreased below 1.5 mM (Fig. 2a after 7.5 h of the incubation), and N2H4 production was not observed in Fig. 2c and e.
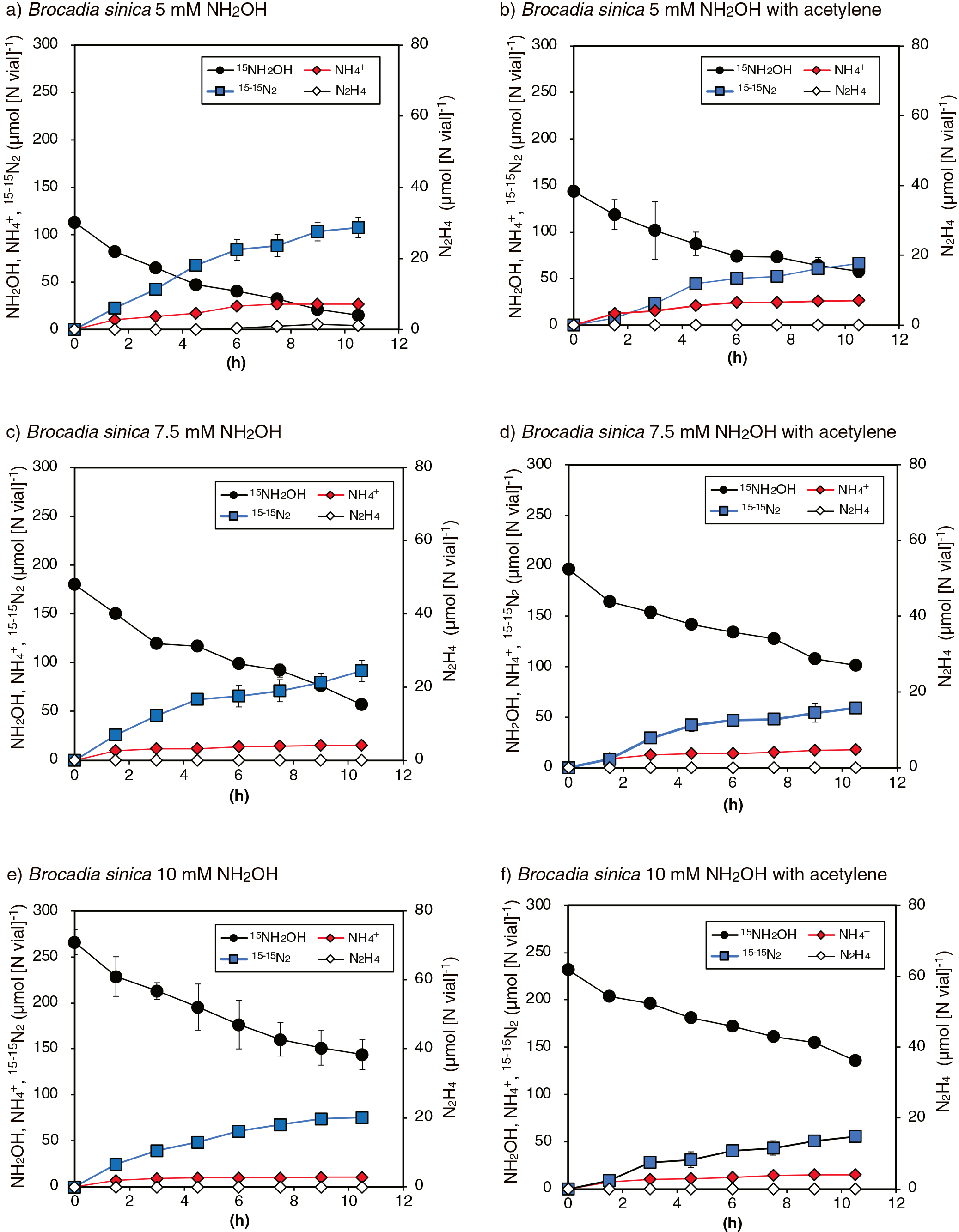
The stoichiometry and nitrogen mass balance of the above batch incubations are shown in Table 1. The theoretical values for Δ15-15N2/Δ15NH2OH and Δ15NH4+/Δ15NH2OH were 0.33 and 0.33, respectively when NH2OH disproportionation occurred according to eq. 2. The values for Δ15-15N2/Δ15NH2OH and Δ15NH4+/Δ15NH2OH obtained from batch incubations with the addition of 2.5 or 5 mM 15NH2OH were in the range of 0.19–0.45 and 0.17–0.36, respectively. These values were generally close to the theoretical values and those for Δ15NH4+/Δ15NH2OH in the K. stuttgartiensis culture (0.19–0.48), except for the following batch incubations: B. sinica with 2.5 mM 15NH2OH without acetylene (0.2 for Δ15NH4+/Δ15NH2OH), B. sinica with 5 mM 15NH2OH with acetylene (0.19 for Δ15-15N2/Δ15NH2OH), and Scalindua sp. with 2.5 mM 15NH2OH with and without acetylene (0.17 or 0.21 for Δ15NH4+/Δ15NH2OH). Batch incubations of B. sinica with 7.5 or 10 mM NH2OH generally resulted in lower values for Δ15-15N2/Δ15NH2OH and Δ15NH4+/Δ15NH2OH, which were in the ranges of 0.14–0.19 and 0.09–0.19, respectively.
Table 1.
Stoichiometry of NH
2OH disproportionation mediated by anammox bacteria. The values obtained in the present study were mean values from triplicate biological replicates, and values were calculated from the initial and final concentrations during batch incubations. The N-mass balance (%) was calculated by dividing the total amounts of
15NH
2OH,
15NH
4+, and
15-15N
2 at the end of the incubation (μmol [N vial]
–1) by the initial amounts of
15NH
2OH (μmol [N vial]
–1).
| Species |
NH2OH (mM) |
Acetylene* |
N-mass balance |
Δ15-15N2/ΔNH2OH (mol/mol) |
Δ15NH4+/ΔNH2OH (mol/mol) |
References |
| B. sinica |
2.5 |
w/o |
91% |
0.35 |
0.20 |
This study |
|
5 |
w/o |
85% |
0.28 |
0.27 |
|
|
7.5 |
w/o |
66% |
0.19 |
0.12 |
|
|
10 |
w/o |
72% |
0.16 |
0.09 |
|
|
2 |
+ |
91% |
0.27 |
0.35 |
|
|
5 |
+ |
81% |
0.19 |
0.30 |
|
|
7.5 |
+ |
76% |
0.16 |
0.19 |
|
|
10 |
+ |
77% |
0.14 |
0.15 |
|
| J. caeni |
1 |
w/o |
83% |
0.26 |
0.25 |
|
|
|
+ |
91% |
0.26 |
0.36 |
|
| Scalindua sp. |
2 |
w/o |
82% |
0.29 |
0.17 |
|
|
2.5 |
+ |
111% |
0.45 |
0.21 |
|
| K. stuttgartiensis |
1.6–10 |
w/o |
NA |
ND |
0.19–0.48 |
van der Star et al., 2008
Soler-Jofra et al., 2020 |
| B. flugida |
4 |
w/o |
NA |
ND |
0.25 |
van der Star et al., 2008 |
*; w/o and +: without and with the addition of 30 μM acetylene, respectively, NA: not available, ND: not determined.
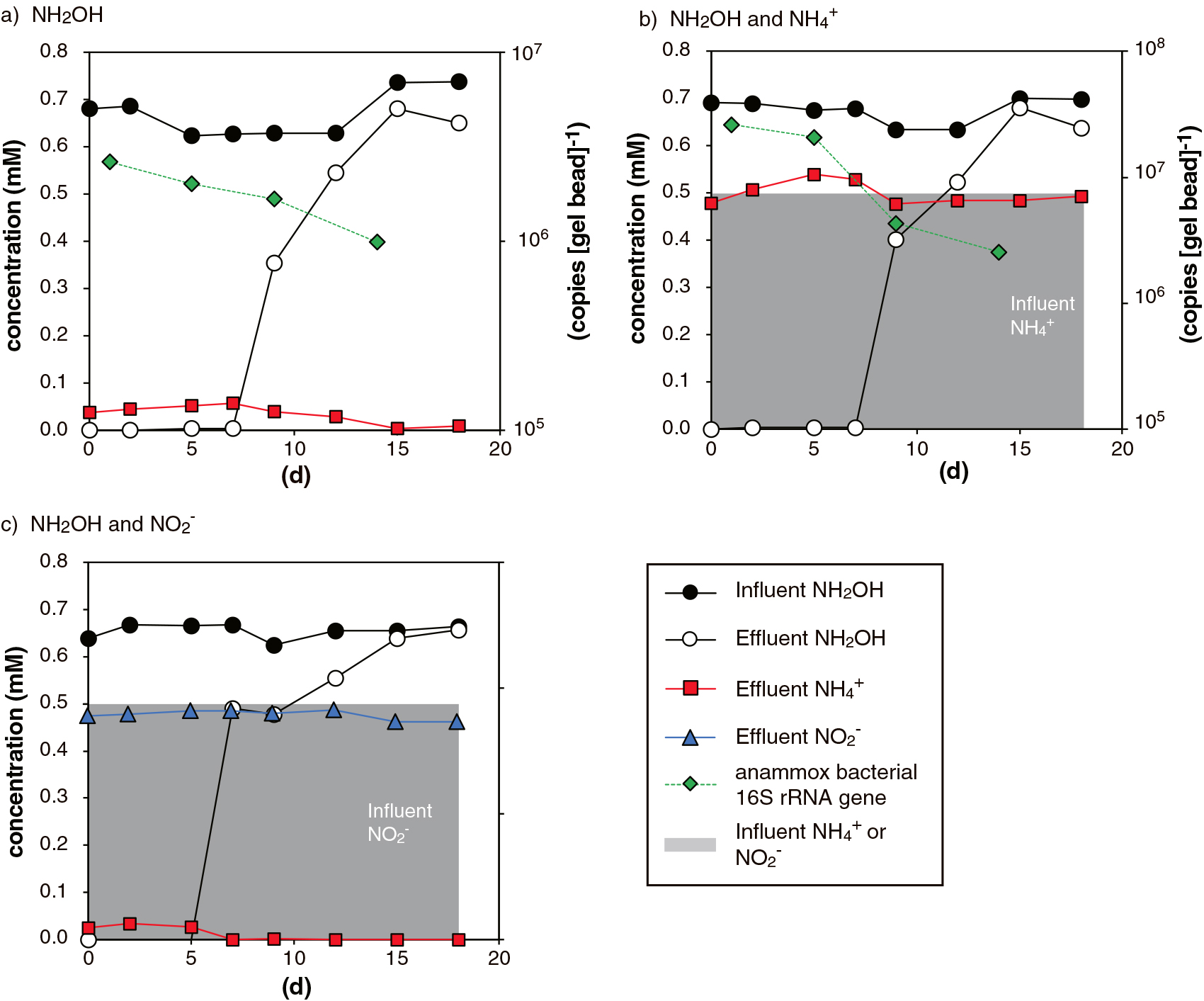
NH2OH consumption and the abundance of the anammox bacterial 16S rRNA gene were examined in the 1) NH2OH-, 2) NH2OH and NH4+-, and 3) NH2OH and NO2–-feeding up-flow column reactors. In all operated reactors, NH2OH consumption markedly decreased after 7 d of operation, and halted after 15 d of operation (Fig. 3). NH4+ concentrations in the effluents of the NH2OH- and NH2OH and NO2–-feeding reactors were in the same range (Fig. 3a and c, respectively), and the consumption of NH4+ did not occur in the NH2OH and NH4+-feeding reactor (Fig. 3b). The copy numbers of the anammox bacterial 16S rRNA gene decreased over time, and its abundance after 14 d of operation was an order of magnitude less than that after 1 d of operation. Copy numbers were not measured in the reactor fed with NH2OH and NO2– (Fig. 3c).
Discussion
NH2OH disproportionation was examined using phylogenetically different anammox bacteria, and the present study clearly indicated that NH2OH disproportionation is a common nitrogen transformation process of anammox bacteria (Table 1). Although previous studies (van der Star et al., 2008; Soler-Jofra et al., 2020) reported anammox bacterial NH2OH disproportionation, they did not examine the amount of N2 gas produced. The present study performed sophisticated 15NH2OH-tracing batch incubations, and revealed that anammox bacteria yielded both N2 gas and NH4+ from NH2OH disproportionation. 15NH2OH-tracing batch incubations enabled the stoichiometry of 15-15N2 and 15NH4+ production to 15NH2OH consumption to be examined (Table 1), and the results obtained indicated that an increase in the initial concentration of NH2OH resulted in a decrease in Δ15-15N2/Δ15NH2OH and Δ15NH4+/Δ15NH2OH for B. sinica. Furthermore, the addition of acetylene resulted in a decrease and increase in Δ15-15N2/Δ15NH2OH and Δ15NH4/Δ15NH2OH, respectively. These variations in stoichiometry suggested that 15N2 and/or 15NH4+ were produced by multiple nitrogen transformation pathways other than NH2OH disproportionation. Anammox bacterial hydroxylamine dehydrogenase oxidizes NH2OH to NO (Maalcke et al., 2014), and N2 gas may be produced using the NO formed by the anammox process (the coupling of NH4+ and NO) (Kartal et al., 2013). Although the reduction of NH2OH to NH4+ by anammox bacterial cells has not yet been demonstrated, anammox bacteria are capable of dissimilatory nitrite reduction to ammonium (Kartal et al., 2007).
N2 and NH4+-forming NH2OH disproportionation has not yet been reported for microorganisms other than anammox bacteria. It requires the formation of a nitrogen-nitrogen bond for N2 gas, and only nitric oxide reductase and hydrazine synthase catalyze this reaction (Dietl et al., 2015). This may be the reason why N2 and NH4+-forming NH2OH disproportionation has not yet been detected in microbial cultures other than anammox bacteria. Apart from N2 and NH4+-forming NH2OH disproportionation, 1) NH4+ and NO-, 2) NH4+ and NO2–-, and 3) N2O and NH4+-forming NH2OH disproportionation are thermodynamically favorable (Pacheco et al., 2011); however, limited information is currently available on their activities or involvement in the nitrogen transformation process in natural and man-made ecosystems.
The addition of acetylene did not induce the accumulation of N2H4, but reduced NH2OH consumption (Fig. 1b, d, and f), which indicated that acetylene inhibited the enzymes involved in both NH2OH consumption and N2H4 production reactions. B. sinica hydrazine synthase utilizes NH2OH as a substrate for N2H4 synthesis (Oshiki et al., 2016a); therefore, hydrazine synthase may be the enzyme that is inhibited by acetylene. The inhibition of anammox bacterial activity by acetylene has been reported (Jensen et al., 2007); however, the underlying mechanism(s) remain unknown. Apart from hydrazine synthase, acetylene inhibited the NO2– reduction reaction of anammox bacteria (Kartal et al., 2011; Oshiki et al., 2016a), indicating that acetylene suppresses multiple nitrogen transformation reactions of anammox bacteria. Although the acetylene inhibition of copper-containing metalloproteins, such as ammonia monooxygenase and nitrous oxide reductase (Amo and Nos, respectively) has been described (Ensign et al., 1993; Gilch et al., 2009), the involvement of copper-containing metalloproteins in anammox bacterial metabolism, particularly N2H4 production, remains unclear. The binding site of acetylene to anammox bacterial enzymes is of interest for obtaining a more detailed understanding of the acetylene inhibition of anammox bacteria. The addition of 30 μM acetylene did not completely inhibit the production of 15-15N2 gas or NH4+ (Fig. 1 and 2). This result suggests the incomplete inhibition of anammox bacterial hydrazine synthase with the addition of 30 μM acetylene (Jensen et al., 2007) and/or the production of 15-15N2 gas and NH4+, but not through N2H4. The corresponding mechanism has not yet been elucidated, and a further understanding of acetylene inhibition by anammox bacteria will provide novel insights.
Apart from the above acetylene inhibition, high NH2OH concentrations (>0.1 mM NH2OH in Fig 1a, c, and e, and >1.5 mM NH2OH in Fig. 2a) did not induce the accumulation of N2H4. This behavior cannot simply be explained by the inhibition of anammox bacterial hydrazine dehydrogenase involved in N2H4 oxidation to N2. The N2H4 oxidation activities of purified anammox bacterial hydrazine dehydrogenase were inhibited in the presence of 2.4 to 7.9 μM NH2OH (Shimamura et al., 2007; Maalcke et al., 2016), and this inhibition provides an explanation for the accumulation of N2H4 during batch incubations, but not for the lack of its accumulation at high NH2OH concentrations. On the other hand, anammox bacterial N2H4 accumulation may be explained by the balance between N2H4 production and consumption reactions (eq. 3 and 4, respectively), as reported in previous studies (van der Star et al., 2008; Soler-Jofra et al., 2020). N2H4 production and consumption reactions require 1 and 2 moles of NH2OH, respectively, and higher NH2OH concentrations increase N2H4 consumption rates more than N2H4 production rates; therefore, N2H4 does not accumulate at high NH2OH concentrations. The affinity constants of eq. 3 and 4 for NH2OH need to be examined in more detail in order to clarify the N2H4 accumulation behavior of anammox bacterial cells.
The results of the up-flow column reactor experiments revealed that B. sinica did not proliferate with NH2OH disproportion (Fig. 3), and this is the first experimental evidence to show that an anammox culture cannot be maintained with NH2OH as the sole energy source. A 1 log reduction in the B. sinica 16S rRNA gene copy number clearly indicated that B. sinica did not proliferate in the operated reactors. B. sinica cells may have been compromised during reactor operation due to the high toxicity and mutagenesis of NH2OH, which resulted in a decrease in the B. sinica 16S rRNA gene copy number in PVA-SA gel beads. We previously reported a log reduction in the anammox bacterial 16S rRNA gene copy number in PVA-SA gel beads under unfavorable cultivation conditions (Zhang et al., 2017b). It is important to note that B. sinica cells preferentially performed NH2OH disproportionation over anammox. Anammox using NO2– or NH2OH did not occur in the NH2OH and NO2–- or NH2OH and NH4+-feeding reactors (Fig. 3c and b, respectively), which indicated that the anammox activities of B. sinica were inhibited in these reactors by NH2OH. Therefore, low NH2OH concentrations need to be maintained in order to avoid the inhibition of anammox activities and achieve the stable performance of nitrogen removal. Although the inhibitory effects of NH2OH on nitrifying bacteria have been investigated (Kindaichi et al., 2004; Soler-Jofra et al., 2021), the IC50 concentration of NH2OH for anammox bacteria has not yet been systematically examined. In the present study, the sudden deterioration of NH2OH consumption occurred after 7 d of operation; however, the concentration of NH2OH in the influent was markedly lower than those in batch incubations in which the marked deterioration of NH2OH consumption did not occur (i.e., 0.7 and 2.5–10 mM, respectively). Therefore, it was not possible to approximate the IC50 concentration of NH2OH inhibition from short-term batch incubations, and further studies are warranted to assess the IC50 concentration of the long-term inhibition of NH2OH. NH2OH may accumulate at the submicromolar range in natural aquatic environments (Fukumori et al., 2003; Bikbulatova et al., 2007) and at the submillimolar range in nitrifying cultures (Soler-Jofra et al., 2021 and references therein); however, the impact of NH2OH on nitrogen transformation reactions remains unclear. Further studies are needed to examine NH2OH transformation, including anammox bacterial NH2OH disproportionation, in natural and man-made ecosystems.
Citation
Oshiki, M., Gao, L., Zhang, L., and Okabe, S. (2022) NH2OH Disproportionation Mediated by Anaerobic Ammonium-oxidizing (Anammox) Bacteria. Microbes Environ 37: ME21092.
https://doi.org/10.1264/jsme2.ME21092
Acknowledgements
This work was supported by JSPS KAKENHI Grant numbers 17K15305 and 19K05805 to M.O. and 19H00776 to S.O., respectively. The authors declare no conflicts of interest associated with this manuscript.
References
- Ali, M., Oshiki, M., Rathnayake, L., Ishii, S., Satoh, H., and Okabe, S. (2015) Rapid and successful start-up of anammox process by immobilizing the minimal quantity of biomass in PVA-SA gel beads. Water Res
79: 147–157.
- Amano, T., Yoshinaga, I., Okada, K., Yamagishi, T., Ueda, S., Obuchi, A., et al. (2007) Detection of anammox activity and diversity of anammox bacteria-related 16S rRNA genes in coastal marine sediment in Japan. Microbes Environ
22: 232–242.
- Bikbulatova, E.M., Stepanova, I.E., and Bikbulatov, E.S. (2007) Concentration and localization of hydroxylamine in the reservoirs and lakes in the territory of European Russia. Water Resour
34: 554–562.
- Daims, H., Brühl, A., Amann, R., Schleifer, K.H., and Wagner, M. (1999) The domain-specific probe EUB338 is insufficient for the detection of all bacteria. Development and evaluation of a more comprehensive probe set. Syst Appl Microbiol
22: 434–444.
- Dietl, A., Ferousi, C., Maalcke, W.J., Menzel, A., Vries, S. de, Keltjens, J.T., et al. (2015) The inner workings of the hydrazine synthase multiprotein complex. Nature
527: 394–397.
- Egli, K., Bosshard, F., Werlen, C., Lais, P., Siegrist, H., Zehnder, A.J.B., and van der Meer, J.R. (2003) Microbial composition and structure of a rotating biological contactor biofilm treating ammonium-rich wastewater without organic carbon. Microb Ecol
45: 419–432.
- Ensign, S.A., Hyman, M.R., and Arp, D.J. (1993) In vitro activation of ammonia monooxygenase from Nitrosomonas europaea by copper. J Bacteriol
175: 1971–1980.
- Ferousi, C., Schmitz, R.A., Maalcke, W.J., Lindhoud, S., Versantvoort, W., Jetten, M.S.M., et al. (2021) Characterization of a nitrite-reducing octaheme hydroxylamine oxidoreductase that lacks the tyrosine cross-link. J Biol Chem
296: 100476.
- Finster, K. (2008) Microbiological disproportionation of inorganic sulfur compounds. J Sulfur Chem
29: 281–292.
- Frear, D.S., and Burrell, R.C. (1955) Spectrophotometric method for determining hydroxylamine reductase activity in higher plants. Anal Chem
27: 1664–1665.
- Fukumori, R., Senga, Y., Okumura, M., Fujinaga, K., and Seike, Y. (2003) Pretreatment and pre concentration methods for hydroxyl amine in environmental water samples using solid-phase extraction, followed by a spectrophotometric determination. Bunseki Kagaku
52: 747–753 (in Japanese with an English abstract).
- Gilch, S., Vogel, M., Lorenz, M.W., Meyer, O., and Schmidt, I. (2009) Interaction of the mechanism-based inactivator acetylene with ammonia monooxygenase of Nitrosomonas europaea. Microbiology
155: 279–284.
- Isobe, K., Koba, K., Ueda, S., Senoo, K., Harayama, S., and Suwa, Y. (2011a) A simple and rapid GC/MS method for the simultaneous determination of gaseous metabolites. J Microbiol Methods
84: 46–51.
- Isobe, K., Suwa, Y., Ikutani, J., Kuroiwa, M., Makita, T., Takebayashi, Y., et al. (2011b) Analytical techniques for quantifying 15N/14N of nitrate, nitrite, total dissolved nitrogen and ammonium in environmental samples using a gas chromatograph equipped with a quadrupole mass spectrometer. Microbes Environ
26: 46–53.
- Jensen, M.M., Thamdrup, B., and Dalsgaard, T. (2007) Effects of specific inhibitors on anammox and denitrification in marine sediments. Appl Environ Microbiol
73: 3151–3158.
- Kartal, B., Kuypers, M.M.M., Lavik, G., Schalk, J., Op den Camp, H.J.M., Jetten, M.S.M., and Strous, M. (2007) Anammox bacteria disguised as denitrifiers: nitrate reduction to dinitrogen gas via nitrite and ammonium. Environ Microbiol
9: 635–642.
- Kartal, B., Maalcke, W.J., de Almeida, N.M., Cirpus, I., Gloerich, J., Geerts, W., et al. (2011) Molecular mechanism of anaerobic ammonium oxidation. Nature
479: 127–130.
- Kartal, B., de Almeida, N.M., Maalcke, W.J., Op den Camp, H.J.M., Jetten, M.S.M., and Keltjens, J.T. (2013) How to make a living from anaerobic ammonium oxidation. FEMS Microbiol Rev
37: 428–461.
- Kindaichi, T., Okabe, S., Satoh, H., and Watanabe, Y. (2004) Effects of hydroxylamine on microbial community structure and function of autotrophic nitrifying biofilms determined by in situ hybridization and the use of microelectrodes. Water Sci Technol
49: 61–68.
- Kindaichi, T., Awata, T., Suzuki, Y., Tanabe, K., Hatamoto, M., Ozaki, N., and Ohashi, A. (2011) Enrichment using an up-flow column reactor and community structure of marine anammox bacteria from coastal sediment. Microbes Environ
26: 67–73.
- Kuypers, M.M.M., Sliekers, A.O., Lavik, G., Schmid, M., Jørgensen, B.B., Kuenen, J.G., et al. (2003) Anaerobic ammonium oxidation by anammox bacteria in the Black Sea. Nature
422: 608–611.
- Kuypers, M.M.M., Lavik, G., Woebken, D., Schmid, M., Fuchs, B.M., Amann, R., et al. (2005) Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation. Proc Natl Acad Sci U S A
102: 6478–6483.
- Lotti, T., Kleerebezem, R., Lubello, C., and van Loosdrecht, M.C.M. (2014) Physiological and kinetic characterization of a suspended cell anammox culture. Water Res
60: 1–14.
- Maalcke, W.J., Dietl, A., Marritt, S.J., Butt, J.N., Jetten, M.S.M., Keltjens, J.T., et al. (2014) Structural basis of biological NO generation by octaheme oxidoreductases. J Biol Chem
289: 1228–1242.
- Maalcke, W.J., Reimann, J., de Vries, S., Butt, J.N., Dietl, A., Kip, N., et al. (2016) Characterization of anammox hydrazine dehydrogenase, a key N2-producing enzyme in the global nitrogen cycle. J Biol Chem
291: 17077–17092.
- Madigan, M.T., Bender, K.S., Buckley, D.H., Sattley, W.M., and Stahl, D.A. (2019) Brock Biology of Microorganisms, 15th edn. London: Pearson Education.
- Mulder, A., van de Graaf, A.A., Robertson, L.A., and Kuenen, J.G. (1995) Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor. FEMS Microbiol Ecol
16: 177–183.
- Oshiki, M., Shimokawa, M., Fujii, N., Satoh, H., and Okabe, S. (2011) Physiological characteristics of the anaerobic ammonium-oxidizing bacterium “Candidatus Brocadia sinica.”
Microbiology
157: 1706–1713.
- Oshiki, M., Awata, T., Kindaichi, T., Satoh, H., and Okabe, S. (2013) Cultivation of planktonic anaerobic ammonium oxidation (anammox) bacteria by using membrane bioreactor. Microbes Environ
28: 436–443.
- Oshiki, M., Ali, M., Shinyako-Hata, K., Satoh, H., and Okabe, S. (2016a) Hydroxylamine-dependent anaerobic ammonium oxidation (anammox) by “Candidatus Brocadia sinica.”
Environ Microbiol
18: 3133–3143.
- Oshiki, M., Satoh, H., and Okabe, S. (2016b) Ecology and physiology of anaerobic ammonium oxidizing (anammox) bacteria. Environ Microbiol
18: 2784–2796.
- Pacheco, A.A., McGarry, J., Kostera, J., and Corona, A. (2011) Techniques for investigating hydroxylamine disproportionation by hydroxylamine oxidoreductases. Methods Enzymol
486: 447–463.
- Quan, Z.X., Rhee, S.K., Zuo, J.E., Yang, Y., Bae, J.W., Park, J.R., et al. (2008) Diversity of ammonium-oxidizing bacteria in a granular sludge anaerobic ammonium-oxidizing (anammox) reactor. Environ Microbiol
10: 3130–3139.
- Schmid, M., Schmitz-Esser, S., Jetten, M., and Wagner, M. (2001) 16S-23S rDNA intergenic spacer and 23S rDNA of anaerobic ammonium-oxidizing bacteria: Implications for phylogeny and in situ detection. Environ Microbiol
3: 450–459.
- Shimamura, M., Nishiyama, T., Shigetomo, H., Toyomoto, T., Kawahara, Y., Furukawa, K., and Fujii, T. (2007) Isolation of a multiheme protein with features of a hydrazine-oxidizing enzyme from an anaerobic ammonium-oxidizing enrichment culture. Appl Environ Microbiol
73: 1065–1072.
- Soler-Jofra, A., Laurenia, M., Warmerdama, M., Pérezb, J., and van Loosdrecht, M.C.M. (2020) Hydroxylamine metabolism of Ca. Kuenenia stuttgartiensis. Water Res
184: 116188.
- Soler-Jofra, A., Pérez, J., and van Loosdrecht, M.C.M. (2021) Hydroxylamine and the nitrogen cycle: A review. Water Res
190: 116723.
- Strous, M., Fuerst, J., Kramer, E., Logemann, S., Muyzer, G., van de Pas-Schoonen, K., et al. (1999) Missing lithotroph identified as new planctomycete. Nature
400: 446–449.
- Strous, M., Pelletier, E., Mangenot, S., Rattei, T., Lehner, A., Taylor, M., et al. (2006) Deciphering the evolution and metabolism of an anammox bacterium from a community genome. Nature
440: 790–794.
- van de Graaf, A.A., de Bruijn, P., Robertson, L.A., Jetten, M.S.M., and Kuenen, J.G. (1996) Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor. Microbiology
142: 2187–2196.
- van de Graaf, A.A., de Bruijn, P., Robertson, L.A., Jetten, M.S.M., and Kuenen, J.G. (1997) Metabolic pathway of anaerobic ammonium oxidation on the basis of 15N studies in a fluidized bed reactor. Microbiology
143: 2415–2421.
- van der Star, W.R.L., van de Graaf, M.J., Kartal, B., Picioreanu, C., Jetten, M.S.M., and van Loosdrecht, M.C.M. (2008) Response of anaerobic ammonium-oxidizing bacteria to hydroxylamine. Appl Environ Microbiol
74: 4417–4426.
- Watt, G.W., and Chrisp, J.D. (1952) A spectrophotometric method for the determination of hydrazine. Anal Chem
24: 2006–2008.
- Yoshinaga, I., Amano, T., Yamagishi, T., Okada, K., Ueda, S., Sako, Y., and Suwa, Y. (2011) Distribution and diversity of anaerobic ammonium oxidation (anammox) bacteria in the sediment of a eutrophic freshwater lake, lake Kitaura, Japan. Microbes Environ
26: 189–197.
- Zhang, L., and Okabe, S. (2017a) Rapid cultivation of free-living planktonic anammox cells. Water Res
127: 204–210.
- Zhang, L., Narita, Y., Gao, L., Ali, M., Oshiki, M., and Okabe, S. (2017b) Maximum specific growth rate of anammox bacteria revisited. Water Res
116: 296–303.
- Zhang, L., and Okabe, S. (2020) Ecological niche differentiation among anammox bacteria. Water Res
171: 115468.