Abstract
Many stinkbugs in the superfamily Coreoidea (Hemiptera: Heteroptera) develop crypts in the posterior midgut, harboring Caballeronia (Burkholderia) symbionts. These symbionts form a monophyletic group in Burkholderia sensu lato, called the “stinkbug-associated beneficial and environmental (SBE)” group, recently reclassified as the new genus Caballeronia. SBE symbionts are separated into the subclades SBE-α and SBE-β. Previous studies suggested a regional effect on the symbiont infection pattern; Japanese and American bug species are more likely to be associated with SBE-α, while European bug species are almost exclusively associated with SBE-β. However, since only a few insect species have been investigated, it remains unclear whether region-specific infection is general. We herein investigated Caballeronia gut symbionts in diverse Japanese, European, and North American populations of a cosmopolitan species, the Western conifer seed bug Leptoglossus occidentalis (Coreoidea: Coreidae). A molecular phylogenetic analysis of the 16S rRNA gene demonstrated that SBE-β was the most dominant in all populations. Notably, SBE-α was rarely detected in any region, while a third clade, the “Coreoidea clade” occupied one fourth of the tested populations. Although aposymbiotic bugs showed high mortality, SBE-α- and SBE-β-inoculated insects both showed high survival rates; however, a competition assay demonstrated that SBE-β outcompeted SBE-α in the midgut crypts of L. occidentalis. These results strongly suggest that symbiont specificity in the Leptoglossus-Caballeronia symbiotic association is influenced by the host rather than geography, while the geographic distribution of symbionts may be more important in other bugs.
Recent studies revealed that insect gut microorganisms play a pivotal role in the evolution and environmental adaptation of insects. Gut microorganisms provide essential nutrients, digest indigestible food materials, and/or degrade phytotoxins and insecticides (Kikuchi et al., 2012; Engel and Moran, 2013; Salem et al., 2014; Sudakaran et al., 2017; Itoh et al., 2018; Moran et al., 2019). Many species of stinkbugs in the superfamily Coreoidea (Hemiptera: Heteroptera) develop numerous crypts at the posterior part of the midgut, wherein specific Caballeronia symbionts (previously included in the genus Burkholderia) densely proliferate - generally as a single species - until almost full occupation of the luminal space (Kikuchi et al., 2005, 2011a; Olivier-Espejel et al., 2011; Garcia et al., 2014; Kuechler et al., 2016; Takeshita and Kikuchi, 2017; Ohbayashi et al., 2019b; Acevedo et al., 2021; Hunter et al., 2022). Caballeronia gut symbionts play important roles in their hosts, such as the recycling of metabolic waste materials and providing essential amino acids and vitamins, thereby enhancing the growth and fecundity of stinkbugs (Kikuchi et al., 2007; Kikuchi and Fukatsu, 2014; Ohbayashi et al., 2019a). A particularity of this mono-species symbiosis is the horizontal transmission of symbionts. Hatchlings are symbiont-free and acquire Caballeronia symbionts from soil during the early instar stages (Kikuchi et al., 2007, 2011b; Ohbayashi et al., 2019b). This implies not only that insects depend on efficient and stringent selection mechanisms to sort environmental bacteria in order to give access to Caballeronia symbionts only (Ohbayashi et al., 2015; Itoh et al., 2019; Kikuchi et al., 2020), but also that the geographic distribution of symbiont species may be a factor influencing the outcome of symbiosis (Ohbayashi et al., 2019b).
The genus Burkholderia was initially separated from Pseudomonas Group II in 1992 (Yabuuchi et al., 1992) into a heterogeneous taxonomic group of more than 100 bacterial species (Eberl and Vandamme, 2016). In a recent reclassification of this Burkholderia “sensu lato” taxonomic group, six new genera (Paraburkholderia, Caballeronia, Robbsia, Mycetohabitans, Pararobbsia, and Trinickia) have been proposed next to the genus Burkholderia sensu strico (Sawana et al., 2014; Dobritsa and Samadpour, 2016; Beukes et al., 2017; Lopes-Santos et al., 2017; Estrada-de Los Santos et al., 2018; Lin et al., 2020). Caballeronia is also called the Stinkbug-associated Beneficial and Environmental (SBE) group of Burkholderia, which is divided into two subgroups, group-α (SBE-α) and group-β (SBE-β).
A previous survey of Caballeronia symbionts in stinkbugs revealed region-specific infection with species of the two subgroups: SBE-α was more likely to be detected in Japanese and American stinkbug species of Coreoidea (Kikuchi et al., 2005, 2011b; Olivier-Espejel et al., 2011; Garcia et al., 2014; Kuechler et al., 2016; Ohbayashi et al., 2019b; Acevedo et al., 2021; Hunter et al., 2022), and SBE-β in European species of Coreoidea (Kuechler et al., 2016; Ohbayashi et al., 2019b). However, since only a few insect species have been investigated, and even fewer species from a wide geographic distribution, it remains unclear whether region-specific infection is general. The mechanisms underlying region-specific infection have not yet been elucidated, but may be influenced by the region-dependent composition of the soil microbiota.
The Western conifer seed bug Leptoglossus occidentalis (Coreoidea: Coreidae) (Fig. 1A), a notorious pest of conifer forests (Lesieur et al., 2014), originates from North America (Heidemann, 1910; Koerber, 1963). This stinkbug has become a serious invading pest worldwide. In 1999, L. occidentalis was found in Europe for the first time in Italy (Taylor et al., 2001) and its population has been rapidly expanding in recent years throughout Europe (Dusoulier et al., 2007; Malumphy et al., 2008; Fent and Kment, 2011; Gapon, 2013; Lesieur et al., 2018; van der Heyden, 2019) and other more distant regions, such as North Africa (Ben Jamaa et al., 2013; Gapon, 2015). It has also spread to South America (Faúndez et al., 2017) and Asia (Ishikawa and Kikuhara, 2009; Ahn et al., 2013). In Japan, L. occidentalis was initially collected in Tokyo in 2008 (Ishikawa and Kikuhara, 2009) and, similar to Europe, it has since rapidly spread to almost all areas of Japan, including the Tohoku and Kyushu districts (Tsuru et al., 2020). Therefore, this cosmopolitan species is ideal for clarifying regional effects on the symbiotic association. In the present study, we investigated the diversity of Caballeronia symbionts of L. occidentalis collected in Japan, North America, and Europe in order to confirm whether the geographical origin affects the gut symbionts of the conifer bug. Furthermore, symbiont inoculation tests of insects reared in the laboratory with SBE-α and SBE-β symbionts were conducted to analyze the gut colonization ability and fitness effects of these two Caballeronia subgroups on the host insect.

Materials and Methods
Insects
Samples of L. occidentalis used in the present study are listed in Table 1. Regarding bacterial inoculation tests, L. occidentalis was collected in Gif-sur-Yvette, France in 2018 and maintained in cages by feeding on pignolia nuts and distilled water containing 0.05% ascorbic acid (DWA) at 25°C under a long-day regime (16 h light, 8 h dark).
Table 1.
Information on insect samples investigated in this study.
| Country |
State/Prefecture |
Locality |
Collection year |
Collector |
Symbiont detection |
Specimens number |
Symbiont isolates/
clones number |
16S rRNA Accession number |
| Japan |
Kumamoto |
Koshi |
March, 2021 |
R. Hara, K. Matsunaga |
Isolating |
3 |
3 |
LC713090–LC713092 |
|
Yamagata |
Yuza |
April, 2021 |
Y. Hatanaka |
Isolating |
4 |
4 |
LC713093–LC713096 |
|
Akita |
Akita |
October, 2018 |
K. Takeshita |
Isolating |
2 |
2 |
LC713097–LC713098 |
|
Akita |
Akita |
April, 2021 |
S. Noriyuki |
Isolating |
4 |
4 |
LC713099–LC713102 |
| France |
Essonne |
Gif-sur-Yvettea |
November, 2016 |
P. Mergaert |
Isolating |
3 |
8 |
LC713103–LC713110 |
|
Essonne |
Gif-sur-Yvettea |
November, 2021 |
G. Lextrait |
Isolating |
6 |
22 |
LC713111–LC713132 |
| Italy |
Piedmont |
Alessandria |
October 2020 |
S. Chiesa |
Cloning |
2 |
6 |
LC713133–LC713138 |
| Spain |
Catalonia |
Artes |
April 2021 |
S. Lopez Romero |
Cloning |
3 |
12 |
LC713139–LC713150 |
| USA |
Idaho |
Lenore |
October 2020 |
S. Cook |
Cloning |
4 |
31 |
LC713151–LC713181 |
| Canada |
Nova Scotia |
Vaughan |
October 2020 |
S. Blatt |
Cloning |
4 |
13 |
LC713182–LC713194 |
|
British Col. |
Vernon |
October 2020 |
W. Strong |
Cloning |
4 |
15 |
LC713195–LC713209 |
a The same collection site
Reared insects were used for inoculation tests with one SBE-α strain and one SBE-β strain. We used strain RPE225 (Kikuchi and Fukatsu, 2014), a green fluorescent protein (GFP)-labeled derivative of B. insecticola (Caballeronia insecticola) strain RPE64, as a typical strain of the SBE-α clade. RPE64 was isolated from the midgut crypts of a Japanese specimen of the bean bug Riptortus pedestris (Coreoidea: Alydidae) (Takeshita et al., 2018). Regarding the SBE-β strain, we selected Caballeronia sp. strain 1876, which was isolated from the midgut crypts of L. occidentalis collected in Gif-sur-Yvette, France in 2016. A GFP-labeled derivative of this SBE-β strain, labeled strain 2482, was constructed by a mini-Tn7 transposon delivery system as previously reported (Kikuchi and Fukatsu, 2014). Insect inoculation tests with these two GFP strains were performed as previously described for other stinkbug species (Ohbayashi et al., 2015; Ohbayashi et al., 2019b). Briefly, the two bacterial strains were pre-cultured in yeast extract and glucose (YG) medium (yeast extract 5.0 g L–1, glucose 4.0 g L–1, and NaCl 1.0 g L–1) containing rifampicin 30 μg mL–1 at 28°C overnight at 180 rpm, and 200 μL of the overnight culture was inoculated into 5 mL fresh YG, incubated at 28°C at 180 rpm for 2 h, and finally diluted to 107 CFU mL–1 in DWA. The cotton pad with DWA was removed from the rearing container with 2nd instar L. occidentalis nymphs, and nymphs were maintained overnight without water to make them thirsty. Symbiont suspensions, prepared as described above, were poured onto new cotton pads and placed into rearing containers. Aposymbiotic nymphs were obtained by placing a cotton pad soaked with DWA only. Containers were maintained as described above until later analyses.
Fluorescence microscopy observations
The infection status of the inoculated nymphs was confirmed based on the detection of GFP signals in the dissected midgut crypts of third instar nymphs. Dissections were performed in phosphate-buffered saline (PBS) using fine forceps and scissors under a fluorescent binocular microscope (Leica, MZ FZ III). Pictures of the dissected midguts were taken by a digital camera (Leica, EC3).
Insect survival
The survival rate of aposymbiotic insects and symbiotic insects infected with either SBE-α strain RPE225 or SBE-β strain 2482 was estimated by regularly observing insect rearing populations (n=27, 13 or 20 insects, respectively) over time until the last adult emergence in the surviving insects. At each observation, the number of alive and dead insects was scored, as well as the number of emerged adults. The survival rate was analyzed by Fisher’s exact test with the Bonferroni correction. The developmental time until adulthood in the aposymbiotic insect sample was removed from the statistical analysis due to a single surviving insect (n=1), and those in SBE-α and SBE-β inoculated insects were analyzed by the Student’s t-test.
Competition assay
In the competition assay, we used red fluorescent protein (RFP) strain RPE525 (SBE-α), a derivative of C. insecticola strain RPE64 (Itoh et al., 2019) and GFP strain 2482 (SBE-β). Exponential phase cells were suspended in DWA and an inoculum containing 107 CFU mL–1 of both strains was prepared from them. The inoculation of insects with the mixed inoculum was performed as described above. At 7 days post inoculation, when insects became 3rd instar nymphs, midgut crypts were dissected as described above. In the microscopy analysis, midgut crypts were observed under a fluorescent microscope (Nikon, Eclipse 80i). Regarding the quantitative assessment of the two strains, midgut crypts were collected in 100 μL of PBS buffer in a 1.5-mL tube and homogenized by a sterilized pestle. The pestle was washed by 400 μL of PBS buffer. The relative number of symbiont cells of GFP and RFP bacteria in the extracts of the midgut crypts and in the bacterial suspension of the inoculum were analyzed by flow cytometry (Beckman Coulter, Cytoflex).
Identification of gut symbionts of L. occidentalis
Gut symbionts were isolated from the midgut crypts of L. occidentalis individuals collected in Japan and France (Table 1 and Fig. 1) by plating crypt contents on YG agar plates. Growing bacteria were identified by direct sequencing of the 16S rRNA gene, as previously described (Kikuchi et al., 2011a). Since conifer bugs captured in Italy, Spain, USA, and Canada were preserved in 100% ethanol, their dissected midgut crypts were subjected to DNA extraction and a clone library analysis of the 16S rRNA gene, as previously reported (Ohbayashi et al., 2019b). Sequences obtained by the bacterial isolation and clone analysis were assembled by ATSQ software ver. 5.2 (Software Development), followed by manual corrections. The most similar bacterial species/strains were identified by BLAST comparisons. Sequences showing more than 99% identity were assigned to the same operational taxonomical unit (OTU).
Molecular phylogenetic analysis of L. occidentalis gut symbionts
A multiple sequence alignment of the 16S rRNA gene was constructed with MAFFT on the EMBL-EBI server (Li et al., 2015). A molecular phylogenetic tree was generated by the maximum likelihood (ML) method with the removal of gap-including and ambiguous sites and with a bootstrap analysis (1,000 replicates) in MEGA software version 10.1.8 (Kumar et al., 2018; Stecher et al., 2020). We selected the Tamura-Nei model of nucleotide substitutions with gamma distributed and invariant sites (G+I) (Tamura and Nei, 1993).
Nucleotide sequence accession numbers
The nucleotide sequence data of the 16S rRNA gene obtained in the present study have been deposited in the DDBJ/EMBL/GenBank public databases with the accession numbers LC713090–LC713209 (Table 1).
Results
Identification of gut symbionts in midgut crypts of L. occidentalis
To investigate the diversity of gut symbionts in the conifer bug L. occidentalis, two methods were used depending on the nature of the insect sample. In the bacterial isolation method, 43 symbionts were isolated from the midgut crypts of 22 individuals collected from 4 and 2 populations of Japan and France, respectively (Table 1). In the clone library analysis of the 16S rRNA gene, 17 specimens collected in Italy, Spain, USA, and Canada were used, from which 77 sequences were obtained (Table 1). The 120 assembled sequences were assigned to 11 OTUs based on the 99% sequence identity threshold (Table S1). Seven OTUs (OTU1–OTU7) were identified as members of Caballeronia by a BLAST search. OTU2 and OTU7 represented 69% and 22%, respectively, of the 110 sequences identified as Caballeronia, indicating that OTU2 and OTU7 are the main gut symbionts of L. occidentalis. These dominant OTUs were detected in both the bacterial isolation and cloning methods, which suggests no method-related bias in the identification of symbionts. The remaining four OTUs (OTU8–OTU11), detected only in four individuals of two insect populations, were identified as Rickettsia spp., which is a well-known intracellular secondary symbiont of diverse insects (Kikuchi, 2009). The insect specimen in which Rickettsia clones were identified, yielded a majority of Caballeronia clones (Table S1), indicating that these individuals were also colonized with Caballeronia symbionts.
Phylogenetic placement of Caballeronia symbionts
We performed a molecular phylogenetic analysis based on the 16S rRNA gene, including sequences of the seven Caballeronia OTUs of L. occidentalis gut symbionts, type strains of Burkholderia sensu lato species (Burkholderia sensu strico, Paraburkholderia, and Caballeronia species), and previously reported Caballeronia gut symbionts of various coreoid insects. Based on phylogenetic divergence, four subclades were identified within the Caballeronia clade: SBE-α, SBE-β, SBE-γ, and Coreoidea clade (Fig. 2). The most dominant OTU2, detected in specimens collected in all countries, was located in SBE-β. SBE-β contained four other OTUs (OTU3, OTU4, OTU5, and OTU6) in addition to OTU2 (Fig. 2). On the other hand, the second most dominant OTU7 was placed in the Coreoidea clade (Fig. 2). This subclade contained neither environmental isolates nor type species, but formed a monophyletic group with many gut symbionts of Japanese and European coreoid bugs (Fig. 2), suggesting their very specialized nature for symbiosis with stinkbug species. OTU1 was located in the SBE-α clade, known as major gut symbionts of Japanese and American coreoid stinkbugs (Fig. 2 and S1).

The detection rates of the three subclades (SBE-α, SBE-β, and the Coreoidea clade) in the world’s populations of conifer bugs are summarized in Fig. 3 and S2 (also see Table S2), which are based on the phylogenetic placement of seven OTUs in the subclades of Caballeronia (Fig. 2). Overall, conifer bugs were almost exclusively associated with SBE-β, while SBE-α was rarely detected (Fig. 3). The Coreoidea clade occupied one fourth of the populations and was frequently detected in the Japanese and European populations, but not in the North American populations of conifer bugs (Fig. 3).
Colonization ability and host fitness effects of SBE-α and SBE-β symbiont strains in the midgut crypts of L. occidentalis
SBE-α species have consistently been found in the midgut crypts of 33 stinkbug species of the superfamily Coreoidea (Kikuchi et al., 2011a, 2005; Olivier-Espejel et al., 2011; Garcia et al., 2014; Kuechler et al., 2016; Ohbayashi et al., 2019b; Acevedo et al., 2021; Hunter et al., 2022). However, in the present study, only one out of 120 symbiont isolates or clones from 39 specimens of L. occidentalis were SBE-α (Fig. 3 and Table S1). To confirm whether SBE-α and SBE-β symbionts are capable of colonizing the midgut crypts of the conifer bug, the GFP-labeled strains, C. insecticola strain RPE225 (SBE-α) and Caballeronia sp. strain 2482 (SBE-β) were inoculated into nymphs of L. occidentalis. The SBE-α and SBE-β strains were both capable of colonizing the midgut crypts of L. occidentalis (Fig. 4A, B, D, and E), as indicated by enlarged crypts and the presence of a GFP signal. Moreover, the swollen M4B midgut region, typical for symbiont colonization, also confirmed proper colonization by both strains (Fig. 4A and B). On the other hand, aposymbiotic (uninfected) insects showed small M4 crypts and a narrow M4B region (Fig. 4C and F).
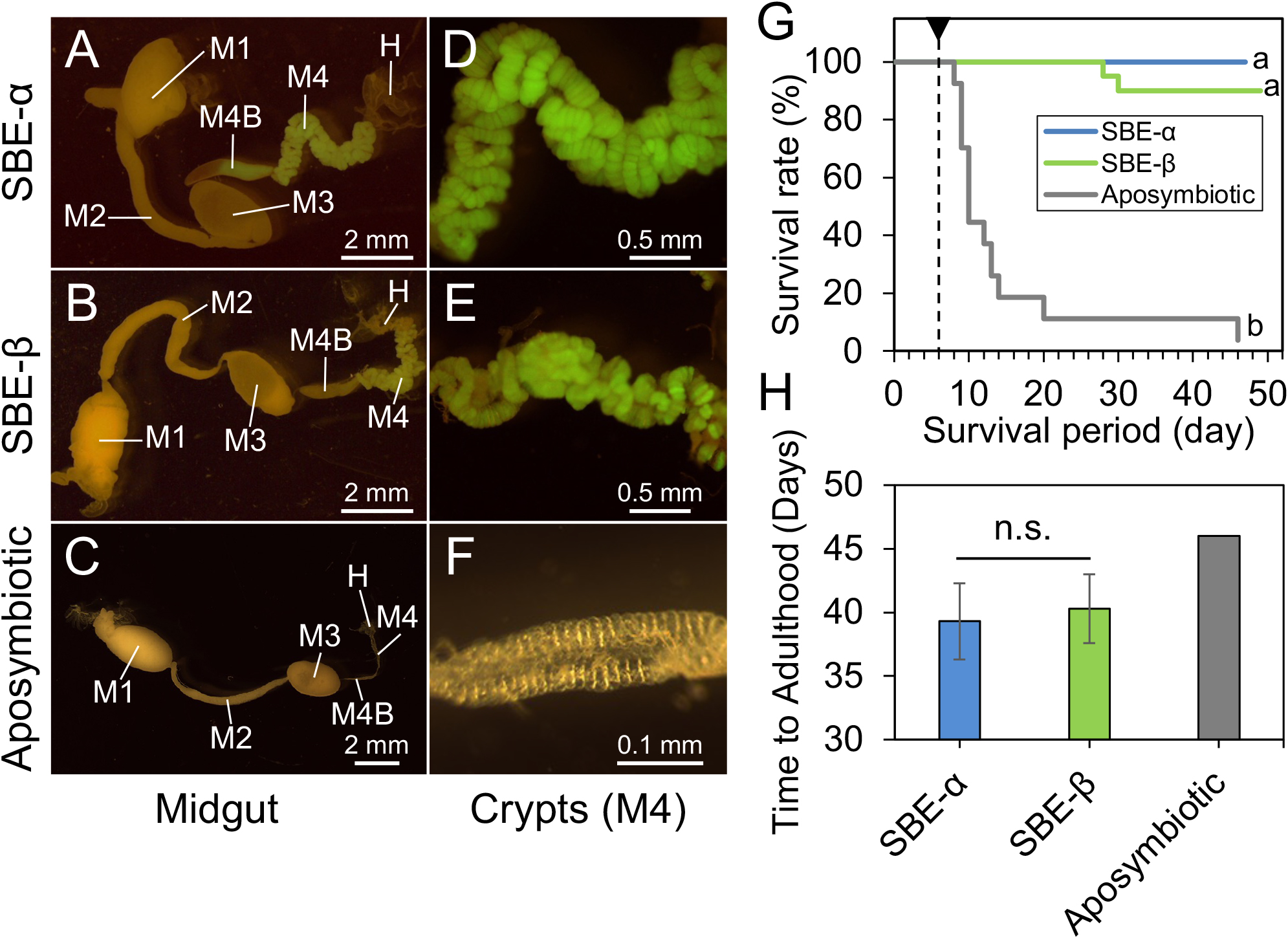
We then investigated the survival of aposymbiotic and SBE-α- or SBE-β-inoculated conifer bugs. Aposymbiotic bugs showed high mortality during their development (Fig. 4G). The majority of insects died during the 2nd to 3rd instar (Fig. 4G). In contrast, most nymphs inoculated with SBE-α or SBE-β strains survived and reached adulthood (Fig. 4G; survival percentage [adult/total investigated insects]=3.7% [1/27] in aposymbiotic insects; 100% [13/13] in SBE-α-inoculated insects; 90% [18/20] in SBE-β-inoculated insects). The developmental times from hatching to adult emergence were 46, 39±3, and 40±3 days (mean±SD) in aposymbiotic, SBE-α-, and SBE-β-inoculated L. occidentalis, respectively (Fig. 4H). Survival rates significantly differed between symbiotic (inoculated with SBE-α or SBE-β) versus aposymbiotic insects, while no significant differences were observed in survival rates or developmental times between SBE-α- and SBE-β-inoculated L. occidentalis (Fig. 4G and H).
On the other hand, a competition assay, in which nymphs were inoculated with an equal mixture of the SBE-α and SBE-β strains, demonstrated that SBE-β significantly outcompeted SBE-α in the midgut crypts of L. occidentalis (Fig. 5). Collectively, these results strongly suggest that the Caballeronia symbiont makes a very large positive contribution to the survival and development of L. occidentalis, and that SBE-α has a sufficient ability to colonize midgut crypts to give the same fitness effects to L. occidentalis as SBE-β. Nevertheless, microbe-microbe competition in the midgut crypts of L. occidentalis may contribute to the observed predominance of SBE-β.
Discussion
The present study revealed that although conifer bug specimens are associated with genetically diverse Caballeronia (SBE-Burkholderia) symbionts, members of the subclade SBE-β were dominant in all of the investigated Japanese, European, and North American populations of the conifer bug, and the Coreoidea clade was also frequently found in Japanese and European insects (Fig. 2 and 3, Table S1 and S2). Previous studies reported that Japanese and American species of the stinkbug superfamily Coreoidea were more likely to harbor symbionts belonging to the SBE-α subclade (Kikuchi et al., 2011a; Ohbayashi et al., 2019b); however, we herein demonstrated that SBE-α was rarely detected in the conifer bug, even in Japanese and American populations (Fig. 3, Table S1 and S2). Based on this broad survey of this cosmopolitan species, we concluded that infection was not affected by geographic origins; therefore, it is more likely to be influenced by selection mechanisms in the host insect.
Experimental inoculation tests revealed no significant differences in colonization ability or fitness effects on the host bug between SBE-α and SBE-β symbionts (Fig. 4). However, the competition assay showed that SBE-β outcompeted SBE-α in the midgut crypts of L. occidentalis, possibly resulting in the low detection rate of SBE-α in the conifer bug (Fig. 3, Table S1 and S2). Infection specificity between the bean bug host and Caballeronia symbiont was demonstrated to be largely influenced by the native symbiont’s colonization competitiveness in the midgut. In midgut crypts in co-infection experiments, Caballeronia symbionts always outcompeted the non-symbiont species Paraburkholderia and Pandoraea, which were fully capable of colonizing crypts in the absence of competition species (Itoh et al., 2019). The present study is the first to report competition-based selection in the stinkbug midgut between species of different SBE clades. Since competition-based mechanisms have not yet been elucidated in detail, further studies are warranted.
Among the Caballeronia gut symbionts of L. occidentalis, the second most dominant OTU7 formed a monophyletic group, the Coreoidea clade, with gut symbionts of other Coreoid species, including Cletus rusticus, Plinachtus bicoloripes, Hygia lativentris, Molipteryx fuliginosa, Acanthocoris sordidus (Kikuchi et al., 2011a), Coreus marginatus (Ohbayashi et al., 2019b), and Dicranocephalus albipes (Kuechler et al., 2016). The Coreoidea clade does not include environmental isolates/clones or any named species of Caballeronia, it only consists of gut symbionts of Coreoidea (Fig. 2), strongly suggesting that these symbiont strains are highly specific to the insect group. To date, two strains—one from A. sordidus and the other the OTU7 clone from L. occidentalis described herein—have been isolated as culturable symbionts of this clade. Further studies are needed to reveal the future genomic and physiological features of these Coreoidea clade symbionts in order to clarify why these Coreoidea clade members are specific to this insect group. The intercontinental infection pattern of the Coreoidea clade (Fig. 3) is notable from the viewpoint of evolution. Since L. occidentalis originated from North America (Heidemann, 1910; Koerber, 1963) and recently invaded European and Asian countries, we speculate that L. occidentalis was originally associated with SBE-β and may have become symbiotic with the Coreoidea clade as its distribution expanded.
Caballeronia symbionts make a very large positive contribution to the fitness of the conifer bug (Fig. 4G), similar to that reported for other coreoid stinkbugs, including C. marginatus, L. zonatus, L. phyllopus, and R. pedestris (Kikuchi et al., 2007, 2011a; Ohbayashi et al., 2019b; Hunter et al., 2022). In R. pedestris, our previous transcriptomic study revealed that Caballeronia provided the host with essential amino acids and vitamins by recycling host metabolic waste materials, such as sulfate, allantoin, and urea (Ohbayashi et al., 2019a). These findings suggest that Caballeronia symbionts critically complement these essential nutrients lacking in conifer seeds in L. occidentalis, resulting in high mortality in aposymbiotic nymphs in L. occidentalis. In contrast, aposymbiotic insects in R. pedestris showed retarded growth, a smaller body size, and lower fecundity than symbiotic insects, although these aposymbiotic nymphs were nevertheless able to develop to adulthood with a high survival rate (Kikuchi et al., 2007; Kikuchi and Fukatsu, 2014). Feeding on soybean seeds with high nutritional value may provide sufficient, albeit non-optimal, nutrition for the development and survival of aposymbiotic R. pedestris. Caballeronia symbionts may play more important metabolic roles for hosts that are feeding on nutritionally poor non-leguminous plants, such as L. occidentalis.
Rickettsia was detected in the American and Canadian populations of L. occidentalis (Table S1). Although some members are human pathogens, most members of the genus Rickettsia are facultative intracellular symbionts in many arthropods (Perlman et al., 2006), and these symbionts are maintained by vertical transmission. Since Rickettsia is a reproductive manipulator that causes male-killing and parthenogenesis in many insects (Perlman et al., 2006), a similar function needs to be considered in the North America populations of the conifer seed bug. The bacterial group has rarely been detected from stinkbug species, except for some species of Miridae (Chang and Musgrave, 1970; Caspi-Fluger et al., 2012; Dally et al., 2020), and a broader survey is required to clarify the prevalence of Rickettsia in heteropteran insects.
The present study on L. occidentalis is the first to demonstrate that the specificity of the Caballeronia symbiont is influenced by host species rather than biogeography. The symbiont’s competitiveness in the gut symbiotic organ appears to be pivotal for specificity. Additionally, differences in fitness effects on host bugs between species, as shown in L. zonatus and L. phyllopus (Hunter et al., 2022), may be involved in specificity. To elucidate the mechanisms underlying host-symbiont specificity in stinkbugs in more detail, the following two points need to be clarified in the future. The worldwide distribution of Caballeronia in soil needs to be analyzed because microbial geography is critical in animals that are closely associated with environmentally transmitted beneficial microorganisms. In addition, further experimental inoculation assays, particularly in vivo competition assays, are crucial for confirming and understanding the mechanisms underlying host-microbe specificity in each stinkbug species.
Citation
Ohbayashi, T., Cossard, R., Lextrait, G., Hosokawa, T., Lesieur, V., Takeshita, K., et al. (2022) Intercontinental Diversity of Caballeronia Gut Symbionts in the Conifer Pest Bug Leptoglossus occidentalis. Microbes Environ 37: ME22042.
https://doi.org/10.1264/jsme2.ME22042
Acknowledgements
We thank R. Hara, K. Matsunaga, Y. Hatanaka, S. Noriyuki, S. Chiesa, S. Lopez Romero, S. Cook, S. Blatt, A. Ravenscraft, M. Hunter, and W. Strong for insect sampling, and J. Lachat and A. Yokota for their assistance with insect rearing. The present study has benefited from the Imagerie-Gif core facility supported by l’Agence Nationale de la Recherche (ANR-11-EQPX-0029/Morphoscope, ANR-10-INBS-04/FranceBioImaging; ANR-11-IDEX-0003-02/Saclay Plant Sciences). This study was supported by a JSPS-CNRS Bilateral Open Partnership Joint Research Project and the CNRS International Research Project “Ménage à Trois” to YK and PM, by the French national research agency (ANR) grant ANR-19-CE20-0007 and a Saclay Plant Sciences research grant to PM, and by the JSPS Research Fellowship for Young Scientists (20170267 and 19J01106) to TO, by the MEXT KAKENHI (18KK0211 to YK and TH; 20H03303 to YK and K. Takeshita), and the Moonshot project JPNP18016, commissioned by the New Energy and Industrial Technology Development Organization (NEDO), to TO and K. Tago.
References
- Acevedo, T.S., Fricker, G.P., Garcia, J.R., Alcaide, T., Berasategui, A., Stoy, K.S., and Gerardo, N.M. (2021) The importance of environmentally acquired bacterial symbionts for the squash bug (Anasa tristis), a significant agricultural pest. Front Microbiol
12: 719112.
- Ahn, S.J., Son, D., Choo, H.Y., and Park, C.G. (2013) The first record on Leptoglossus occidentalis (Hemiptera: Coreidae) in Korea, a potential pest of the pinaceous tree species. J Asia-Pac Entomol
16: 281–284.
- Ben Jamaa, M., Mejri, M., Naves, P., and Sousa, E. (2013) Detection of Leptoglossus occidentalis
Heidemann, 1910 (Heteroptera: Coreidae) in Tunisia. Afr Entomol
21: 165–167.
- Beukes, C.W., Palmer, M., Manyaka, P., Chan, W.Y., Avontuur, J.R., van Zyl, E., et al. (2017) Genome data provides high support for generic boundaries in Burkholderia sensu lato. Front Microbiol
8: 1154.
- Caspi-Fluger, A., Inbar, M., Mozes-Daube, N., Katzir, N., Portnoy, V., Belausov, E., et al. (2012) Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc Biol Sci
279: 1791–1796.
- Chang, K.P., and Musgrave, A.J. (1970) Ultrastructure of rickettsia-like microorganisms in the midgut of a plant bug, Stenotus binotatus Jak. (Heteroptera: Miridae). Can J Microbiol
16: 621–622.
- Dally, M., Lalzar, M., Belausov, E., Gottlieb, Y., Coll, M., and Zchori-Fein, E. (2020) Cellular localization of two Rickettsia symbionts in the digestive system and within the ovaries of the Mirid Bug, Macrolophous pygmaeus. Insects
11: 530.
- Dobritsa, A.P., and Samadpour, M. (2016) Transfer of eleven species of the genus Burkholderia to the genus Paraburkholderia and proposal of Caballeronia gen. nov. to accommodate twelve species of the genera Burkholderia and Paraburkholderia. Int J Syst Evol Microbiol
66: 2836–2846.
- Dusoulier, F., Lupoli, R., Aberlenc, H.P., and Streito, J.C. (2007) L’invasion orientale de Leptoglossus occidentalis en France: bilan de son extension biogéographique en 2007 (Hemiptera Coreidae). L’Entomologiste
63: 303–308 (in French with an English abstract).
- Eberl, L., and Vandamme, P. (2016) Members of the genus Burkholderia: good and bad guys. F1000Research
5: 1007.
- Engel, P., and Moran, N.A. (2013) The gut microbiota of insects-diversity in structure and function. FEMS Microbiol Rev
37: 699–735.
- Estrada-de Los Santos, P., Palmer, M., Chavez-Ramirez, B., Beukes, C., Steenkamp, E.T., Briscoe, L., et al. (2018) Whole genome analyses suggests that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes
9: 389.
- Faúndez, E.I., Rocca, J.R., and Villablanca, J. (2017) Detection of the invasive western conifer seed bug Leptoglossus occidentalis
Heidemann, 1910 (Heteroptera: Coreidae: Coreinae) in Chile. Arquivos Entomolóxicos
17: 317–320.
- Fent, M., and Kment, P. (2011) First record of the invasive western conifer seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Turkey. North West J Zool
7: 72–80.
- Gapon, D.A. (2013) First records of the western conifer seed bug Leptoglossus occidentalis Heid. (Heteroptera, Coreidae) from Russia and Ukraine, regularities in its distribution and possibilities of its range expansion in the palaearctic region. Entomol Rev
93: 174–181.
- Gapon, D.A. (2015) First record of Leptoglossus occidentalis (Heteroptera: Coreidae) in Morocco. Heteropterus Rev Entomol
15: 161–163.
- Garcia, J.R., Laughton, A.M., Malik, Z., Parker, B.J., Trincot, C., Chiang, S.L., et al. (2014) Partner associations across sympatric broad-headed bug species and their environmentally acquired bacterial symbionts. Mol Ecol
23: 1333–1347.
- Heidemann, O. (1910) New species of Leptoglossus from North America. Proc Entomol Soc Wash
12: 191–197.
- Hunter, M.S., Umanzor, E.F., Kelly, S.E., Whitaker, S.M., and Ravenscraft, A. (2022) Development of common leaf-footed bug pests depends on the presence and identity of their environmentally-acquired symbionts. Appl Environ Microbiol
88: e01778-21.
- Ishikawa, T., and Kikuhara, Y. (2009) Leptoglossus occidentalis Heidemann (Hemiptera: Coreidae), a presumable recent invader to Japan. Jpn J Entomol
12: 115–116 (in Japanese with an English abstract).
- Itoh, H., Tago, K., Hayatsu, M., and Kikuchi, Y. (2018) Detoxifying symbiosis: microbe-mediated detoxification of phytotoxins and pesticides in insects. Nat Prod Rep
35: 434–454.
- Itoh, H., Jang, S., Takeshita, K., Ohbayashi, T., Ohnishi, N., Meng, X.Y., et al. (2019) Host-symbiont specificity determined by microbe-microbe competition in an insect gut. Proc Natl Acad Sci U S A
116: 22673–22682.
- Kikuchi, Y., Meng, X.Y., and Fukatsu, T. (2005) Gut symbiotic bacteria of the genus Burkholderia in the broad-headed bugs Riptortus clavatus and Leptocorisa chinensis (Heteroptera: Alydidae). Appl Environ Microbiol
71: 4035–4043.
- Kikuchi, Y., Hosokawa, T., and Fukatsu, T. (2007) Insect-microbe mutualism without vertical transmission: a stinkbug acquires a beneficial gut symbiont from the environment every generation. Appl Environ Microbiol
73: 4308–4316.
- Kikuchi, Y. (2009) Endosymbiotic bacteria in insects: their diversity and culturability. Microbes Environ
24: 195–204.
- Kikuchi, Y., Hosokawa, T., and Fukatsu, T. (2011a) An ancient but promiscuous host-symbiont association between Burkholderia gut symbionts and their heteropteran hosts. ISME J
5: 446–460.
- Kikuchi, Y., Hosokawa, T., and Fukatsu, T. (2011b) Specific developmental window for establishment of an insect-microbe gut symbiosis. Appl Environ Microbiol
77: 4075–4081.
- Kikuchi, Y., Hayatsu, M., Hosokawa, T., Nagayama, A., Tago, K., and Fukatsu, T. (2012) Symbiont-mediated insecticide resistance. Proc Natl Acad Sci U S A
109: 8618–8622.
- Kikuchi, Y., and Fukatsu, T. (2014) Live imaging of symbiosis: spatiotemporal infection dynamics of a GFP-labelled Burkholderia symbiont in the bean bug Riptortus pedestris. Mol Ecol
23: 1445–1456.
- Kikuchi, Y., Ohbayashi, T., Jang, S., and Mergaert, P. (2020) Burkholderia insecticola triggers midgut closure in the bean bug Riptortus pedestris to prevent secondary bacterial infections of midgut crypts. ISME J
14: 1627–1638.
- Koerber, T.W. (1963) Leptoglossus occidentalis (Hemiptera, Coreidae), a newly discovered pest of coniferous seed. Ann Entomol Soc Am
56: 229–234.
- Kuechler, S.M., Matsuura, Y., Dettner, K., and Kikuchi, Y. (2016) Phylogenetically diverse Burkholderia associated with midgut crypts of spurge bugs, Dicranocephalus spp. (Heteroptera: Stenocephalidae). Microbes Environ
31: 145–153.
- Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol
35: 1547–1549.
- Lesieur, V., Yart, A., Guilbon, S., Lorme, P., Auger-Rozenberg, M.-A., and Roques, A. (2014) The invasive Leptoglossus seed bug, a threat for commercial seed crops, but for conifer diversity?
Biol Invasions
16: 1833–1849.
- Lesieur, V., Lombaert, E., Guillemaud, T., Courtial, B., Strong, W., Roques, A., and Auger-Rozenberg, M.A. (2018) The rapid spread of Leptoglossus occidentalis in Europe: a bridgehead invasion. J Pest Sci
92: 189–200.
- Li, W., Cowley, A., Uludag, M., Gur, T., McWilliam, H., Squizzato, S., et al. (2015) The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res
43: W580–584.
- Lin, Q.H., Lv, Y.Y., Gao, Z.H., and Qiu, L.H. (2020) Pararobbsia silviterrae gen. nov., sp. nov., isolated from forest soil and reclassification of Burkholderia alpina as Pararobbsia alpina comb. nov. Int J Syst Evol Microbiol
70: 1412–1420.
- Lopes-Santos, L., Castro, D.B.A., Ferreira-Tonin, M., Correa, D.B.A., Weir, B.S., Park, D., et al. (2017) Reassessment of the taxonomic position of Burkholderia andropogonis and description of Robbsia andropogonis gen. nov., comb. nov. Antonie van Leeuwenhoek
110: 727–736.
- Malumphy, C., Botting, J., Bantock, T., and Reid, S. (2008) Influx of Leptoglossus occidentalis Heidemann (Coreidae) in England. Het News Newsletter of the UK Heteroptera Recording Schemes 2nd Series 12 pp. 7–9. URL https://www.britishbugs.org.uk/HetNews/Issue%2012_Autumn%202008_2422Kb.pdf
- Moran, N.A., Ochman, H., and Hammer, T.J. (2019) Evolutionary and ecological consequences of gut microbial communities. Annu Rev Ecol Evol Syst
50: 451–475.
- Ohbayashi, T., Takeshita, K., Kitagawa, W., Nikoh, N., Koga, R., Meng, X.Y., et al. (2015) Insect’s intestinal organ for symbiont sorting. Proc Natl Acad Sci U S A
112: E5179–E5188.
- Ohbayashi, T., Futahashi, R., Terashima, M., Barriere, Q., Lamouche, F., Takeshita, K., et al. (2019a) Comparative cytology, physiology and transcriptomics of Burkholderia insecticola in symbiosis with the bean bug Riptortus pedestris and in culture. ISME J
13: 1469–1483.
- Ohbayashi, T., Itoh, H., Lachat, J., Kikuchi, Y., and Mergaert, P. (2019b) Burkholderia gut symbionts associated with European and Japanese populations of the dock bug Coreus marginatus (Coreoidea: Coreidae). Microbes Environ
34: 219–222.
- Olivier-Espejel, S., Sabree, Z.L., Noge, K., and Becerra, J.X. (2011) Gut microbiota in nymph and adults of the giant mesquite bug (Thasus neocalifornicus) (Heteroptera: Coreidae) is dominated by Burkholderia acquired de novo every generation. Environ Entomol
40: 1102–1110.
- Perlman, S.J., Hunter, M.S., and Zchori-Fein, E. (2006) The emerging diversity of Rickettsia. Proc Biol Sci
273: 2097–2106.
- Salem, H., Bauer, E., Strauss, A.S., Vogel, H., Marz, M., and Kaltenpoth, M. (2014) Vitamin supplementation by gut symbionts ensures metabolic homeostasis in an insect host. Proc Biol Sci
281: 20141838.
- Sawana, A., Adeolu, M., and Gupta, R.S. (2014) Molecular signatures and phylogenomic analysis of the genus Burkholderia: proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front Genet
5: 429.
- Stecher, G., Tamura, K., and Kumar, S. (2020) Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol Biol Evol
37: 1237–1239.
- Sudakaran, S., Kost, C., and Kaltenpoth, M. (2017) Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol
25: 375–390.
- Takeshita, K., and Kikuchi, Y. (2017) Riptortus pedestris and Burkholderia symbiont: an ideal model system for insect-microbe symbiotic associations. Res Microbiol
168: 175–187.
- Takeshita, K., Tamaki, H., Ohbayashi, T., Meng, X.-Y., Sone, T., Mitani, Y., et al. (2018) Burkholderia insecticola sp. nov., a gut symbiotic bacterium of the bean bug Riptortus pedestris. Int J Syst Evol Microbiol
68: 2370–2374.
- Tamura, K., and Nei, M. (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol
10: 512–526.
- Taylor, S.J., Tescari, G., and Villa, M. (2001) A nearctic pest of Pinaceae accidentally introduced into Europe: Leptoglossus occidentalis (Heteroptera: Coreidae) in northern Italy. Entomol News
112: 101–103.
- Tsuru, T.K., Yuito, O., and Akio, T. (2020) First record of alien leaf-footed bug, Leptoglossus occidentalis (Insecta, Hemiptera, Coreidae) from Tottori Prefecture, western Japan, with remarks on its range expansion in Japan. Bulletin of the Tottori Prefectural Museum
57: 37–43 (in Japanese with an English abstract).
- van der Heyden, T. (2019) Summarized data on the European distribution of Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae: Coreinae: Anisoscelini). Rev Chil Entomol
45: 499–502.
- Yabuuchi, E., Kosako, Y., Oyaizu, H., Yano, I., Hotta, H., Hashimoto, Y., et al. (1992) Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol Immunol
36: 1251–1275.

