Review
Information Efficiency in E. coli Chemotaxis
2022 Volume 62 Issue 5 Pages 276-279
Details
2022 Volume 62 Issue 5 Pages 276-279
生物は感覚情報を利用して行動上の意思決定を行う.情報という抽象的な量と,実際の生物の行動のパフォーマンスにはどのような関係があるのだろうか.筆者らは最近,大腸菌の走化性行動をモデル系として,細胞が環境から獲得する情報量と行動のパフォーマンスの関係を解析した.主要な結果をここで平易に解説する.
Organisms acquire and use sensory information to guide their behaviors. However, it is unclear whether and how this information constrains the ability of organisms to perform behavioral tasks. In a recent work, using E. coli chemotaxis as a model system, we showed that the sensory information that a bacterium acquires sets an upper limit on its behavioral performance. Furthermore, combined with quantitative experiments, we quantified the rate at which E. coli acquire information during navigation and discovered that E. coli use the acquired information efficiently. Here, we succinctly review the key findings of the work.
生物は環境から感覚器を通して情報を獲得し,これを運動の制御に利用することで生物学的なタスクを遂行する.例えば獲物を追いかけるチーターは,視覚により獲物の位置情報を得て走る方向や速度を制御する.生物の適応的な行動の背後には,明らかに高度に組織化された感覚系と運動系の連携がある.感覚運動系(sensorimotor systems)が適応的に振る舞う時,それは系のどのような性質によるのだろうか.これを理解するためには,感覚系が検出し処理する「情報」の概念を定量的に扱い,これを運動の結果としての行動のパフォーマンスに関連付けることが必要であるように思われる.
情報の概念は情報理論により定量化される1),2).これを生物系の解析に用いた研究は,生物の感覚系が理論限界に近い情報量を環境から引き出し得ることを明らかにしてきた3).しかしながら感覚系が獲得する情報の多寡は,その個体の行動のパフォーマンスの高低に必ずしも直結しない.感覚系を通して得られる全ての情報がその個体が行うタスクに関連するわけではないし,タスクに関連する情報が得られたとしても運動系がそれをタイムリーに利用しなければパフォーマンスには繋がらないからである4).生物はタスクを遂行する際,どれくらいの情報を環境から獲得するのだろうか.また獲得した情報のうちどれくらいをそのタスクの遂行に使うのだろうか.
筆者は最も単純な感覚運動系のモデルとして大腸菌の走化性を調べてきた5)-8).大腸菌は細胞外の走化性誘引分子(リガンド)を膜上の受容体で検出する.リガンド濃度に空間的な勾配がある環境では,大腸菌は,空間移動により濃度の空間勾配の情報を自身が感知する濃度の時間変化の情報に変換する.この情報は細胞内で一連の化学反応(シグナリング系)により処理され,べん毛モーターの状態制御,ひいては細胞の直進運動・方向転換運動という運動状態の制御に利用される.このシステム全体は,細胞が感知するリガンド濃度が上昇するとその直進運動の時間が増加するようにデザインされており,その結果,細胞はリガンド濃度の空間勾配を上る性質,つまり走化性を示す9),10).
この感覚運動系の性質を理解するには,ちょうど行動神経科学が神経活動の測定を基本的なツールとするように,細胞内部の化学反応動態を測定する技術が必須だろう.筆者は蛍光観察,特に蛍光共鳴エネルギー移動(FRET)測定の技術を軸としてこのような実験系の開発に携わってきた5),6)(図1).最近Mattinglyと神野ら8)(以下,筆者ら)は,この実験系を応用することで,一つの大腸菌細胞が走化性において環境から毎秒獲得する情報の量を測定し,さらにその情報が行動へ利用される効率を評価した.本稿では,情報理論等の知識を仮定せずこの論文の主要な結果を解説する.
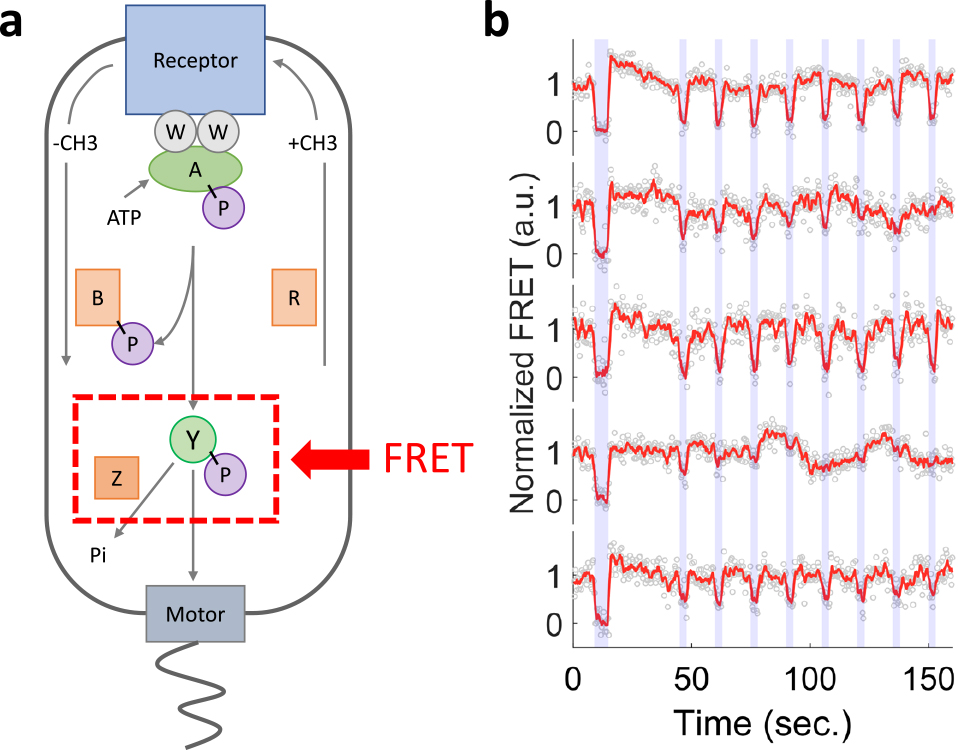
a)大腸菌の走化性シグナリング経路.本文中のFRET測定では,受容体と複合体を形成するキナーゼ(CheA)によりリン酸化される応答制御因子(CheY)とその脱リン酸化酵素(CheZ)を蛍光標識し,それらの間の相互作用の程度をFRETシグナルから推定する.この分子間相互作用はキナーゼの活性度に比例するため,FRETによりキナーゼの活性度を知ることができる.b)一細胞FRET測定により得られるキナーゼ活性動態(赤).マイクロ流体デバイスにより瞬時に(~0.1秒)リガンド濃度を切り替え,それに対する応答を観察することができる.青で示した背景はリガンド濃度を上昇させた時間帯を示す.
ある系が獲得する情報とパフォーマンスの関係を直感的に理解するために,Netflixなどのビデオストリーミングサービスを使って手元のコンピュータで動画を再生する状況を考える.コンピュータはインターネット回線を通してbits/sの単位で量られる動画の情報を連続的に獲得する.ここでは動画再生のパフォーマンスの指標として動画の解像度を考える.
この系の状態はインターネット速度と解像度で特徴付けられるだろう(図2).高い解像度を達成するために,速いインターネットは必要ではあるが十分ではない.動画と関係のない情報は解像度には貢献し得ないし,動画に関する情報は適切に利用されなければならないからだ.有限のインターネット速度環境では,これをなるべく効率良く利用するよう(例えばデータ転送の仕組みを)工夫することで解像度を高められるだろう.しかし工夫にも限度がある.一定のインターネット速度の下で達成可能な解像度には,理論的な限界があるはずだ.この理論限界は,インターネット速度と解像度の二軸からなる平面を達成可能・不可能な領域に分割する一つの曲線として表現できる(図2;緑線).コンピュータがこの理論限界の近くで作動している時,動画の解像度は情報により制限されていると言うことができるだろう.そこから解像度をさらに高めるためには,インターネット速度,つまり情報量を高めるしか方法がないからである.
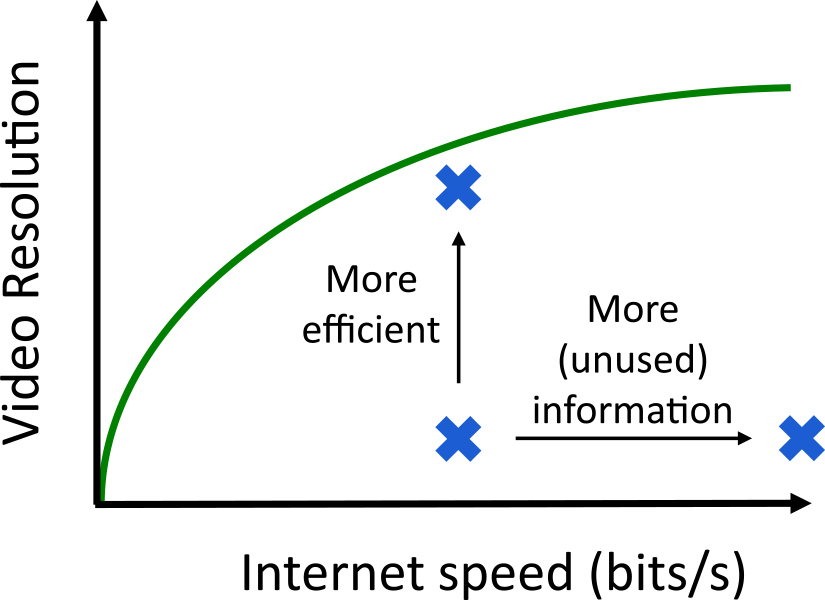
ビデオストリーミングにおける情報量(Internet speed)とパフォーマンス(Video resolution)の関係.特定のシステムはこの二軸からなる平面上の一点として特徴付けられるが,達成可能な領域は理論限界(緑線)の下の領域に限られる.
ストリーミングメディアの再生を例に考察した情報とパフォーマンスの関係は,生物の行動にも適用できるだろう3).生物は,やはりbits/sの単位で記述される感覚情報を環境から獲得し,これを利用してタスクを遂行する.この状況は,少なくとも原理的には個体が獲得する情報レートと行動のパフォーマンスにより特徴付けられる.そして生物の場合もやはり,一定の情報量の下では達成可能なパフォーマンスに理論的な限界があるだろう.
筆者らは,大腸菌の走化性においてこの理論限界が具体的にどのようなものであり,また実際の大腸菌がこの理論限界に比べてどの程度のパフォーマンスを達成するのかを問うた.走化性の文脈では,パフォーマンスはドリフトスピードvd,つまり濃度勾配方向の平均移動スピードで評価できるだろう.この量が,大腸菌が走化性を示す際に得る情報量―より正確には単位時間あたりに得るリガンドの濃度変化に関する情報(情報レート)İ―にどのように依存しているのか,という問題設定である.
まず理論限界を導くために,筆者らは大腸菌の走化性行動の数理モデルを立てこれを解析した.このモデルは,分子的詳細は捨象するものの,行動レベルで知られている性質を網羅した詳細なものである(例えばモデルにおける「直進」運動は,実際の細胞と同様,以下の式でDrで表現される回転拡散の効果により完全な直進ではない).このモデルから,筆者らはまずvdとİを導いた.導かれたvdとİは,任意の(汎)関数として表現される細胞のリガンド応答特性―細胞が感知したリガンド濃度の時系列を運動状態の時系列にどのようなルールで変換するか―にそれぞれ異なる仕方で依存する.ここで筆者らが解きたい問題は,İをある値に固定した時のvdの理論的上限値,つまり物理・化学的制約を外してあらゆるリガンド応答特性が可能とした場合に最も高いvdの値はどれくらいか,という問題である.この最適化問題を解くことで,達成可能なvdの値は次の不等式により規定されることが分かる:
ここでv0は直進運動時のスピード(実際の大腸菌では約20 μm/s),f(θ)は0から1の間の値を取る関数で走化性行動を特徴付けるパラメタのセットθに依存する.
筆者らはこの式の右辺に現れるパラメタ(θ, Dr)の値を大腸菌の遊泳行動を観察することで測定し,上式右辺の曲線を決定した(図3緑線).この曲線を見ることで,あるİが与えられた時にどれくらいのvdが理論的に達成可能なのか,また逆に,あるvdを達成するためにはどれくらいのİが必要なのか,を知ることができる.先行研究において,大腸菌が走化性を示すのに必要な情報レートİは1 bit/sのオーダーであると推定されていた11).この推定の根拠は,大腸菌は約1秒ごとに繰り返される直進運動の度に濃度勾配を上っているか否か(つまり直進運動時間を延ばすべきか縮めるべきか)を判断している,つまり,1 bit―二つの可能性から一つを選ぶのに必要な情報量―を要求するbinary decisionを毎秒行っている,というものだった.ところがこの理論曲線は,その1/100程度のわずかな情報レートで直進運動時のスピードv0の数%程度のドリフトスピード―1秒あたり細菌の体長程度移動するスピード―が達成可能であることを示している.すなわち,上述のbinary decisionの「正答率」はかなり低くても走化性は可能なのである.
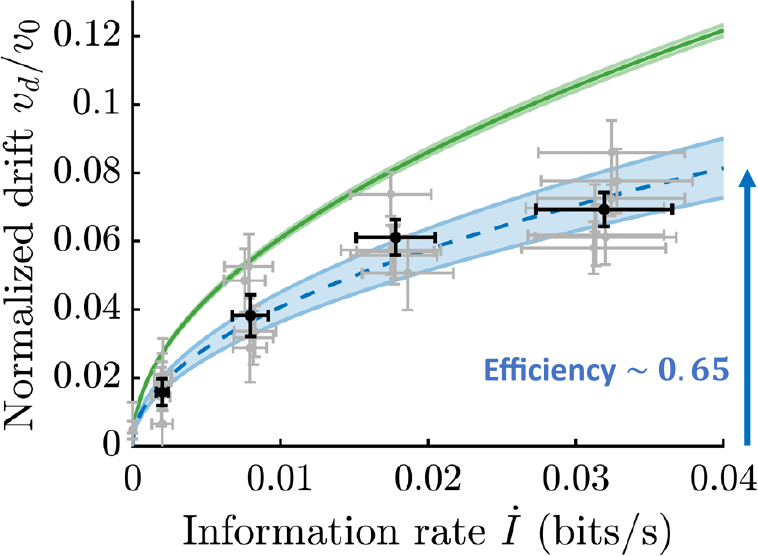
大腸菌の走化性における情報とパフォーマンス(ドリフトスピード)の関係.モデルの解析により得られる理論限界(緑)と実際の細胞からの測定により決まる曲線(青)を示す.大腸菌の走化性は理論限界と同程度(~65%)のパフォーマンスを示し,効率の高い情報処理システムであることが分かる.
実際の細胞のパフォーマンスは上述の理論限界と比較してどの程度なのだろうか.理論解析によると,細胞が示すリガンド応答が無限に速くかつ完全にノイズレスな場合にのみ,その細胞のパフォーマンスvdは理論限界に到達することができる.実際の細胞のリガンド応答は,化学反応からなるノイジーで有限時間を要する現象であるため,そのパフォーマンスvdは理論限界よりも必ず小さい.しかしながら,それはどれくらい小さいのだろうか.
これを知るために,筆者らは大腸菌の走化性におけるvdとİを測定した.vdは,チャンバー中にリガンド濃度勾配を形成し,その中での大腸菌の走化性行動を観察することで比較的容易に測定できる.一方İの測定は,細胞が感知するリガンド濃度の時間変化と細胞内状態の時間変化の相関を決めることを意味し,より繊細な実験を要求する.筆者らは図1に示したFRETによる一細胞測定系を用いることで,シグナリング系のリガンド応答特性とそのノイズの統計的性質を測定し,İを決定した.
これらの測定により,vdとİがそれぞれリガンド濃度勾配ɡの関数として得られた.この二つの関数からɡを消去することで,İの関数としてvdが求まる(図3).これを先に求めたvdの理論限界と比較した結果,驚くべきことに,実際の細胞は理論限界と同程度(~65%)のパフォーマンスvdを示すことが分かった.これは「走化性を示すために理論的に必要な情報量」と「細胞が走化性において実際に獲得する情報量」が同程度であることを意味する.これらが互いに近くならなければならない必然的な理由はない:理論限界を示す曲線はマクロな行動レベル(~10 μm)のモデルと測定により決定され,実際の細胞を記述する曲線は,ミクロな分子レベル(~10 nm)の測定により決定されたものである.にも関わらずこれらの曲線が互いに近いことは,大腸菌の走化性シグナリング系が,細胞内分子動態にエンコードされたリガンドの情報を非常に効率良く用いるようにデザインされていることを示唆している.
上述の結果はいくつかの問いを誘発する.まず大腸菌の高い情報利用効率は,どのようなメカニズムによっているのだろうか.一般に情報利用効率を下げる要因は,(i)パフォーマンスに寄与し得ないirrelevantな情報の獲得,及び(ii)獲得したrelevantな情報を処理・伝達する際に生じる情報の散逸,の二点である.今回の研究は(i)と(ii)の総和が小さいことを明らかにしたが,これらの比率や,それぞれを小さく抑えるメカニズムについては答えていない.これらを明らかにするのは今後の課題である.
今回見出された高い情報利用効率は,どの程度細胞の情報処理に普遍的な性質なのだろうか.細胞のスケールにおいてはノイジーな分子的環境から感覚情報を獲得することは難しく,それ故に獲得した情報をより有効に利用する細胞が進化的に選択されるという可能性が考えられる.今回の研究は,特定の細胞株の特定のリガンドに対する走化性行動のみを対象としているため,結果の一般性について言えることは限られている.今後の方向として,同様の解析を別の系に適用し結果を比較するのは興味深いだろう.細胞の行動,シグナリングにおいてはしばしばその多様性が強調される12),13)が,これらを情報の観点から眺めることで,共通性が見えてくるかもしれない.
技術面にも課題がある.今回の研究は,技術的には細菌の内部状態がFRETにより一細胞レベルで観察できるようになったことで実現可能となった.この背景には,イメージング関連機器や蛍光タンパク質などの性能の向上がある.細菌の行動やシグナリングは比較的単純で数理的な解析と相性が良く,より定量的な一細胞測定が可能になることで,その理解は加速するだろう.しかしながら細菌一細胞の蛍光(特にFRET)観察は,未だに測定パラメタのファイン・チューニングを要するチャレンジングな実験である.その最大の原因は,体積の小さい細菌は細胞内に発現できる蛍光タンパク質の絶対量が小さく,細菌の103倍程度の体積を持つ真核細胞に比べて(例えば文献14, 15)圧倒的にデータのSN比が低いことにある.筆者はこの状況を改善すべく,機械学習の技術を使って,情報理論的に最適な形で生データからFRETシグナルを抽出する方法の開発に取り組んでいる16).細菌一細胞レベルのシグナリング動態をより精密に測定し特徴付けることで,我々は細胞の情報処理をより深く理解できるようになるだろう.
本稿は,Henry H. Mattingly,Benjamine B. Machta,Thierry Emonet(全てイェール大学)と共同で行った研究成果8)に基づく.
神野圭太(かみの けいた)
中央研究院 分子生物研究所(Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan)Assistant Research Fellow
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/62-5ie2p01.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)