Karyotype Report
Novel karyotype of triploid Taraxacum officinale Weber ex F. H. Wigg. s.l. (common dandelion; Asteraceae) in Japan
2024 年 89 巻 1 号 p. 71-73
詳細
2024 年 89 巻 1 号 p. 71-73
A new karyotype in the form of 2n=24=19(M+m)+1mcs+1mcscs+3sm was identified in Japanese triploid Taraxacum officinale Weber ex F. H. Wigg. s.l. Such a line can diversify and evolve without sexual intervention. These findings indicate that karyotypic diversities of apomictic Taraxacum are increased by the occurrence of chromosome mutations, such as structural changes in chromosomes. The present study adds to knowledge of the speciation of apomictic plants.
The genus Taraxacum classified in the family Asteraceae is represented by about 400 species, of which 15 species occur in Japan (Morita 2017). Taraxacum is a monobasic genus with x=8 (Darlington and Wylie 1955), commonly known as a complex taxonomic group, and many of its species are difficult to distinguish from one another. Hybridization and agamospermy (asexual reproduction through seeds) are common in the genus and contribute to its taxonomic complexity. The great majority of polyploid Taraxacum species are obligate agamospermous, although diploids are always sexual (Richards 1997).
The common dandelion (Taraxacum officinale Weber ex F. H. Wigg. s.l.) is one of the best-known introduced perennial plants widely distributed in Japan. The cosmopolitan species exhibits a range of ploidy levels, from diploidy (2x) to hexaploidy (6x). However, in Japan, only triploid (2n=3x=24) and tetraploid (2n=4x=32) plants are found (Sato et al. 2014).
Sato et al. (2014) investigated the geographic distribution of these two cytotypes of T. officinale s.l. and revealed that triploid and tetraploid plants coexist and have a wide distribution in the lowlands of Honshu, Japan. They yielded a total of eight forms of 3x T. officinale s.l. (Types 1 to 8) and four forms of 4x T. officinale s.l. (Types 9 to 12). Karyotypes 1 and 7 have also been reported by Sato et al. (2007b).
Normally, Taraxacum plants are known to possess characteristic satellited chromosomes (Mogie and Richards 1983). The satellited chromosome, which has one large satellite in the long or short arm, has attracted considerable attention because of its distinctiveness and is referred to as ‘H’ (Sørensen and Gudjónsson 1946), ‘Type VI’ (Malecka 1962), ‘1.8+1.0’ (Richards 1972), or ‘Taraxacum type’ (Mogie and Richards 1983). The satellited chromosome usually occurs at a frequency of one per genome (i.e., haploid set). The number of satellites corresponds to the number of secondary constrictions on the chromosome arms. In the Japanese dandelion, native sexual diploid species also have one chromosome with a large satellite per genome (Sato et al. 2007a).
Cytological investigations can provide useful information on the genetic diversity of agamospermous polyploid Taraxacum species. Here we report a new triploid karyotype detected by karyotype analysis.
Triploid T. officinale s.l. plants (2n=3x=24) used in the present study overlap with those previously examined by Sato et al. (2008) collected from Shiga Heights in Nagano Prefecture of central Honshu, Japan. Plant materials were collected at 1,300 m altitude. All 34 individuals were potted and maintained in the botanical garden of the Department of Science, University of Toyama.
For mitotic studies, root tips were collected from potted plants, pretreated with 2.1 mM 8-hydroxyquinoline for 1 h at ca. 25°C, and kept for 15 h at 5°C. Fixation was performed in an ethyl alcohol : glacial acetic acid (3 : 1) solution for 1 h at ca. 25°C. Fixed root tips were macerated in 1M hydrochloric acid at ca. 25°C for 1 h and at 60°C for 10 min, and then washed in tap water. Meristems were stained with a drop of 1.5% lacto-propionic orcein on a glass slide, and an ordinary squash method was applied in preparation.
In each individual, a minimum of 10 metaphase plates were studied to determine karyotypes. Well-spread metaphase plates were analyzed and photographed. Cells with fully spread metaphase chromosomes were used to describe chromosomal forms according to the nomenclature of Levan et al. (1964), which is based on the position of the centromere.
The chromosome numbers of the 34 individuals were found to be 2n=24, consistent with the previous determination by Sato et al. (2008) (Fig. 1A, B). The chromosome morphology differed between metacentric (M and m) and submetacentric (sm) classes. As a result of karyotyping, two distinctly different karyotypes (SH1 and SH2) were observed (Table 1). The details are provided below.
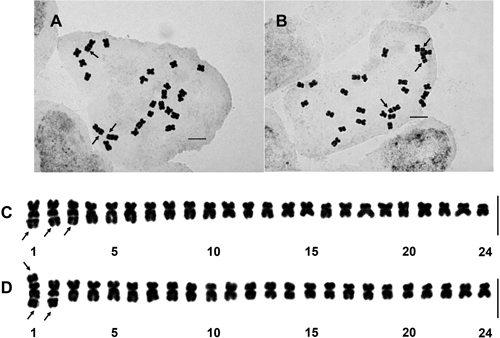
(A), (C): karyotype SH1; (B), (D): karyotype SH2. Arrows indicate satellites. Scale bars=5 µm.
| Karyotype form | Ploidy | No. of chromosomes | Karyotype formula | No. of satellites | Range of arm ratio | Range of length (µm) | Total length (µm) | Longest chromosome length / shortest chromosome length | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| M | m | sm | Total | ||||||||
| SH1 | 3x | 0 | 21 | 3 | 24 | 2n=24=21m+3smcs | 3 | 1.1–1.9 | 1.7–2.9 | 54.2 | 1.71 |
| SH2 | 3x | 1 | 20 | 3 | 24 | 2n=24=19(M+m)+1mcs+1mcscs+3sm | 3 | 1.0–2.0 | 1.7–3.7 | 54.4 | 2.18 |
Metaphase chromosomes were 1.7–2.9 µm in length and 1.1–1.9 in arm ratio, with the ratio of the longest chromosome to the shortest chromosome being 1.71 (Table S1). Of the 24 chromosomes, satellites were found on the long arms of the three longest submetacentric chromosomes (Fig. 1A, C). The karyotype formula was expressed as 2n=24=21m+3smcs. This karyotype was determined to be identical to ‘Karyotype 1’ reported by Sato et al. (2014) (i.e., the longest three chromosomes with satellites on the long arms).
Karyotype SH2Metaphase chromosomes were 1.7–3.7 µm in length and 1.0–2.0 in arm ratio, with the ratio of the longest chromosome to the shortest chromosome being 2.18 (Table S2). Among the 24 chromosomes, satellites were found on the long arm of one chromosome and on both arms of one chromosome (Fig. 1B, D). This karyotype, with the formula 2n=24=19(M+m)+1mcs+1mcscs+3sm, is novel in this species.
Since the number of satellites per cell is three, which is the same as that in other triploid individuals including karyotype SH1 plants, the new karyotype SH2 is considered to have resulted from a somatic reciprocal translocation between the arms of chromosomes with satellites. The new karyotype identified in the present study strongly suggests that chromosomal differentiation is caused by somatic recombination. Although many apomicts are of hybrid origin and have arisen as allopolyploids from sexual parents, this unique karyotype is unlikely to have arisen from hybridization between different races. Chromosome inversion or translocation has been suggested to play an important role in the diversification of some agamospermous Taraxacum, since tetraploid T. venustum, the congener, has four strains distinguished by their karyotypes (Sato et al. 2020). The new karyotype detected in this study likewise suggests that chromosomal inversion or translocation contributed to cytological diversification.
Solbrig (1971) demonstrated that populations of common dandelions consisted of mixtures of genotypes that differ in their ability to compete, as well as in their strategies of survival, by two sets of experiments. According to Stebbins (1950), causes of variation in agamic complexes are, first, hybridization and allopolyploidy between the original sexual ancestors of the agamic complex; second, hybridization between facultative apomicts, with the resulting segregation, or between apomicts and sexual species; and third, chromosomal and genic changes within apomictic clones themselves. Richards (1989) reported remarkably high levels of chromosome breakage and somatic recombination within some Taraxacum taxa, with marked variation between lines or even within an agamospecies. The karyotypic variation found in some Japanese agamospermous Taraxacum is consistent with this. Such a line can diversify and evolve without sexual intervention.
The findings of the present study indicate that karyotypic diversities of apomictic Taraxacum are increased by the occurrence of chromosome mutations, such as structural changes in chromosomes. Cytological data clearly prove that the mechanisms of the karyotype evolution of Taraxacum vary between the usual sexual diploids and apomictic polyploids.
This research was supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number JP21K05633. Our special thanks go to Mr. T. Yamazaki for providing us with the plant materials.
K. S. designed the study and performed the experiments. K. S. and Y. I. approved the final version of the manuscript.
Supplementary information including Tables S1 and S2 is available online.