Abstract
Evidence has been accumulating that environmental chemicals contained in foods can cause
neurodevelopmental disabilities. However, regular developmental neurotoxicity (DNT)
studies require large numbers of animals for detection of subtle dose-response changes,
and it is urgent concern to establish a rapid and efficient evaluation system of DNT.
Evidence from our recent studies points to the notion that adult neurogenesis in the
hippocampus may represent vulnerable endpoint to cause DNT. Adult neurogenesis is the
postnatal process of continued production of new neurons through the adult stage in the
brain. Monitoring of granule cell lineage in the subgranular zone (SGZ) and γ-aminobutyric
acid (GABA)-ergic interneurons in the dentate hilus is effective for detection of target
cell populations of DNT as manifested by disruption of neurogenesis. Especially,
reelin-expressing GABAergic interneurons are a useful marker to detect disruption of the
migration and correct positioning of developing neurons following impairment of
neurogenesis. Because axon terminal toxicants target granule cell lineage population
growing dendritic processes, there may be common target mechanisms between the DNT and
adult-type neurotoxicity affecting mature nervous system. Adult neurogenesis may also be a
suitable endpoint for detection of DNT in a scheme of standard regular 28-day toxicity
study. In other words, adult-type neurotoxicity could be detected by measuring the
cellular responses in adult neurogenesis. Moreover, it should be stressed that there may
be epigenome toxicity mechanisms to affect the process of hippocampal neurogenesis
involving both neuronal stem cells and interneuron subpopulations, with continued
disruption through the adult stage. These findings suggest that hippocampal neurogenesis
is considered to be a critical target of environmental neurotoxicants contained in
foods.
1. Introduction
It is reported that around 13 percent of children were reported to have ever had a
developmental disability and in most cases these disabilities affect the nervous
system1). Such
neurodevelopmental disorders involving inherent neural dysfunctions include pervasive
developmental disorder (autism spectrum disorder), learning disability, cerebral palsy,
seizures, attention-deficit hyperactivity disorder (ADHD) and mental retardation. Their
causes are mostly unknown. Among them, there is an increasing trend in prevalence of ADHD
and learning disability in a survey conducted in the United States1). The disabilities in these disorders are permanent
in nature, and there are no effective therapeutic choices; they become a heavy burden to
families and to society.
There is a growing concern on infantile health effects by exposure to environmental
contaminants through ingestion of foods, and developmental neurotoxicity (DNT) by ingested
chemicals is one of the urgent global concerns because of lifelong disability.
Neurodevelopmental disabilities caused by environmental chemicals are mostly subclinical
ones2). Environmental
chemicals, such as lead, methylmercury, arsenicals, polychlorinated biphenyls (PCBs), and
toluene, can cause developmental neurotoxicities in man2). An expert committee from the US National Research Council
concluded that around 3% of developmental neurotoxicities are considered to be the direct
effect due to exposure to such environmental chemicals, and another 25% are regarded to be
the outcome of interactions between individual genetic susceptibility and environmental
factors3). These estimates
were based on scarce information about neurotoxicity and could therefore underestimate the
true prevalence of chemically-induced abnormalities2).
The developing human brain is inherently much more susceptible to injury caused by toxic
agents than is the brain of an adult4). If a developmental process of the brain is inhibited or
disrupted, there is little potential for later repair, and the consequences can therefore be
permanent5). Generally,
during embryonic and fetal period, organogenesis stage is most sensitive for induction of
serious morphological defects against exposure to exogenous agents. On the other hand, brain
development continues even after organogenesis period, and a considerable amount of cellular
and physiological events occur during postnatal development, such as neurite outgrowth,
neuronal guidance, synaptogenesis, myelination and maturation in the central nervous
system6). Of note, the human
brain continues to develop postnatally, and the period of heightened vulnerability therefore
extends over many months, through infancy and into early childhood. Although most neurons
are formed by the time of birth, growth of glial cells and myelination of axons continues
for several years5,7). It is also noted that
reorganization of synaptic plasticity of neuronal networks in the brain continues during
postnatal period in children8).
Therefore, all of these neurodevelopmental events are suggested to be vulnerable endpoints
to cause DNT.
During development, neonates and infants have particularity in the variety and content of
foods daily ingested as well as in the behavior under the living environment, such as
crawling and hand-to-mouth behavior at the living floor9). Therefore, these immature populations have inherent
exposure risk to environmental contaminants different from those of adult populations. For
example, infants have limited variety in foods, and they are fed initially and solely breast
milk and then baby foods made largely of fruits and dairy products. Breast milk may be
contaminated with persistent lipophilic chemicals, including specific pesticides and
halogenated industrial compounds, and limited variety of daily foods may increase the
exposure risk to chemicals contaminated in particular foods. Persistent lipophilic
chemicals, such as PCBs, accumulate in maternal adipose tissue and are passed on to the
infant via breast milk, resulting in infant exposure that exceeds the mother’s own exposure
by 100-fold on the basis of body weight10). The susceptibility of infants and children to chemicals is
further enhanced by their increased exposures and absorption rates, and diminished ability
to detoxify many exogenous compounds, relative to that of adults11). Moreover, infants intake higher amount of foods
per body weight basis reflecting higher body burden on exposed chemicals than adults.
There are hundreds of chemicals that have been proved to cause neurotoxic insults in
adults2). There also are
many other chemicals that have been shown to be neurotoxic experimentally. However, the
neurotoxic effects of such chemicals in the developing humans are largely unknown and they
are not subjected to regulatory control to protect from exposure of humans including
children. There are two major impediments to prevention of DNT by chemical exposure. One is
the lack of available mechanism-based models in testing chemicals for DNT and another being
the high level of proof required for regulation. Novel and precision approaches that capture
the unique vulnerability of the developing brain are necessary for testing and control of
chemicals.
The hippocampus is belonging to the limbic system and plays essential roles in the memory
formation for short and long term, as well as consolidation of information, and spatial
navigation. The hippocampal formation comprises a group of cortical tissues including the
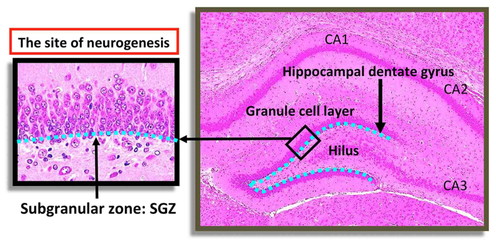
cornu ammonis and dentate gyrus (Fig. 1). The
dentate gyrus is the primary input into the hippocampal formation receiving connections from
the entorhinal cortex. The subgranular zone (SGZ) of the dentate gyrus, a subregion of the
hippocampus located at the base of granule cell layer (GCL), uniquely continues to generate
new neurons during postnatal life12). This process is called as “adult neurogenesis”. Adult
neurogenesis in the dentate gyrus is critical for some forms of learning and memory and is
modulated by pathological conditions13). and some neurological and psychiatric disorders are
associated with malformation and dysfunction of the dentate gyrus14). Of note, hippocampal neurogenesis is prominent in
adult rodents as well as primates and humans15,16,17,18).
Current DNT studies require large numbers of animals for detection of subtle dose-response
changes. For screening purposes of many new chemicals, smaller scale studies, preferably
with short-term experiments, employing suitable and sensitive neurodevelopmental endpoints
focusing on histopathological parameters need to be established. To establish a rapid
screening system for developmental neurotoxicants, it is reasonable to focus on cellular
response in the hippocampal adult neurogenesis and its regulatory system using rodent
models19,20,21,22,23,24,25,26). Because adult neurogenesis includes all processes from
neuron production to their maturation, it can be hypothesized that monitoring of the dentate
gyrus may provide a valuable tool for the detection of developmental neurotoxicants. The aim
of this review is to overview our recent study findings regarding the availability of
hippocampal neurogenesis to monitor as a critical target of environmental neurotoxicants
contained in foods. At first, this review provides the information regarding the
availability of monitoring the γ-aminobutyric acid (GABA)-ergic interneurons distributed in
the hilus of the dentate gyrus for detection of chemicals that can affect neurogenesis and
migration using a rat model of developmental hypothyroidism. The next topic is regarding the
presence of vulnerable targets of DNTs in the process of neurogenesis by neurotoxicants that
can injure mature nerve tissue using acrylamide and glycidol as examples in rats. The third
topic is regarding the availability of adult neurogenesis for detection of DNT in the
framework of 28-day regular toxicity study using rats. As a last topic, an example of
epigenome toxicity in the process of neurogenesis is provided in a mouse model of manganese
(Mn)-induced neurogenesis toxicity.
2. Adult Neurogenesis in the Hippocampus
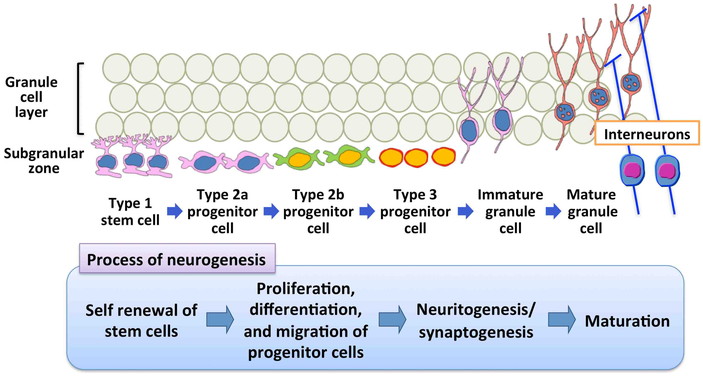
In mammals, adult neurogenesis consists of multiple processes, including a number of
developmental phases such as the self-renewal of stem cells, facilitation of cell division
of precursor cells to produce newborn granule cells, and the subsequent differentiation and
migration of new granule cells to the correct position within the GCL (Figs. 2 and 3)12). Stem cells (type-1 cells) exist
in the SGZ of the dentate gyrus and divide slowly to produce intermediate progenitor cells,
a type of transient amplifying cells27,28,29). Undifferentiated intermediate progenitor cells (type-2a and
type-2b) divide rapidly to produce neuronally committed intermediate progenitor cells
(type-3) and react to stimuli that influence neuronal generation in the hippocampal dentate
gyrus28). Type-3
intermediate progenitor cells produce postmitotic immature neurons, and those neurons that
come through regulatory period integrate into the GCL as granule cells27). By monitoring of cellular
subpopulations in the SGZ and GCL as well as cell proliferation activity and apoptosis in
the SGZ, it is possible to detect changes in entire processes of the granule cell lineages
(Fig. 4). For this purpose, immunohistochemical
analysis could be a powerful tool for detection of target cells using primary antibodies
against proliferating cell nuclear antigen (PCNA), a cell proliferation marker, brain lipid
binding protein (BLBP), expressed in radial glia as type-1 cells in the brain30), paired box 6 (Pax6), expressed in
type-1 stem cells and type-2a progenitor cells31), SRY (sex determining region Y)-box 2 (Sox2), expressed in
type-1 stem cells and type-2a and type-2b progenitor cells31), T box brain 2 (Tbr2), expressed in type-2b
progenitor cells31),
dihydropyrimidinase-like 3 (Dpysl3), also known as Tuc4, an early postmitotic cell marker of
immature granule cells32),
NeuroD1 (Nd1), expressed in type-3 progenitor cells and immature granule cells33), and doublecortin (Dcx), expressed
in type-3 progenitor cells and immature granule cells27), as well as measurement of apoptosis utilizing terminal
deoxynucleotidyl transferase-mediated dUTP-biotin nick end-labeling assay.
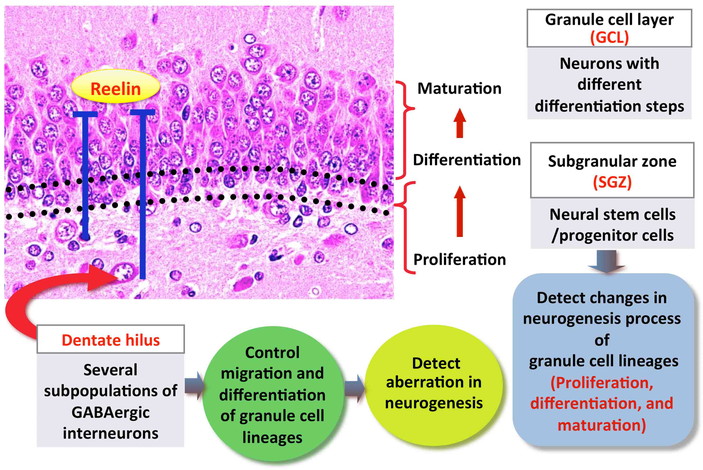

In the hilar region of the dentate gyrus, GABAergic interneurons are known to make synaptic
connections with adult-born dentate granule cells and play an important role in regulating
adult neurogenesis (Figs. 2 and 3)34). In particular, subpopulation GABAergic interneurons secrete
reelin, an extracellular matrix glycoprotein that modulates progenitor cell migration to
maintain normal integration of granule cells in the neonatal and adult dentate
gyrus35). Moreover, there
are unique populations of GABAergic interneurons expressing calcium-binding proteins, such
as calbindin D-28k (Calb1), parvalbumin (Pvalb), and calbindin D-29k (Calb2, also known as
calretinin), that form a family of proteins containing the EF-hand Ca-binding motif, and
potentially serve critical roles in the development and functioning of the brain36). It is known that interneurons
innervate progenitor cells in the SGZ resulting in promotion of progenitor cell
differentiation36,37). Because the number of GABAergic interneurons in the dentate
hilus changes in relation with the disruption of neurogenesis, immunohistochemical
approaches for analysis of these cellular populations also provide valuable information with
regard to the type and severity of aberration in neurogenesis (Fig. 4).
Various neurons outside the hippocampus also make synaptic connection with neurons in the
dentate gyrus to function on progenitor cell proliferation and differentiation. For example,
cholinergic neurons originated from the septal nucleus and nucleus of the diagonal band of
Broca innervates neurons in the dentate hilus. Noradrenergic neurons in the locus ceruleus
innervate neurons in the SGZ38). Several studies indicated that acetylcholine can act directly
on neuronal progenitors in the adult brain39,40), and cholinergic denervation is reported to cause cell death
and decrease the numbers of new neurons in the hippocampal SGZ41). Therefore, dysfunction of cholinergic inputs,
such as that evidenced by the presence of histopathological changes in the hippocampal
fimbria may directly induce the reduction of intermediate progenitor cells in the
hippocampal dentate gyrus. Of note, synaptic connections from the outside the hippocampus
are seen with SGZ/GCL cells and/or GABAergic interneurons42). By monitoring neuronal inputs from the outside of
the dentate gyrus or their postsynaptic receptors distributed within the dentate gyrus, it
is also possible to detect aberrations in adult neurogenesis. For this purpose,
immunohistochemical analysis and/or real-time reverse transcription-polymerase chain
reaction technique with regard to related molecules, such as ligands, receptors and
transporters, may be applied, as well as histopathological analysis of nerve tract
changes.
3. Effect of Hypothyroidism and Thyroid Hormone-disrupting Chemicals on
Neurogenesis
Thyroid hormones (THs) play essential roles for normal brain development in mammals. It is
well known that THs regulate neuronal proliferation, migration, and differentiation in
discrete brain regions during development43). In an experimental model of developmental hypothyroidism,
exposure to anti-thyroid agents (ATAs) leads to growth retardation, neurological
abnormalities and impaired performance in a variety of behavioral learning actions44,45). Rat offspring exposed maternally to ATA show
impaired brain growth involving neuronal mismigration as well as white matter
hypoplasia46), due to
limited axonal myelination and reduction in oligodendroglial population47,48). The outcome of this type of neural deficit is
permanent accompanied by apparent structural and functional abnormalities.
There are environmental chemicals that are thought to have a TH-disrupting potential to
cause abnormal brain development49). Particularly, brominated flame retardants (BFRs), some of
which are environmental contaminants used in electronic circuitry, plastics, textiles and
other materials to prevent fires, are known to be weak TH-disruptors. Therefore, there is a
growing concern with regard to the DNT by BFRs50,51). Decabromodiphenyl ether (DBDE), tetrabromobisphenol A
(TBBPA) and 1,2,5,6,9,10-hexabromocyclododecane (HBCD) are three major and widely used
BFRs52). In experimental
studies using developmental exposure models in rats and mice, these BFRs have been reported
to cause neurobehavioral and auditory response effects51,53,54,55,56,57,58). These BFRs have shown weak anti-thyroid activities in some
studies51,58,59,60,61). In contrast, there are experimental studies suggestive of
direct effect of these BFRs on brain development62,63,64,65).
In a study to detect cellular evidence of the disruption of hippocampal neurogenesis due to
developmental hypothyroidism19), we performed immunohistochemical analysis in the hippocampal
dentate gyrus in rat offspring after developmental exposure to ATAs, 6-propyl-2-thiouracil
and methimazole (MMI). Effects on neurogenesis were examined in terms of the detection of
target cells in the granule cell lineages and interneuron subpopulations. For this purpose,
distribution of reelin-producing interneurons was analyzed in the hippocampal dentate hilus.
Obtained results indicate persistent increases in GABAergic interneurons producing reelin in
an immature population through the adult stage in the dentate hilus after developmental
hypothyroidism (Fig. 5). Interestingly, we
observed sustained increases of immature GABAergic interneurons synthesizing reelin in the
hilus, and this phenomenon was considered to be a signature of compensatory regulation for
persistent impairment of neurogenesis and following migration during the neuronal
development as a hypothyroidism-related brain effect.

In a study to investigate the impact and reversibility of developmental exposure to BFRs,
DBDE, TBBPA or HBCD, we immunohistochemically examined reelin-producing interneuron
population and cell proliferation and apoptosis in the dentate SGZ using rats66). Obtained results indicate that
all the BFRs examined had developmental exposure effect on neurogenesis (Fig. 5). Our results suggested that a direct effect of
DBDE and TBBPA on hippocampal neurogenesis. However, effect of hypothyroidism may also be
operated at least at the highest dose of DBDE as well as that of HBCD. Because of the
reversibility at the adult stage, it was considered that the effect of these BFRs on
neurogenesis and following neuronal migration to the GCL occur during the exposure period.
We also observed increase in neuron-specific nuclear protein (NeuN)+ mature
neuronal populations at the adult stages; however, the its biological significance should be
further assessed.
There are many environmental chemicals having a potential to interfere with TH signaling in
the developing brain. The above-mentioned study results of ATAs and BFRs suggest that
reelin-expressing GABAergic interneurons in the dentate hilus may be a useful marker for
assessment of the effects of developmental neurotoxicants that can disrupt the migration and
positioning of developing neurons following disruption of neurogenesis.
4. Possibility of DNT by Adult-type Neurotoxicants
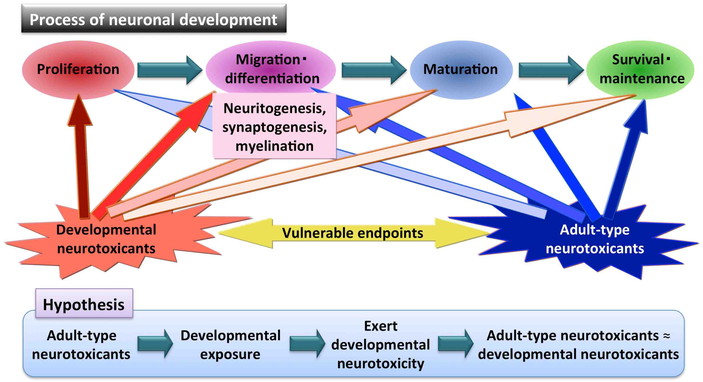
In the hippocampal dentate gyrus, all of the cell populations and their inherent phenomena
involved in the process of adult neurogenesis may be sensitive target of DNT. Especially,
self-renewal of stem cells, proliferation and migration of progenitor cells, neuritogenesis,
synaptogenesis and myelinogenesis may be the vulnerable developmental processes against
chemical toxicity. Because the molecular mechanisms to control neuronal development
processes have many similarities with those described for mechanisms for neuronal
maintenance after maturation, neurogenesis process may be vulnerable to “adult-type
neurotoxicants” to show affection of mature nervous tissue (Fig. 6). Of note, among adult-type neurotoxicants, myelin toxicants
may affect myelination of interneuron populations and neuronal inputs from outside the SGZ,
and granule cells may not directly be affected because they are consisted of non-myelinated
fibers.
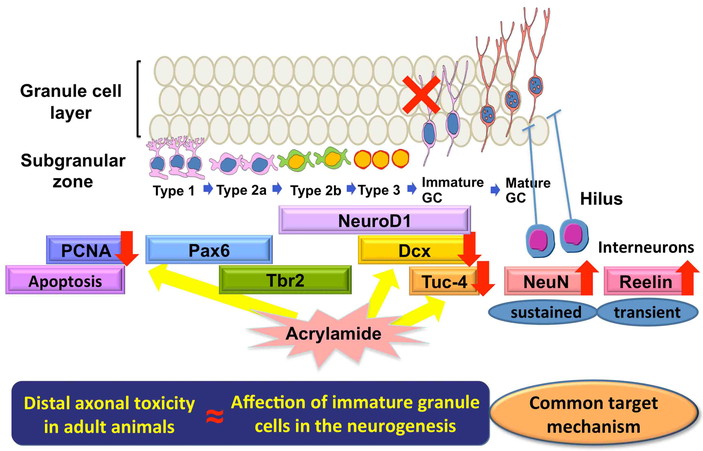
In this review, DNT of acrylamide in the rat hippocampal neurogenesis is presented, and
similarity in the target mechanisms between developmental and adult-stage neurotoxicity is
discussed. Acrylamide, a widely used chemical in many industries, is known to be a neuronal
and reproductive toxicant, and to act as a carcinogen in animals67). Because it was recently found that acrylamide is
generated during heating of foods containing carbohydrate and asparagine, risk assessment
studies of acrylamide in foodstuffs have been conducted globally68,69). Acrylamide is a well-known axon terminal toxicant in both
the central and peripheral nervous systems in adult animals70).
Axon guidance during development and after axonal injury is an important process in
maintaining neuronal plasticity at the axon terminals71). It is well established that the molecular mechanisms
controlling neuronal migration during development have many similarities with those
necessary for axon guidance72).
Therefore, both migrating neuronal progenitor cells and immature axon terminals may have
sensitivity to acrylamide.
In a study of maternal exposure study of acrylamide using rats20), we found that acrylamide impairs hippocampal
neurogenesis in rat offspring as evident by the increase in GABAergic interneurons producing
reelin in the dentate hilus (Fig. 7). We then
examined the cellular target of acrylamide on hippocampal neurogenesis and its reversibility
after maternal exposure using the similar developmental exposure model21), and found reversible affection of
neurogenesis targeting the proliferation of type-3 progenitor cells resulting in a reduction
of immature granule cells (Fig. 7). In the SGZ, it
is known that immature granule cells already have both dendritic growth cones and recurrent
basal dendrites, suggesting an entry into the process of synaptogenesis73). Because acrylamide targets nerve
terminals due to impairment of neurotransmission by affecting diverse nerve terminal
processes74), it was
suggested that acrylamide directly injure immature granule cells by affecting the newly
generating nerve terminals. Importantly, we also found that acrylamide suppressed progenitor
cell proliferation without accompanying facilitation of their apoptosis. Because immature
granule cells can no longer proliferate, acrylamide may rather target earlier type-3
progenitor cells to suppress their proliferation causing a reduction in immature granule
cells.
We found another case of axon terminal toxicant to show similar target cell population at
the late-stage differentiation in the hippocampal neurogenesis after developmental exposure.
In a developmental exposure study of glycidol25), we found induction of distal axonopathy in dams and
reduction of immature granule cells in the SGZ of offspring (Fig. 8). Glycidol is a carcinogen that has been classified by the
International Agency for Research on Cancer (IARC) as a group 2A carcinogen, which is
“probably carcinogenic to humans”75). Exposure to glycidol through food has recently become a
worldwide concern based on possible release of glycidol by hydrolysis of glycidol fatty acid
esters in the gastrointestinal tract. Glycidol fatty acid esters can be found in refined
edible oils and fats, including infant formulas, especially in diacylglycerol oil at high
concentrations76,77). In a repeated-oral-dose toxicity
study for 13 weeks in rats and mice, neurotoxicity involving cerebellar necrosis was
reported78). Our study
results in the developmental exposure study of glycidol suggest that glycidol targets the
newly generating nerve terminals of immature granule cells, resulting in the suppression of
late-stage hippocampal neurogenesis, similar to acrylamide (Fig. 8).
Our study results of developmental exposure to acrylamide or glycidol suggest that there
may be common target mechanisms between the DNT and adult-type neurotoxicity (Fig. 6).
5. Detection of DNT in a Framework of 28-day Regular Toxicity Study
Because postnatal adult neurogenesis continues through the adult stage, it is reasonable to
capture the toxicity in the process of neurogenesis in a framework of regular toxicity
studies, such as 28-day repeated oral dose toxicity study. To clarify this possibility, we
compared the effects on neurogenesis in the DNT study and repeated oral dose toxicity study
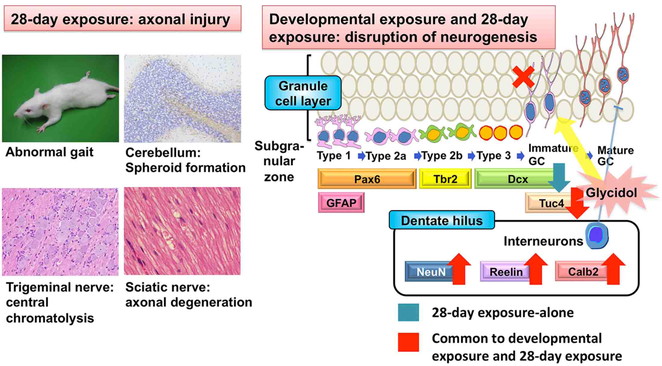
using MMI as an ATA and glycidol as an axon terminal toxicant25,79,80).
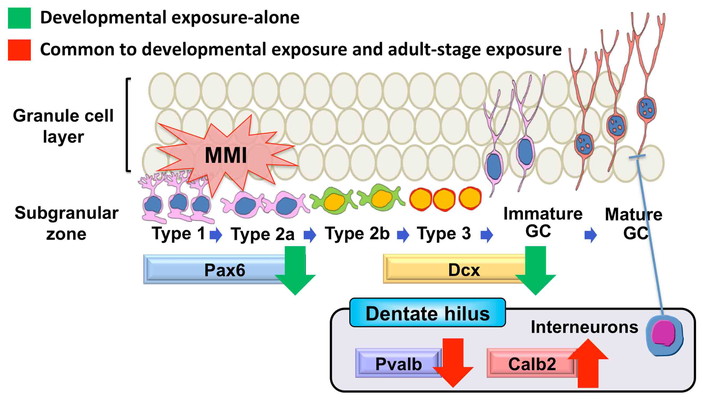
In the study of MMI, pregnant rats were exposed to MMI from gestational day (GD) 10 until
weaning on day 21 after delivery (developmental hypothyroidism)79). Adult male rats were also exposed to MMI by
setting an identical exposure period from postnatal day (PND) 46 through to PND 77
(adult-stage hypothyroidism). As a result, a sustained reduction of Pax6+ stem or
early progenitor cells and a transient reduction of Dcx+ late-stage progenitor
cells were observed after developmental hypothyroidism (Fig. 9). These cells were unchanged by adult-stage hypothyroidism. On the other
hand, the number of Pvalb+ GABAergic was reduced and the number of
Calb2+ interneurons was increased in the dentate hilus after both developmental
and adult-stage hypothyroidism with MMI (Fig. 9).
Our results indicate that fluctuations in GABAergic interneuron subpopulations are the
common feature in both developmental and adult-stage hypothyroidism. Considering their role
in neurogenesis, fluctuations in GABAergic interneuron subpopulations may provide a
sensitive tool for detection of aberrant neurogenesis even after the adult-stage exposure.
Our study also suggests that analysis of interneuron subpopulations may provide a tool for
detection of subtle changes in neurogenesis, particularly for regular toxicity tests such as
the 28-day repeated-oral dose-toxicity test.
In a study of glycidol, glycidol was exposed to pregnant rats from GD 6 until weaning on
day 21 after delivery (developmental exposure study) and male young adult rats by gavage for
28 days (adult-stage exposure study)25,80). In the developmental exposure study, offspring reduced
Dpysl3+ immature granule cells in the SGZ and increased immature
reelin+ or Calb2+ GABAergic interneurons and NeuN+ mature
neurons in the dentate hilus on weaning (Fig. 8).
Hilar changes remained until adult stage, with the increases in reelin+
interneurons and NeuN+ mature neurons being present, although the SGZ change
disappeared. In the adult-stage exposure study, animals revealed aberrations in neurogenesis
at the late-stage differentiation as evidenced by reductions of both Dcx+ and
Dpysl 3+ cells in the SGZ and increases of reelin+ and
Calb2+ GABAergic interneurons and NeuN+ mature neurons in the
dentate hilus (Fig. 8). These results suggest that
glycidol targets the newly generating nerve terminals of immature granule cells, resulting
in the suppression of late-stage hippocampal neurogenesis in both developmental and
adult-stage exposure studies.
These results suggest that adult neurogenesis in the SGZ may provide a suitable endpoint
for detection of DNT in a standard regular 28-day toxicity study. In other words, both
developmental and adult-type neurotoxicity may be detected by analysis of the cellular
responses in the adult neurogenesis using adult animals. From this point of view, a rapid
screening system for developmental neurotoxicants may be constructed by measurement of the
effect on adult neurogenesis in a framework of 28-day toxicity study. After initial
screening in the 28-day toxicity study, DNT may be definitively detected in a framework of
DNT studies by incorporating the analysis of neurogenesis-related parameters like those
presented here in the histopathological endpoint, in combination with functional and
behavioral observation battery. It is necessary to collect information on adult-type
neurotoxicants regarding relationship between the toxicity target on adult nervous system
and that on neurogenesis.
6. Epigenome Toxicity in the Process of Hippocampal Neurogenesis
Recent studies indicate that various epigenetic mechanisms, including DNA methylation of
gene promoter region, histone deacetylation and microRNAs, are involved in regulating
multiple processes of adult mammalian neurogenesis81), and environmental factors can influence this
regulation82). The
understanding of epigenetic gene regulation as a long-lasting cellular memory necessary to
maintain or evolve a cellular phenotype has recently been challenged by discoveries of its
dynamic nature82). In general,
gene expression is related inversely to DNA methylation83). This relationship is particularly evident in CpG islands
(CGIs) at gene promoter regions, where DNA methylation may directly interfere with
transcription factor binding to DNA and/or indirectly suppress transcription through
methylated DNA binding proteins that recruit histone deacetylases, leading to chromatin
condensation and subsequent gene silencing84). Although the results of epigenetic regulation changes on
neurogenesis have remained unexplored, environmentally induced disruption of DNA methylation
clearly deserves further study85) given the clear importance of DNA methylation to processes of
neuronal development.
Mn is a trace essential element important for protein and energy metabolism, bone
mineralization, metabolic regulation, and cellular protection from reactive oxygen
species86). Excess Mn
exposure causes serious neurotoxicity, such as manganism, which shares multiple features
with Parkinson’s disease87). We
have recently found that maternal developmental Mn exposure in mice affects neurogenesis
targeting immature granule cells in the SGZ of offspring even at the adult stage,
accompanied with a sustained increase in the immature population of reelin-synthesizing
GABAergic interneurons88).
These changes may represent continued aberrations in neurogenesis and subsequent migration
to cause an excessive response to overproduce immature granule cells through the adult
stage. To clarify the effects of maternal Mn exposure on epigenetic gene regulation
contributing to this sustained disruption, we searched epigenetically downregulated genes by
global promoter methylation analysis in the hippocampal dentate gyrus of Mn-exposed
offspring at the end of maternal exposure on weaning and further examined to confirm
methylation status as well as transcript expression levels of candidate genes on both
weaning and adult stage89).
By global promoter methylation analysis using CpG promoter microarrays, we have found that
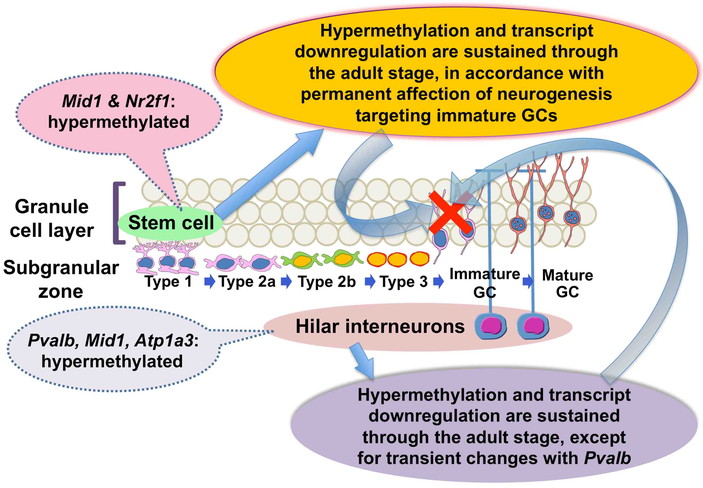
maternal Mn exposure induced CGI hypermethylation of the promoter region and transcript
downregulation of Pvalb, Mid1 (midline 1),
Atp1a3 (ATPase, Na+/K+ transporting, alpha 3
polypeptide) and Nr2f1 (nuclear receptor subfamily 2, group F, member 1;
also known as COUP-TF1) in the hippocampal dentate gyrus of offspring89). Sustained promoter
hypermethylation and transcript downregulation through adult stage were confirmed with
Mid1, Atp1a3 and Nr2f1, whereas
Pvalb showed a transient hypermethylation only on weaning (Fig. 10). By immunohistochemical analysis, we also
observed reductions of Pvalb+ and ATP1a3+ neurons suggestive of
GABAergic interneurons in the hilus, Mid1+ cells suggestive of late-stage
progenitors or postmitotic granule cells in the SGZ and interneurons in the hilus and
COUP-TF1+ cells suggestive of stem cells and early-stage progenitors in the
SGZ. All of these reductions except for those of Pvalb+ cells were sustained
through the adult stage. These observations suggested continued disruption of neurogenesis
and subsequent neuronal differentiation by Mn exposure.
Evidence suggests that Pvalb+ cells, a subpopulation of GABAergic interneurons,
play a central role in the inhibition of granule cell activity90). With regard to ATP1a3, an ion pump component that
maintains sodium and potassium gradients across the plasma membrane, a subpopulation of
GABAergic interneurons in the hippocampus were reported to express this protein91). Because GABAergic interneurons
provide direct connection to type-2 progenitor cells in the SGZ to promote progenitor cell
differentiation37),
reduction of Pvalb+ cells or ATP1a3+ cells may lead to inhibition of
neuronal differentiation after Mn exposure.
Mid1 is a gene encoding a ubiquitin ligase92) and affects brain development by regulating
expression of Gli393), a Wnt target gene involved in hippocampal
development94). Loss of
Mid1 function by mutation is involved in X-linked Opitz G/BBB syndrome
characterized by defective midline development95). Therefore, promoter hypermethylation and subsequent
Mid1 downregulation may be linked to Mn-induced disruption of
neurogenesis. Interestingly, it is reported that Mid1 is expressed at the right side of
Hensen’s node of chick embryos to play a key role in gene cascade downregulation during the
early stages of left-right determination through sonic hedgehog pathway inhibition96). We have also found right-side
predominance of Mid1+ cells and its abolishment by Mn exposure by promoter
hypermethylation of this side, suggestive of the disruption of higher brain functions
specialized in the left or right side of the brain89).
COUP-TF1 is one of the most characterized orphan receptors of the steroid/thyroid hormone
receptor superfamily and is an important regulatory component of neurogenesis and following
differentiation during development97). COUP-TF1 is expressed in both neuronal progenitors and
postmitotic neurons98).
Therefore, our result suggests a permanent effect on epigenetic gene control of
Nr2f1 involving stem cells by developmental Mn exposure to trigger
disruption of neurogenesis and subsequent neuronal differentiation because epigenetic gene
changes can be inherited to daughter cells.
These findings point to epigenetic mechanisms as mediators, through which developmentally
administered neurotoxicants modulate postnatal adult neurogenesis with long-lasting and
stable repercussions.
7. Conclusion
Monitoring of granule cell lineage in the SGZ and GABAergic interneurons in the dentate
hilus is effective for detection of target cell populations of DNT in the process of adult
neurogenesis. Especially, monitoring of reelin-expressing GABAergic interneurons is useful
for detection of neuronal mismigration following impairment of neurogenesis. Because axon
terminal toxicants target granule cell lineage population growing dendritic processes, there
may be common target mechanisms between the DNT and adult-type neurotoxicity. Adult
neurogenesis may also serve as a suitable endpoint for detection of DNT in a scheme of
standard regular 28-day toxicity study. In other words, it is suggested that both
developmental and adult-type neurotoxicity could be predicted by the cellular responses in
the adult neurogenesis. Moreover, epigenome toxicity mechanisms may be operated in the
process of hippocampal neurogenesis involving both neuronal stem cells and interneuron
subpopulations, with continued disruption through the adult stage. These findings suggest
that hippocampal neurogenesis is considered to be a critical target of environmental
neurotoxicants contained in foods. Considering pluripotent nature of neural stem cells in
the SGZ to differentiate to both neuronal and glial cell lineages, further studies are
necessary on the late-stage onset of stem cell toxicity in the neurogenesis.
Conflict of Interest Statement
The authors declare that no conflicts of interest exist.
Acknowledgments
Our studies covered by this review were supported in part by Health and Labour Sciences
Research Grants (Research on Risk of Chemical Substances) from the Ministry of Health,
Labour and Welfare of Japan, by a Grant-in-Aid for Scientific Research (B) from the Ministry
of Education, Culture, Sports, Science and Technology of Japan (Grant No. 25292170) or by a
grant from the Ministry of Economy, Trade and Industry (METI), Japan.
References
- 1. Boyle CA, Boulet S, Schieve LA, et al. Trends in
the prevalence of developmental disabilities in US children, 1997-2008. Pediatrics. 2011;
127(6): 1034–1042.
- 2. Grandjean P, Landrigan PJ. Developmental
neurotoxicity of industrial chemicals. Lancet. 2006; 368(9553): 2167–2178.
- 3. National Research Council Scientific frontiers in
developmental toxicology and risk assessment. Washington, DC: National Academy Press;
2000.
- 4. Dobbing J. Vulnerable periods in developing
brain. In Applied Neurochemistry (A.N. Davison, J. Dobbing, eds.). Oxford: Blackwell,
Scientific Publications; 1968; 287–316.
- 5. Rice D, Barone S, Jr Critical periods of
vulnerability for the developing nervous system: evidence from humans and animal models.
Environ Health Perspect. 2000; 108 (Suppl 3): 511–533.
- 6. Tschopp O, Yang ZZ, Brodbeck D, et al. Essential
role of protein kinase B gamma (PKB gamma/Akt3) in postnatal brain development but not in
glucose homeostasis. Development. 2005; 132(13): 2943–2954.
- 7. Rodier PM. Developing brain as a target of
toxicity. Environ Health Perspect. 1995; 103 (Suppl 6): 73–76.
- 8. Johnston MV. Plasticity in the developing brain:
implications for rehabilitation. Dev Disabil Res Rev. 2009; 15(2): 94–101.
- 9. Xue J, Zartarian V, Moya J, et al. A
meta-analysis of children’s hand-to-mouth frequency data for estimating nondietary
ingestion exposure. Risk Anal. 2007; 27(2): 411–420.
- 10. Jensen AA, Slorach SA. Factors affecting the
levels of residues in human milk. In Chemical contaminants in human milk (A.A. Jensen,
S.A. Slorach, eds.). Boca Raton, FL: CRC Press; 1991; 199–208.
- 11. Ginsberg G, Hattis D, Sonawane B. Incorporating
pharmacokinetic differences between children and adults in assessing children’s risks to
environmental toxicants. Toxicol Appl Pharmacol. 2004; 198(2): 164–183.
- 12. McDonald HY, Wojtowicz JM. Dynamics of
neurogenesis in the dentate gyrus of adult rats. Neurosci Lett. 2005; 385(1): 70–75.
- 13. Zhao C, Deng W, Gage FH. Mechanisms and
functional implications of adult neurogenesis. Cell. 2008; 132(4): 645–660.
- 14. Sato N, Hatakeyama S, Shimizu N, Hikima A, Aoki
J, Endo K. MR evaluation of the hippocampus in patients with congenital malformations of
the brain. Am J Neuroradiol. 2001; 22(2): 389–393.
- 15. Eriksson PS, Perfilieva E, Björk-Eriksson T, et
al. Neurogenesis in the adult human hippocampus. Nat Med. 1998; 4(11): 1313–1317.
- 16. Spalding KL, Bergmann O, Alkass K, et al.
Dynamics of hippocampal neurogenesis in adult humans. Cell. 2013; 153(6): 1219–1227.
- 17. Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis
in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor
proliferation. J Neurosci. 1996; 16(6): 2027–2033.
- 18. Gould E, Tanapat P, McEwen BS, Flügge G, Fuchs E.
Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is
diminished by stress. Proc Natl Acad Sci U S A. 1998; 95(6): 3168–3171.
- 19. Saegusa Y, Woo GH, Fujimoto H, et al. Sustained
production of Reelin-expressing interneurons in the hippocampal dentate hilus after
developmental exposure to anti-thyroid agents in rats. Reprod Toxicol. 2010; 29(4):
407–414.
- 20. Ogawa B, Ohishi T, Wang L, et al. Disruptive
neuronal development by acrylamide in the hippocampal dentate hilus after developmental
exposure in rats. Arch Toxicol. 2011; 85(8): 987–994.
- 21. Ogawa B, Wang L, Ohishi T, et al. Reversible
aberration of neurogenesis targeting late-stage progenitor cells in the hippocampal
dentate gyrus of rat offspring after maternal exposure to acrylamide. Arch Toxicol. 2012;
86(5): 779–790.
- 22. Ohishi T, Wang L, Akane H, et al. Transient
suppression of late-stage neuronal progenitor cell differentiation in the hippocampal
dentate gyrus of rat offspring after maternal exposure to nicotine. Arch Toxicol. 2014;
88(2): 443–454.
- 23. Ohishi T, Wang L, Akane H, et al. Reversible
effect of maternal exposure to chlorpyrifos on the intermediate granule cell progenitors
in the hippocampal dentate gyrus of rat offspring. Reprod Toxicol. 2013; 35: 125–136.
- 24. Ohishi T, Wang L, Akane H, et al. Reversible
aberration of neurogenesis affecting late-stage differentiation in the hippocampal dentate
gyrus of rat offspring after maternal exposure to manganese chloride. Reprod Toxicol.
2012; 34(3): 408–419.
- 25. Akane H, Shiraki A, Imatanaka N, et al. Glycidol
induces axonopathy by adult-stage exposure and aberration of hippocampal neurogenesis
affecting late-stage differentiation by developmental exposure in rats. Toxicol Sci. 2013;
134(1): 140–154.
- 26. Wang L, Ohishi T, Akane H, et al. Reversible
effect of developmental exposure to chlorpyrifos on late-stage neurogenesis in the
hippocampal dentate gyrus in mouse offspring. Reprod Toxicol. 2013; 38: 25–36.
- 27. Kempermann G, Jessberger S, Steiner B, Kronenberg
G. Milestones of neuronal development in the adult hippocampus. Trends Neurosci. 2004;
27(8): 447–452.
- 28. Steiner B, Kronenberg G, Jessberger S, Brandt MD,
Reuter K, Kempermann G. Differential regulation of gliogenesis in the context of adult
hippocampal neurogenesis in mice. Glia. 2004; 46(1): 41–52.
- 29. Steiner B, Klempin F, Wang L, Kott M, Kettenmann
H, Kempermann G. Type-2 cells as link between glial and neuronal lineage in adult
hippocampal neurogenesis. Glia. 2006; 54(8): 805–814.
- 30. Li H, Babiarz J, Woodbury J, Kane-Goldsmith N,
Grumet M. Spatiotemporal heterogeneity of CNS radial glial cells and their transition to
restricted precursors. Dev Biol. 2004; 271(2): 225–238.
- 31. Hodge RD, Kowalczyk TD, Wolf SA, et al.
Intermediate progenitors in adult hippocampal neurogenesis: Tbr2 expression and coordinate
regulation of neuronal output. J Neurosci. 2008; 28(14): 3707–3717.
- 32. Knoth R, Singec I, Ditter M, et al. Murine
features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 years.
PLoS One. 2010; 5(1): e8809.
- 33. Breunig JJ, Silbereis J, Vaccarino FM, Sestan N,
Rakic P. Notch regulates cell fate and dendrite morphology of newborn neurons in the
postnatal dentate gyrus. Proc Natl Acad Sci U S A. 2007; 104(51): 20558–20563.
- 34. Toni N, Laplagne DA, Zhao C, et al. Neurons born
in the adult dentate gyrus form functional synapses with target cells. Nat Neurosci. 2008;
11(8): 901–907.
- 35. Gong C, Wang TW, Huang HS, Parent JM. Reelin
regulates neuronal progenitor migration in intact and epileptic hippocampus. J Neurosci.
2007; 27(8): 1803–1811.
- 36. Freund TF, Buzsáki G. Interneurons of the
hippocampus. Hippocampus. 1996; 6(4): 347–470.
- 37. Tozuka Y, Fukuda S, Namba T, Seki T, Hisatsune T.
GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor
cells. Neuron. 2005; 47(6): 803–815.
- 38. Conrad M. Evolutionary learning circuits. J Theor
Biol. 1974; 46(1): 167–188.
- 39. Filippov V, Kronenberg G, Pivneva T, et al.
Subpopulation of nestin-expressing progenitor cells in the adult murine hippocampus shows
electrophysiological and morphological characteristics of astrocytes. Mol Cell Neurosci.
2003; 23(3): 373–382.
- 40. Nguyen L, Rigo JM, Rocher V, et al.
Neurotransmitters as early signals for central nervous system development. Cell Tissue
Res. 2001; 305(2): 187–202.
- 41. Cooper-Kuhn CM, Winkler J, Kuhn HG. Decreased
neurogenesis after cholinergic forebrain lesion in the adult rat. J Neurosci Res. 2004;
77(2): 155–165.
- 42. Itahashi M, Abe H, Tanaka T, et al. Maternal
exposure to hexachlorophene targets intermediate-stage progenitor cells of the hippocampal
neurogenesis in rat offspring via dysfunction of cholinergic inputs by myelin vacuolation.
Toxicology. 2015; 328: 123–134.
- 43. Porterfield SP. Thyroidal dysfunction and
environmental chemicals—potential impact on brain development. Environ Health Perspect.
2000; 108 (Suppl 3): 433–438.
- 44. Comer CP, Norton S. Effects of perinatal
methimazole exposure on a developmental test battery for neurobehavioral toxicity in rats.
Toxicol Appl Pharmacol. 1982; 63(1): 133–141.
- 45. Akaike M, Kato N, Ohno H, Kobayashi T.
Hyperactivity and spatial maze learning impairment of adult rats with temporary neonatal
hypothyroidism. Neurotoxicol Teratol. 1991; 13(3): 317–322.
- 46. Shibutani M, Woo G-H, Fujimoto H, et al.
Assessment of developmental effects of hypothyroidism in rats from in utero and lactation
exposure to anti-thyroid agents. Reprod Toxicol. 2009; 28(3): 297–307.
- 47. Lavado-Autric R, Ausó E, García-Velasco JV, et
al. Early maternal hypothyroxinemia alters histogenesis and cerebral cortex
cytoarchitecture of the progeny. J Clin Invest. 2003; 111(7): 1073–1082.
- 48. Schoonover CM, Seibel MM, Jolson DM, et al.
Thyroid hormone regulates oligodendrocyte accumulation in developing rat brain white
matter tracts. Endocrinology. 2004; 145(11): 5013–5020.
- 49. Bansal R, Zoeller RT. Polychlorinated biphenyls
(Aroclor 1254) do not uniformly produce agonist actions on thyroid hormone responses in
the developing rat brain. Endocrinology. 2008; 149(8): 4001–4008.
- 50. de Wit CA. An overview of brominated flame
retardants in the environment. Chemosphere. 2002; 46(5): 583–624.
- 51. Rice DC, Reeve EA, Herlihy A, Zoeller RT,
Thompson WD, Markowski VP. Developmental delays and locomotor activity in the C57BL6/J
mouse following neonatal exposure to the fully-brominated PBDE, decabromodiphenyl ether.
Neurotoxicol Teratol. 2007; 29(4): 511–520.
- 52. Birnbaum LS, Staskal DF. Brominated flame
retardants: cause for concern? Environ Health Perspect. 2004; 112(1): 9–17.
- 53. Viberg H, Fredriksson A, Jakobsson E, Orn U,
Eriksson P. Neurobehavioral derangements in adult mice receiving decabrominated diphenyl
ether (PBDE 209) during a defined period of neonatal brain development. Toxicol Sci. 2003;
76(1): 112–120.
- 54. Viberg H, Fredriksson A, Eriksson P. Changes in
spontaneous behaviour and altered response to nicotine in the adult rat, after neonatal
exposure to the brominated flame retardant, decabrominated diphenyl ether (PBDE 209).
Neurotoxicology. 2007; 28(1): 136–142.
- 55. Van der Ven LT, Van de Kuil T, Verhoef A, et al.
Endocrine effects of tetrabromobisphenol-A (TBBPA) in Wistar rats as tested in a
one-generation reproduction study and a subacute toxicity study. Toxicology. 2008;
245(1-2): 76–89.
- 56. Lilienthal H, Verwer CM, van der Ven LT, Piersma
AH, Vos JG. Exposure to tetrabromobisphenol A (TBBPA) in Wistar rats: neurobehavioral
effects in offspring from a one-generation reproduction study. Toxicology. 2008; 246(1):
45–54.
- 57. Eriksson P, Fischer C, Wallin M, Jakobsson E,
Fredriksson A. Impaired behaviour, learning and memory, in adult mice neonatally exposed
to hexabromocyclododecane (HBCDD). Environ Toxicol Pharmacol. 2006; 21(3): 317–322.
- 58. Ema M, Fujii S, Hirata-Koizumi M, Matsumoto M.
Two-generation reproductive toxicity study of the flame retardant hexabromocyclododecane
in rats. Reprod Toxicol. 2008; 25(3): 335–351.
- 59. Kitamura S, Kato T, Iida M, et al. Anti-thyroid
hormonal activity of tetrabromobisphenol A, a flame retardant, and related compounds:
Affinity to the mammalian thyroid hormone receptor, and effect on tadpole metamorphosis.
Life Sci. 2005; 76(14): 1589–1601.
- 60. Fujimoto H, Woo GH, Inoue K, et al. Impaired
oligodendroglial development by decabromodiphenyl ether in rat offspring after maternal
exposure from mid-gestation through lactation. Reprod Toxicol. 2011; 31(1): 86–94.
- 61. Saegusa Y, Fujimoto H, Woo GH, et al.
Developmental toxicity of brominated flame retardants, tetrabromobisphenol A and
1,2,5,6,9,10-hexabromocyclododecane, in rat offspring after maternal exposure from
mid-gestation through lactation. Reprod Toxicol. 2009; 28(4): 456–467.
- 62. Nakajima A, Saigusa D, Tetsu N, Yamakuni T,
Tomioka Y, Hishinuma T. Neurobehavioral effects of tetrabromobisphenol A, a brominated
flame retardant, in mice. Toxicol Lett. 2009; 189(1): 78–83.
- 63. Ibhazehiebo K, Iwasaki T, Xu M, Shimokawa N,
Koibuchi N. Brain-derived neurotrophic factor (BDNF) ameliorates the suppression of
thyroid hormone-induced granule cell neurite extension by hexabromocyclododecane (HBCD).
Neurosci Lett. 2011; 493(1-2): 1–7.
- 64. Ibhazehiebo K, Iwasaki T, Shimokawa N, Koibuchi
N. 1,2,5,6,9,10-αHexabromocyclododecane (HBCD) impairs thyroid hormone-induced dendrite
arborization of Purkinje cells and suppresses thyroid hormone receptor-mediated
transcription. Cerebellum. 2011; 10(1): 22–31.
- 65. Ibhazehiebo K, Iwasaki T, Kimura-Kuroda J,
Miyazaki W, Shimokawa N, Koibuchi N. Disruption of thyroid hormone receptor-mediated
transcription and thyroid hormone-induced Purkinje cell dendrite arborization by
polybrominated diphenyl ethers. Environ Health Perspect. 2011; 119(2): 168–175.
- 66. Saegusa Y, Fujimoto H, Woo GH, et al. Transient
aberration of neuronal development in the hippocampal dentate gyrus after developmental
exposure to brominated flame retardants in rats. Arch Toxicol. 2012; 86(9): 1431–1442.
- 67. WHO/IPCS Summary and conclusions of the
sixty-fourth meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA) Rome,
8–17 February 2005. summary_report_64_final.pdf. 2006; Available from:
ftp://ftp.fao.org/es/esn/jecfa/jecfa64_summary.pdf. Accessed November 24,
2014.
- 68. Exon JH. A review of the toxicology of
acrylamide. J Toxicol Environ Health B Crit Rev. 2006; 9(5): 397–412.
- 69. Parzefall W. Minireview on the toxicity of
dietary acrylamide. Food Chem Toxicol. 2008; 46(4): 1360–1364.
- 70. LoPachin RM. The changing view of acrylamide
neurotoxicity. NeuroToxicology. 2004; 25(4): 617–630.
- 71. Bashaw GJ, Klein R. Signaling from axon guidance
receptors. Cold Spring Harb Perspect Biol. 2010; 2(5): a001941.
- 72. Nóbrega-Pereira S, Marín O. Transcriptional
control of neuronal migration in the developing mouse brain. Cereb Cortex. 2009; 19 (Suppl
1): i107–i113.
- 73. Ribak CE, Korn MJ, Shan Z, Obenaus A. Dendritic
growth cones and recurrent basal dendrites are typical features of newly generated dentate
granule cells in the adult hippocampus. Brain Res. 2004; 1000(1-2): 195–199.
- 74. LoPachin RM, Barber DS, Gavin T. Molecular
mechanisms of the conjugated alpha,beta-unsaturated carbonyl derivatives: relevance to
neurotoxicity and neurodegenerative diseases. Toxicol Sci. 2008; 104(2): 235–249.
- 75. International Agency for Research on Cancer (IARC).
Some industrial chemicals. IARC Monographs on the Evaluation of Carcinogenic Risk of
Chemicals to Humans, vol.77, International Agency for Research on Cancer, Lyon, France,
2000: 469–486.
- 76. Bakhiya N, Abraham K, Gürtler R, Appel KE, Lampen
A. Toxicological assessment of 3-chloropropane-1,2-diol and glycidol fatty acid esters in
food. Mol Nutr Food Res. 2011; 55(4): 509–521.
- 77. Bundesinstitut für Risikobewertung (BfR). Initial
evaluation of the assessment of levels glycidol fatty acid esters detected in refined
vegetable fats. BfR Opinion No. 007/2009. 2009.
- 78. National Toxicology Program (NTP). Toxicology and
Carcinogenesis Studies of Glycidol (CAS No. 556-52-5) In F344/N Rats and B6C3F1 Mice
(Gavage Studies). Natl Toxicol Program Tech Rep Ser. 1990; 374: 1–229.
- 79. Shiraki A, Akane H, Ohishi T, et al. Similar
distribution changes of GABAergic interneuron subpopulations in contrast to the different
impact on neurogenesis between developmental and adult-stage hypothyroidism in the
hippocampal dentate gyrus in rats. Arch Toxicol. 2012; 86(10): 1559–1569.
- 80. Akane H, Shiraki A, Imatanaka N, et al. Glycidol
induces axonopathy and aberrations of hippocampal neurogenesis affecting late-stage
differentiation by exposure to rats in a framework of 28-day toxicity study. Toxicol Lett.
2014; 224(3): 424–432.
- 81. Sun J, Sun J, Ming GL, Song H. Epigenetic
regulation of neurogenesis in the adult mammalian brain. Eur J Neurosci. 2011; 33(6):
1087–1093.
- 82. Covic M, Karaca E, Lie DC. Epigenetic regulation
of neurogenesis in the adult hippocampus. Heredity (Edinb). 2010; 105(1): 122–134.
- 83. Laird PW, Jaenisch R. DNA methylation and cancer.
Hum Mol Genet. 1994; 3(Spec No): 1487–1495.
- 84. Jones PL, Veenstra GJ, Wade PA, et al. Methylated
DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet. 1998;
19(2): 187–191.
- 85. Ceccatelli S, Bose R, Edoff K, Onishchenko N,
Spulber S. Long-lasting neurotoxic effects of exposure to methylmercury during
development. J Intern Med. 2013; 273(5): 490–497.
- 86. Aschner M, Erikson KM, Herrero Hernández E,
Tjalkens R. Manganese and its role in Parkinson’s disease: from transport to
neuropathology. Neuromolecular Med. 2009; 11(4): 252–266.
- 87. Dobson AW, Erikson KM, Aschner M. Manganese
neurotoxicity. Ann N Y Acad Sci. 2004; 1012: 115–128.
- 88. Wang L, Ohishi T, Shiraki A, et al. Developmental
exposure to manganese chloride induces sustained aberration of neurogenesis in the
hippocampal dentate gyrus of mice. Toxicol Sci. 2012; 127(2): 508–521.
- 89. Wang L, Shiraki A, Itahashi M, et al. Aberration
in epigenetic gene regulation in hippocampal neurogenesis by developmental exposure to
manganese chloride in mice. Toxicol Sci. 2013; 136(1): 154–165.
- 90. Gulyás AI, Buzsáki G, Freund TF, Hirase H.
Populations of hippocampal inhibitory neurons express different levels of cytochrome c.
Eur J Neurosci. 2006; 23(10): 2581–2594.
- 91. Bøttger P, Tracz Z, Heuck A, Nissen P,
Romero-Ramos M, Lykke-Hartmann K. Distribution of Na/K-ATPase alpha 3 isoform, a
sodium-potassium P-type pump associated with rapid-onset of dystonia parkinsonism (RDP) in
the adult mouse brain. J Comp Neurol. 2011; 519(2): 376–404.
- 92. Trockenbacher A, Suckow V, Foerster J, et al.
MID1, mutated in Opitz syndrome, encodes an ubiquitin ligase that targets phosphatase 2A
for degradation. Nat Genet. 2001; 29(3): 287–294.
- 93. Krauss S, Foerster J, Schneider R, Schweiger S.
Protein phosphatase 2A and rapamycin regulate the nuclear localization and activity of the
transcription factor GLI3. Cancer Res. 2008; 68(12): 4658–4665.
- 94. Hasenpusch-Theil K, Magnani D, Amaniti EM, Han L,
Armstrong D, Theil T. Transcriptional analysis of Gli3 mutants identifies Wnt target genes
in the developing hippocampus. Cereb Cortex. 2012; 22(12): 2878–2893.
- 95. Quaderi NA, Schweiger S, Gaudenz K, et al. Opitz
G/BBB syndrome, a defect of midline development, is due to mutations in a new RING finger
gene on Xp22. Nat Genet. 1997; 17(3): 285–291.
- 96. Granata A, Quaderi NA. The Opitz syndrome gene
MID1 is essential for establishing asymmetric gene expression in Hensen’s node. Dev Biol.
2003; 258(2): 397–405.
- 97. Park JI, Tsai SY, Tsai MJ. Molecular mechanism of
chicken ovalbumin upstream promoter-transcription factor (COUP-TF) actions. Keio J Med.
2003; 52(3): 174–181.
- 98. O’Leary DD, Chou SJ, Sahara S. Area patterning of
the mammalian cortex. Neuron. 2007; 56(2): 252–269.
