Abstract
In the early 90s’, Europe was shaken by the fear that the prions from “mad cow disease”
(bovine spongiform encephalopathy) would transmit the disease to humans via beef products.
In 1996, the first variant Creutzfeldt-Jakob (vCJD) patients were described, and the same
year our Bovine Spongiform Encephalopathy (BSE) transmission studies to cynomolgus
macaques demonstrated that the BSE prion was highly infectious for primates, inducing
brain lesions identical to those observed in vCJD patients. These studies provided the
first experimental evidence that vCJD was BSE in humans. Subsequent studies established
the BSE/vCJD-infected cynomolgus macaque as a robust model to study the pathogenesis of
vCJD. We showed rapid adaptation of BSE prions to primates upon subsequent passage, and
their distribution in peripheral tissues and blood. Some key studies are summarized in the
present paper.
1. Transmission of Bovine Spongiform Encephalopathy (BSE) to CynomolgusMacaques
Reproduces vCJD: Establishment of a non-human Primate Model for vCJD
1.1. BSE, a New Disease in Cattle
In 1987, a new prion disease affecting dairy cattle was described in the United
Kindom1). Affected cows
presented signs of aggressiveness, anxiety, ataxia and were finally found recumbent. The
disease was rapidly classified in the group of “transmissible spongiform
encephalopathies”, or TSEs, due to the transmissibility of the disease2), as well as the similarities of
the neuropathological lesions and molecular hallmark with those found in sheep scrapie and
human CJD: neuronal death, spongiform changes, and accumulation of misfolded and
aggregated prion protein (termed PrPsc)3). PrPsc is the infectious form of the host prion protein PrP.
It is also called a prion (for “proteinaceaous infectious particle4)) or TSE agent. The number of
affected cows increased rapidly to top at a 37,280 diagnosed animals in the year of 1992
(OIE data). Thankfully, British epidemiologists recognized that BSE was due to the
consumption of prion-tainted meat and bone meal (MBM)5), and the first feed-ban was implemented in 1988,
prohibiting the feeding of ruminants with ruminant-derived MBM.
1.2. vCJD in Humans and Transmission of BSE to Non-human Primates
In 1991, BSE was reported in a domestic cat that presumably was contaminated via pet
food6). Transmission of
scrapie from small ruminants to cats had never been described, raising concern that BSE
might be more pathogenic than scrapie not only for cats, but also for humans. In order to
probe the cow-to-primates species barrier of the BSE agent, we inoculated cynomolgus
macaques (Macaca fascicularis) with BSE-infected cow brains at the French
Atomic Energy Commision (CEA).
In 1996, 10 young individuals were described in the UK and one in France, harboring an
unusual form of CJD that was coined variant CJD (vCJD)7,8). Besides patients being exceptionally young (adolescents and
young adults, while sporadic CJD (sCJD) affects people over the age of 60), they exhibited
unusual symptoms. Early symptoms were dysaesthesia, behavioral symptoms, depression,
ataxia, with myoclonus appearing later on, contrasting with the cognitive course of the
disease (memory impairment, dementia) preceding motor impairment, which is most frequently
observed in sCJD. Moreover, vCJD patients presented specific neuropathological features
with spongiosis and neuronal loss most evident in the basal ganglia and thalamus, and the
presence of PrP amyloid plaques (abundant in the cerebral cortex and cerebellum) that were
surrounded by vacuoles, giving them a flower-like appearance. These peculiar plaques were
called florid plaques7).
At the same time as the first vCJD patients were being described, we were examining the
brains of our 3 macaques that had all come down with disease 3 years after intracerebral
(IC) inoculation with BSE-infected cow brain. Clinical signs were characterized by
behavioral signs such as depression or edginess, as well as truncal ataxia (broad-based
gait, tremors) and myoclonus. Neuropathological examination of the brains of the
BSE-macaques revealed the presence of florid plaques and other neuropathological features
similar to those observed in vCJD patients (Fig.
1). Florid plaques were not present in the brains of macaques inoculated with
Kuru or sCJD, and thus were considered specific for infection by the BSE prion. Moreover,
PrPsc in BSE-infected macaques and vCJD patients exhibited a similar electrophoretic
pattern by western blot (Fig. 1).
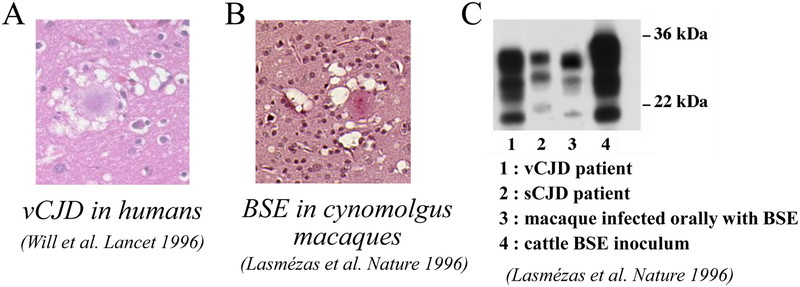
In summary, macaques infected by BSE reproduced the behavioral and motor symptoms, the
neuropathology and the biochemical signature of vCJD in humans. This study provided the
first experimental evidence supporting that vCJD was due to human infection by the BSE
agent9), and an
experimental model to study the new disease.
1.3. Determination of the Minimal Infectious BSE Dose in Non-human Primates
In a concerted European effort involving 5 laboratories including ours, the BSE-macaque
model was then used to evaluate the minimal amount of BSE-infected material necessary to
induce vCJD in primates. Results so far show that 5g of infectious BSE cattle brain is
sufficient to induce the disease in all recipient animals by the oral route, with 500 mg
yielding an incomplete attack rate10,11). The ID50 of BSE cattle brain is 200 mg for
cattle12). These results
suggest a low species barrier between cattle and non-human primates.
1.4. Adaptation of the BSE Agent to Non-human Primates: Consequences for Human
Health
The macaque BSE model provided an opportunity to evaluate the possible risk for humans of
secondary inter-human transmission of the BSE/vCJD prion. Accidental human-to-human
transmissions of sCJD, resulting in iatrogenic CJD (iCJD) has occurred in several
unfortunate circumstances (described in ref.13). One of them was the infection of children with CJD-contaminated human
growth hormone (hGH) extracted from cadaveric hypophyses. These iCJD patients had been
treated for short stature by injection of hGH in childhood, and 226 of them died of iCJD
as young adults, mainly in France and the USA13). Other dramatic iCJD cases had been linked to the
surgical implantation of dura-mater grafts, resulting in 228 deaths13). A few cases were also due to
corneal grafts and intracranial electrodes. Although all known iCJD cases prior to 2004
had been linked to contamination with central nervous system (CNS) tissue, the possibility
existed that the BSE agent would harbor a different distribution in primates than the sCJD
agent, thus representing a higher risk of transmission via organ/tissue grafts,
contamination of surgical instruments or even blood transfusion.
As a first step for risk assessment, we transmitted the BSE prion from macaque to macaque
via different routes. We also established a dose-response (incubation time) for the IC
route to provide a baseline for subsequent infectivity measurement studies. This
experiment showed that the BSE agent adapts rapidly to primates, as incubation periods
shortened from 3 to 1.5 years upon secondary passage at the highest dose14). It also showed that, for a
given amount of BSE material (40 mg BSE brain homogenate), the incubation period was the
same whether inoculation was done by the IC or the intravenous (IV) route.
2. The Cynomolgus Macaque as a model to Understand the Pathogenesis of Variant
Creutzfeldt-Jakob Disease (vCJD) and Model Risk of Interhuman Transmission
2. 1. Distribution of Prions in Tissues and Organs of BSE/vCJD Macaques after Oral or
Intravenous (IV) Inoculation
We compared second passage macaques inoculated with BSE prions by the oral or IV
routes15). PrPsc was
detected by immunohistochemistry and by ELISA after “scrapie associated fibril” (SAF)
purification15). In
addition to the brain, we detected PrPsc in spleen, tonsils, intestine and sciatic nerve
in amounts that did not depend on the inoculation route, with the exceptions of the spleen
where PrPsc amounts were up to 4% the amounts found in the brain after IV inoculation, and
up to 0.2% those of the brain after oral dosing15).
2. 2. Distribution of Prions in Tissues and Organs of vCJD, Sporadic and Iatrogenic
CJD Infected Macaques
We also infected macaques with vCJD, sCJD, iCJD16). As determined earlier, BSE and vCJD prions correspond to
the same prion strain, and one or the other denomination is used depending on the species
of origin for the brain tissue used as inoculum. All prion strains were inoculated in the
same manner (intracerebral and intratonsillar combined), in order to be able to directly
compare tissue distribution of PrPsc between strains.
Disease-associated PrP deposits were detected by immunocytochemistry in various organs.
They were found in the Peyer’s patches of the gut and other lymphoreticular system (LRS)
tissue of BSE/vCJD infected animals (Fig. 2). By
PET-blot, we showed that these deposits corresponded to proteinase K-resistant PrP, a
biochemical subset of PrPsc. Interestingly, not all Peyer’s patches of a single animal
were PrPsc positive (Fig. 2), showing that
PrPsc-negative LRS tissue biopsies may lead to false negative diagnostic results.
Pathological PrP deposits were also detected in the enteric nervous system in macaques
infected with all prion strains. Fig. 3 shows
the localization of pathological PrP in the pericarya of neurons of the myenteric plexus,
as well as in small nerve fibers of the inner muscular layer of the intestine.
Pathological PrP deposits were also found in peripheral nerves and in muscle for all CJD
strains (Table 1). In the peripheral nerves,
they were found mostly at the surface of Schwann cells (Fig. 4). In muscle, they were localized to specific foci in the vicinity of
nerve fibers (Fig. 5). Our results suggest that
the heterogeneous, patchy distribution of pathological PrP deposits in muscles corresponds
to the distribution zones of motor end plates. This study provides a possible explanation
for the variably positive detection of pathological PrP in muscle samples of sCJD
patients17).


PrPsc amounts were also measured semi-quantitatively using a sensitive biochemical
detection method including phosphotungstic acid precipitation as a concentration method,
and western blot or ELISA detection16). This method revealed the presence of PrPsc in the spleen
of the sCJD infected macaque and tonsils of the iCJD infected macaque, but no PrPsc could
be detected in lymph nodes and Peyer’s patches of these animals, a result most likely due
to the presence of PrPsc amounts at the threshold of detection in the LRS of sCJD and iCJD
macaques, and sampling variations. These results are summarized in Table 1.
In summary, PrPsc was detectable at high levels in organs and tissues of the LRS only in
BSE/vCJD infected animals (0.1% to 10% of the amounts found in the brains of the same
animals). We interpreted these results as the BSE prion being highly lymphotropic in
primates. These findings correlated indeed with the tonsils, spleens and appendices of
vCJD patients being found positive for PrPsc18,19,20). We therefore proposed that LRS tissues be considered
‘high-risk’ in vCJD patients only.
However, lower amounts of PrPsc were detected in adrenals, muscles and intestinal tissue
of macaques infected with BSE/vCJD as well as sCJD and iCJD, associated with peripheral
nerves. Levels were less than 10,000 times lower than brain PrPres levels (<0.001%). We
therefore proposed that these tissues be considered “low-risk” for all CJD patients.
Our results expanded upon observations made in vCJD patients that PrPsc is detectable in
tonsils, emphasizing that BSE prions are largely lymphotropic in primates, and may
replicate in lymph notes, tonsils, spleen and Peyer’s patches before the symptomatic
phase. Our subsequent studies confirmed that lymph node biopsies of BSE-inoculated
macaques were positive for PrPsc prior to the onset of clinical signs (see below). In
another study, gut-associated lymphoid tissue and gut-draining lymph nodes were found
positive for PrPsc within one year of oral infection of macaques with cattle BSE21). On the other hand, distribution
of PrPsc in muscle of macaques inoculated with vCJD, sCJD and iCJD suggests a centrifugal
spread of prions from the CNS to muscle motor plates via motor nerves, occurring after CNS
invasion by prions. In addition, it is probable that centripetal spread of prions via
peripheral nerves also occurs in earlier stages of infection stochastically from various
points of entry, even in the absence of prior LRS replication. We demonstrated this
paradigm earlier in severely immunodeficient (SCID) mice infected with mouse-adapted
scrapie22). Moreover,
spread from the gut to the CNS via autonomic nerve fibers has been shown in experimental
scrapie and BSE23,24,25). Fig.
6 illustrates the distribution and proposed propagation of prions in our
non-human primate model.

The lymphotropic properties of BSE prions raised the important question of the presence
of infectivity in blood of vCJD patients.
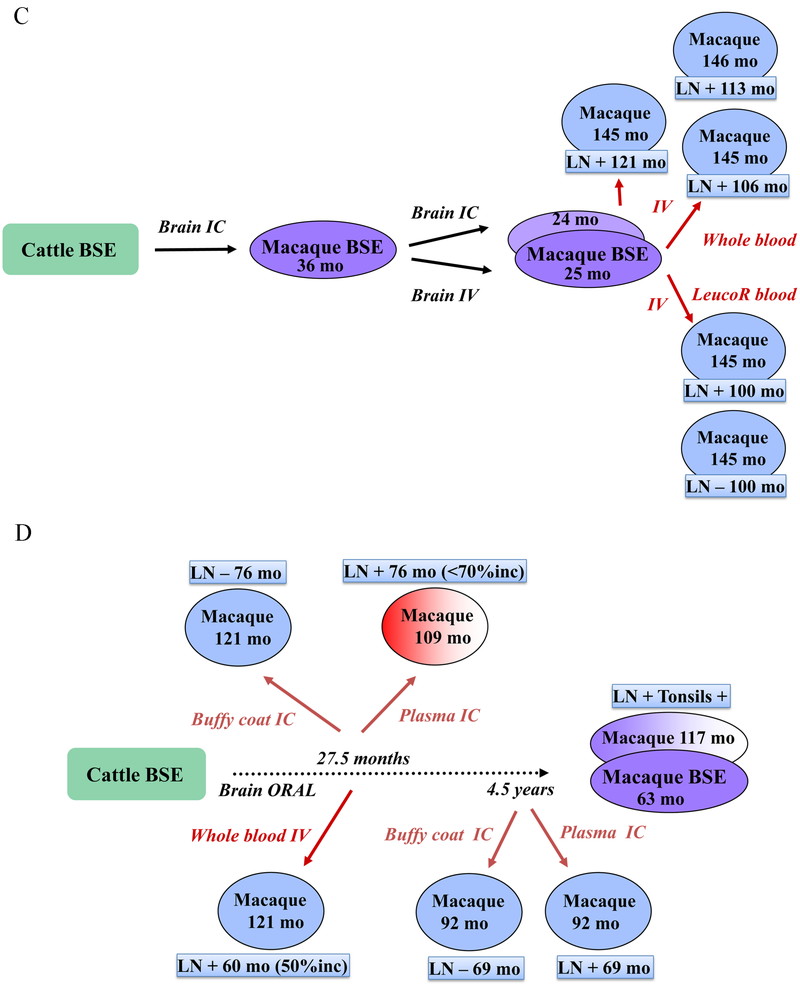
We initiated a large blood transfusion study where whole blood, white blood cells or
plasma from either vCJD patients or BSE/vCJD macaques was injected by IV or IC to
recipient macaques. This experiment led to most macaques surviving over prolonged periods
of time (>10 years), and few coming down with BSE/vCJD or intercurrent illnesses. These
studies continued after the author of this manuscript left the CEA. Interim transmission
results are shown in Fig. 7, and some important
observations were as follows. Blood depleted for red blood cells (RBC) from a vCJD patient
(7.5 mL) injected intravenously did not result in any clinical disease in the recipient
macaque after 10 years, yet this animal harbored positive IHC staining in the inguinal
lymph nodes (LNs). Another macaque, who had received 25 mL of RBC-depleted blood
intravenously from another vCJD patient, died suddenly at 42 months after inoculation, and
harbored PrPsc positive inguinal LNs. Two other animals, that received 500 µL of buffy
coat (BC) from vCJD patients by IC, were still alive 10 years after inoculation (with
PrPsc positive LNs, Fig. 7A). A whole blood
transfusion of 40 mL from a vCJD macaque (who died 3 years after intracerebral +
intratonsillar inoculation of human vCJD brain homogenate) induced clinical signs of vCJD
in the recipient macaque 66 months after the transfusion. Inguinal lymph nodes biopsies
had been positive since 45 months, i.e. 75% of the incubation period (Fig. 7B). Other macaques transfused with blood from BSE-macaques
survived more than 10 years, but some had positive LNs (Fig. 7C). Another notable result was the transmission of BSE infection by the
plasma from a macaque that had been dosed orally with cattle BSE 27.5 months earlier. The
donor macaque died with a behavioral syndrome of self-injury 117 months after challenge
with a diagnosis of probable BSE, hence infectivity was present in its blood at a quarter
of the incubation period (Fig. 7D).

In 2004, the first transfusion-related case of vCJD was described in a patient who had
been transfused with non-leucoreduced red blood cells from a donor who developed vCJD
3.5 years after the donation26). A total of four transfusion-related vCJD transmissions
have been reported to date27).
Acknowledgments
I thank the many people who supported and participated in this work at the CEA: Dominique
Dormont, Jean-Philippe Deslys, Christian Herzog, Nathalie Lescoutra, Nicole Salès, Emmanuel
Comoy, René Rioux. I also thank Ray Bradley and Michael Dawson for providing BSE-infected
cattle brain homogenates. I am thankful to Robert Will and Nicolas Kopp for providing vCJD
samples, and to James Ironside for his collaboration on the neuropathology of BSE-infected
macaques. I am grateful to my European collaborators Maurizio Pocchiari, Gerhard Hunsmann,
Johannes Löwer, Pär Bierke, Loredana Ingrosso, Uwe Hahmann, Dirk Motzkus, Edgar Holznagel,
for their friendship and for embarking on this challenging project.
References
- 1. Wells GAH, Scott AC, Johnson CT, et al. A novel
progressive spongiform encephalopathy in cattle. Vet Rec. 1987;
121: 419–420. doi: 10.1136/vr.121.18.419
- 2. Fraser H, Mcconnell I, Wells GAH, Dawson M.
Transmission of bovine spongiform encephalopathy to mice. Vet Rec. 1988;
123: 472. doi: 10.1136/vr.123.18.472
- 3. Hope J, Reekie LJD, Hunter N, et al. Fibrils from
brains of cows with new cattle disease contain scrapie-associated protein.
Nature. 1988; 336: 390–392. doi:
10.1038/336390a0
- 4. Prusiner SB. Novel proteinaceous infectious
particles cause scrapie. Science. 1982; 216: 136–144. doi:
10.1126/science.6801762
- 5. Wilesmith JW, Wells GAH, Cranwell MP, Ryan JBM.
Bovine spongiform encephalopathy: epidemiological studies. Vet Rec. 1988;
123: 638–644.
- 6. Pearson GR. Feline Spongiform Encephalopathy.
Vet Rec. 1991; 128: 532. doi:
10.1136/vr.128.22.532-b
- 7. Will RG, Ironside JW, Zeidler M, et al. A new
variant of Creutzfeldt-Jakob disease in the UK. Lancet. 1996;
347: 921–925. doi: 10.1016/S0140-6736(96)91412-9
- 8. Chazot G, Broussolle E, Lapras C, Blätter T,
Aguzzi A, Kopp N. New variant of Creutzfeldt-jakob disease in a 26 year-old French man.
Lancet. 1996; 347: 1181. doi:
10.1016/S0140-6736(96)90638-8
- 9. Lasmézas CI, Deslys J-P, Robain O, et al. BSE
transmission to macaques. Nature. 1996; 381: 743–744. doi:
10.1038/381743a0
- 10. Lasmézas CI, Comoy E, Hawkins S, et al. Risk of
oral infection with bovine spongiform encephalopathy agent in primates.
Lancet. 2005; 365: 781–783. doi:
10.1016/S0140-6736(05)71001-1
- 11. Holznagel E, Yutzy B, Schulz-Schaeffer W, Kruip
C, Hahmann U, Bierke P, Torres JM, Kim YS, Thomzig A, Beekes M, Hunsmann G, Loewer J.
Foodborne transmission of bovine spongiform encephalopathy to nonhuman primates. Emerg
Infect Dis. 2013;19(5):712–20. doi: 10.3201/eid1905.120274 PubMed PMID: 23647575; PubMed
Central PMCID: PMCPMC3647490.
- 12. Wells GA, Konold T, Arnold ME, et al. Bovine
spongiform encephalopathy: the effect of oral exposure dose on attack rate and incubation
period in cattle. J Gen Virol. 2007; 88: 1363–1373. doi:
10.1099/vir.0.82421-0
- 13. Brown P, Brandel JP, Sato T, et al. Iatrogenic
Creutzfeldt-Jakob disease, final assessment. Emerg Infect Dis. 2012;
18: 901–907.; published online; doi:
10.3201/eid1806.120116
- 14. Lasmézas CI, Fournier JG, Nouvel V, Boe H, Marce
D, Lamoury F, Kopp N, Hauw JJ, Ironside J, Bruce M, Dormont D, Deslys JP. Adaptation of
the bovine spongiform encephalopathy agent to primates and comparison with
Creutzfeldt-Jakob disease: implications for human health. Proc Natl Acad Sci U S A.
2001;98(7):4142–7. Epub 2001/03/22. doi: 10.1073/pnas.041490898 [pii]. PubMed PMID:
11259641; PubMed Central PMCID: PMC31193.
- 15. Herzog C, Sales N, Etchegaray N, et al. Tissue
distribution of bovine spongiform encephalopathy agent in primates after intravenous or
oral infection. Lancet. 2004; 363: 422–428. doi:
10.1016/S0140-6736(04)15487-1
- 16. Herzog C, Riviere J, Lescoutra-Etchegaray N, et
al. PrPTSE distribution in a primate model of variant, sporadic, and iatrogenic
Creutzfeldt-Jakob disease. J Virol. 2005; 79: 14339–14345.
doi: 10.1128/JVI.79.22.14339-14345.2005
- 17. Glatzel M, Abela E, Maissen M, Aguzzi A.
Extraneural pathologic prion protein in sporadic Creutzfeldt-Jakob disease. N Engl
J Med. 2003; 349: 1812–1820. doi:
10.1056/NEJMoa030351
- 18. Hilton DA, Fathers E, Edwards P, Ironside JW,
Zajicek J. Prion immunoreactivity in appendix before clinical onset of variant
Creutzfeldt-Jakob disease. Lancet. 1998; 352: 703–704. doi:
10.1016/S0140-6736(98)24035-9
- 19. Ironside JW, Head MW, Bell JE, McCardle L, Will
RG. Laboratory diagnosis of variant Creutzfeldt-Jakob disease.
Histopathology. 2000; 37: 1–9. doi:
10.1046/j.1365-2559.2000.00946.x
- 20. Wadsworth JD, Joiner S, Hill AF, et al. Tissue
distribution of protease resistant prion protein in variant Creutzfeldt-Jakob disease
using a highly sensitive immunoblotting assay. Lancet. 2001;
358: 171–180. doi: 10.1016/S0140-6736(01)05403-4
- 21. Holznagel E, Schulz-Schaeffer W, Yutzy B, Bierke P,
Hunsmann G, Löwer J. Spread of BSE prions in cynomolgus monkeys (Macaca fascicularis)
after oral transmission. Prion 2009, Thessaloniki, Greece. 2009.
- 22. Lasmézas CI, Cesbron JY, Deslys JP, et al. Immune
system-dependent and -independent replication of the scrapie agent. J
Virol. 1996; 70: 1292–1295.
- 23. McBride PA, Schulz-Schaeffer WJ, Donaldson M, et
al. Early spread of scrapie from the gastrointestinal tract to the central nervous system
involves autonomic fibers of the splanchnic and vagus nerves. J Virol.
2001; 75: 9320–9327. doi: 10.1128/JVI.75.19.9320-9327.2001
- 24. Kaatz M, Fast C, Ziegler U, et al. Spread of
classic BSE prions from the gut via the peripheral nervous system to the brain. Am
J Pathol. 2012; 181: 515–524. doi:
10.1016/j.ajpath.2012.05.001
- 25. Holznagel E, Yutzy B, Kruip C, Bierke P,
Schulz-Schaeffer W, Lower J. Foodborne-Transmitted Prions From the Brain of Cows With
Bovine Spongiform Encephalopathy Ascend in Afferent Neurons to the Simian Central Nervous
System and Spread to Tonsils and Spleen at a Late Stage of the Incubation Period.
J Infect Dis. 2015; 212: 1459–1468. doi:
10.1093/infdis/jiv232
- 26. Llewelyn CA, Hewitt PE, Knight RS, et al.
Possible transmission of variant Creutzfeldt-Jakob disease by blood transfusion.
Lancet. 2004; 363: 417–421. doi:
10.1016/S0140-6736(04)15486-X
- 27. Urwin P, Mackenzie J. CA L, Will R, Hewitt P.
Creutzfeldt-Jakob disease and blood transfusion: updated results of the UK Transfusion
Medicine epidemiology Review Study. Vox Sang. 2016; 110:
310–316.; published online; doi: 10.1111/vox.12371


