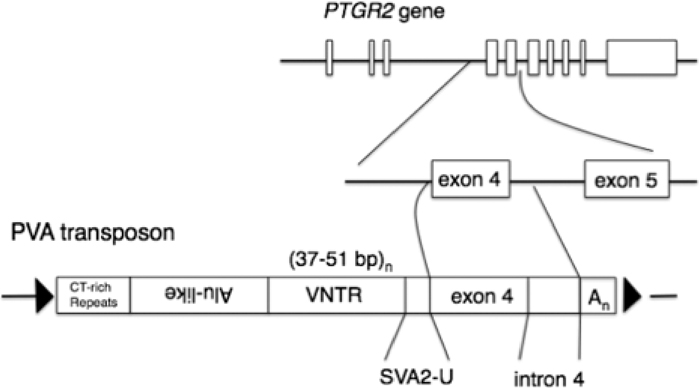
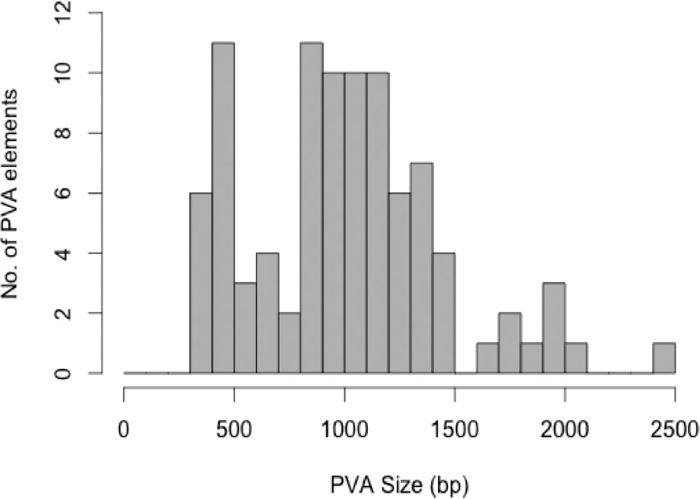
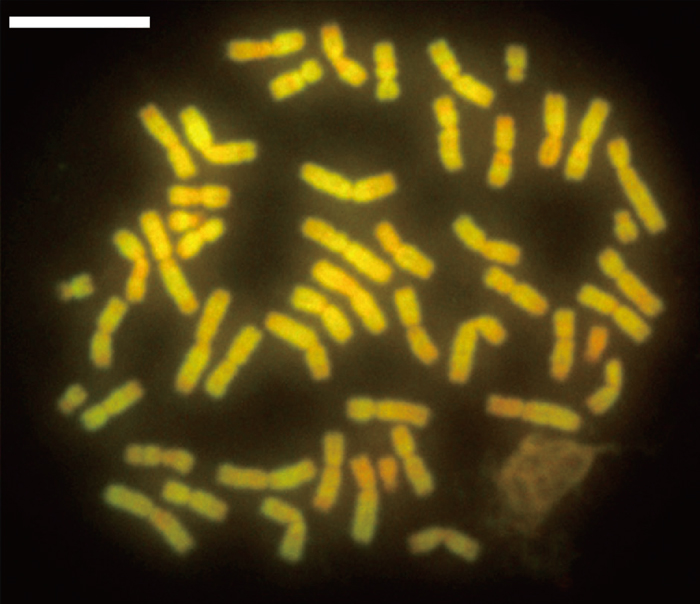
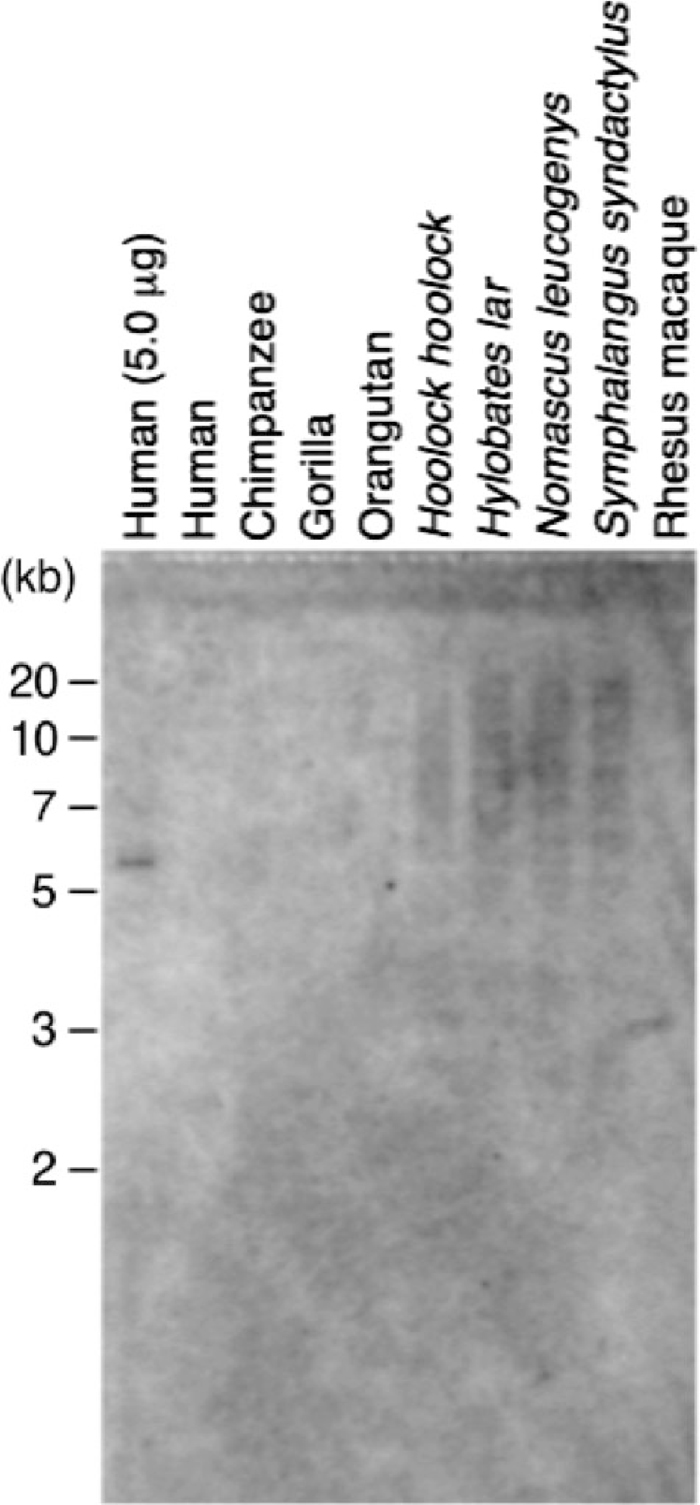
| 1 | 5' truncation | 323 | 11 | GL397437, +, 794473 – 794795 | 48 | canonical | 1003 | 14 | GL397272, +, 37697076 – 37698078 |
| 2 | 5' truncation | 324 | 9 | GL397437, +, 803094 – 803417 | 49 | canonical | 1026 | 12 | GL397355, +, 7377493 – 7378518 |
| 3 | 5' truncation | 366 | 13 | GL397389, +, 1156295 – 1156660 | 50 | canonical | 1040 | 13 | GL397301, +, 3412318 – 3413357 |
| 4 | 5' truncation | 382 | 4 | GL397450, +, 71920 – 72301 | 51 | canonical | 1045 | 12/13 | GL397442, –, 233512 – 234556 |
| 5 | 5' truncation | 391 | 21 | GL397302, +, 9602076 – 9602466 | 52 | canonical | 1054 | 10 | GL397296, +, 605120 – 606173 |
| 6 | 5' truncation | 399 | 14 | GL397515, –, 313131 – 313529 | 53 | 5' truncation | 1057 | 16 | GL397285, +, 17358306 – 17359362 |
| 7 | 5' truncation | 401 | 14 | GL397555, +, 195346 – 195746 | 54 | 3' trunsduction | 1070 | 16 | GL397318, +, 1025408 – 1026477 |
| 8 | 5' truncation | 402 | 18 | GL397299, +, 2732709 – 2733110 | 55 | 3' trunsduction | 1077 | 14 | GL397305, +, 9677654 – 9678730 |
| 9 | 5' truncation | 417 | 11/12 | GL397262, +, 13609205 – 13609621 | 56 | canonical | 1088 | 13 | GL397326, +, 14041076 – 14042163 |
| 10 | 5' truncation | 432 | 23 | ADFV01139252, –, 12256 – 12687 | 57 | canonical | 1095 | 6 | GL397306, +, 4087605 – 4088699 |
| 11 | 5' truncation | 434 | 15 | GL397340, –, 9706804 – 9707237 | 58 | 5' truncation | 1111 | 12 | GL397302, –, 9342609 – 9343719 |
| 12 | 5' truncation | 449 | 15 | GL397261, –, 54947513 – 54947961 | 59 | canonical | 1111 | 11 | GL397499, –, 735571 – 736681 |
| 13 | 5' truncation | 474 | 3 | GL397287, –, 20289161 – 20289634 | 60 | 5' truncation | 1135 | 4 | GL397283, +, 24873343 – 24874477 |
| 14 | 5' truncation | 477 | 10 | GL397367, +, 4311612 – 4312088 | 61 | 3' trunsduction | 1139 | 12 | GL397303, –, 18558538 – 18559676 |
| 15 | 5' truncation | 483 | 10 | GL397359, –, 4224834 – 4225316 | 62 | canonical | 1139 | 19 | GL397372, +, 951128 – 952266 |
| 16 | 5' truncation | 486 | 26 | GL397382, +, 1672651 – 1673136 | 63 | canonical | 1144 | 14 | ADFV01136848, +, 26939 – 28082 |
| 17 | 5' truncation | 499 | 13 | GL397322, –, 3845419 – 3845917 | 64 | canonical | 1155 | 16/17 | GL397267, +, 30459064 – 30460218 |
| 18 | 5' truncation | 516 | 3 | GL397268, +, 43078580 – 43079095 | 65 | canonical | 1169 | 11 | GL397453, +, 1546227 – 1547395 |
| 19 | 5' truncation | 536 | 19 | GL397381, +, 1103186 – 1103721 | 66 | 3' trunsduction | 1185 | 10 | GL397356, +, 2962496 – 2963680 |
| 20 | 5' truncation | 539 | 19 | GL397326, –, 9270245 – 9270783 | 67 | canonical | 1192 | 6 | GL397584, +, 90896 – 92087 |
| 21 | 5' truncation | 625 | 13 | GL397301, +, 444008 – 444632 | 68 | canonical | 1205 | 17 | GL397373, +, 1913876 – 1915080 |
| 22 | 5' truncation | 638 | 11 | GL397368, +, 5768930 – 5769567 | 69 | canonical | 1217 | 20 | GL397475, +, 1281245 – 1282461 |
| 23 | 5' truncation | 673 | 10 | GL397477, –, 1710403 – 1711075 | 70 | canonical | 1259 | 14 | GL397332, +, 9132309 – 9133567 |
| 24 | unknown | 700 | 13 | GL397335, –, 9618886 – 9619585 | 71 | 3' trunsduction | 1275 | 16/17 | GL397272, +, 3000911 – 3002185 |
| 25 | 5' truncation | 748 | 9 | GL397323, –, 3593499 – 3594246 | 72 | canonical | 1277 | 14 | GL397306, +, 18017623 – 18018899 |
| 26 | 5' truncation | 771 | 11 | GL397370, +, 3420251 – 3421021 | 73 | canonical | 1284 | 16 | GL397261, +, 61072814 – 61074097 |
| 27 | 5' truncation | 806 | 6 | GL398191, +, 17016 – 17821 | 74 | canonical | 1320 | 14 | GL397266, +, 8121916 – 8123235 |
| 28 | 5' truncation | 827 | 11 | GL397437, –, 1941131 – 1941957 | 75 | canonical | 1324 | 7 | GL397544, –, 459850 – 461173 |
| 29 | canonical | 833 | 17 | GL397277, +, 30334916 – 30335748 | 76 | canonical | 1327 | 20 | GL397301, +, 3434467 – 3435793 |
| 30 | canonical | 845 | 13 | GL397279, +, 22935106 – 22935963 | 77 | canonical | 1362 | 12 | GL397316, –, 2729091 – 2730452 |
| 31 | canonical | 864 | 11 | GL397537, –, 894729 – 895592 | 78 | canonical | 1372 | 15 | GL397309, +, 10757450 – 10758821 |
| 32 | 5' truncation | 868 | 20 | GL397504, –, 108248 – 109115 | 79 | canonical | 1380 | 16/18 | GL397274, +, 14619951 – 14621330 |
| 33 | canonical | 877 | 12 | GL397312, +, 5975854 – 5976730 | 80 | 5' trunsduction | 1398 | 14 | GL397393, –, 2492234 – 2493631 |
| 34 | canonical | 889 | 14/16 | GL397272, –, 3879557 – 3880445 | 81 | canonical | 1401 | 12 | GL398298, +, 10757 – 12157 |
| 35 | canonical | 895 | 15 | GL397328, –, 31778 – 32672 | 82 | 3' trunsduction | 1416 | 13 | GL397287, +, 8426737 – 8428152 |
| 36 | 5' truncation | 898 | 12 | GL397275, +, 1489372 – 1490269 | 83 | 5' truncation | 1426 | 10 | GL397281, –, 22760805 – 22762230 |
| 37 | canonical | 900 | 9 | GL397479, +, 1316872 – 1317771 | 84 | 3' trunsduction | 1443 | 14/15 | GL397334, –, 9508105 – 9509547 |
| 38 | canonical | 911 | 11 | GL397296, +, 247203 – 248113 | 85 | canonical | 1658 | 12 | GL397276, –, 32330003 – 32331660 |
| 39 | canonical | 926 | 8/10 | GL397273, +, 1205654 – 1206579 | 86 | canonical | 1705 | 16 | GL397425, +, 1494179 – 1495883 |
| 40 | canonical | 957 | 12 | GL397288, –, 19518386 – 19519342 | 87 | 3' trunsduction | 1708 | 4 | GL397285, –, 26879474 – 26881181 |
| 41 | canonical | 964 | 18 | GL397299, +, 14146871 – 14147834 | 88 | 5' truncation | 1891 | 13 | GL397303, –, 6304456 – 6306346 |
| 42 | canonical | 973 | 6 | GL397266, +, 24563664 – 24564636 | 89 | canonical | 1902 | 14 | GL397266, +, 47555703 – 47557604 |
| 43 | canonical | 979 | 17 | GL397275, +, 18454251 – 18455229 | 90 | 5' truncation | 1982 | 15 | GL397402, +, 2093059 – 2095040 |
| 44 | canonical | 982 | 15 | GL397385, +, 1453448 – 1454429 | 91 | 5' trunsduction | 1995 | 15 | GL397544, +, 87629 – 89623 |
| 45 | canonical | 985 | 10 | GL397327, +, 3471001 – 3471985 | 92 | 5' trunsduction | 2030 | 13 | GL397314, –, 10577965 – 10579994 |
| 46 | canonical | 991 | 12 | GL397299, +, 1277922 – 1278912 | 93 | canonical | 2478 | 12 | GL397381, –, 917928 – 920405 |
| 47 | canonical | 991 | 18 | GL397318, –, 13178629 – 13179619 | | | | | |