MAIN
Paired box (Pax) 6 belongs to the Pax family of transcription factors, which are highly conserved and are involved in regulating the morphogenesis of many tissues and organs through ontogeny, playing pivotal roles in, for example, development of the eye, central nervous system and pancreas. Pax6 is so potent that ectopic expression of the mouse and Xenopus Pax6 genes can induce ectopic eye development in the fruit fly (Halder et al., 1995; Gehring and Ikeo, 1999). According to Cvekl and Callaerts (2017), the Pax6 gene is one of a small group that encodes master regulatory proteins that switch molecular networks, initiating the development of specific cell types. Elucidating the functional role of Pax6 during development is an essential step for the phylogenetic comparison of the developmental mechanism(s) in which it is involved.
Pax6, like most other members of the Pax family, consists of two DNA-binding domains, the paired domain (PD) and the homeodomain (HD). Compared to our accumulated knowledge about the structure and function of the HD, the PD is not well understood and needs further study. One such issue is to clarify the amino acid residues responsible for the function of the nuclear localization signal (NLS) in the PD (PD-NLS). Regarding the PD-NLS, Carrière et al. (1993, 1995) reported that deletion of exon 5 of quail (Coturnix coturnix japonica) Pax6 (Pax-QNR) significantly decreased the localization of the resulting Pax6 isoform in the nucleus. They predicted a very strong NLS in the amino acid sequence encoded by exon 5 of the Pax-QNR, which would be more active than the NLS that spans the N-terminus of the HD (HD-NLS, also called HNLS5 [LKRKLQR, residues L206 – R212]) (Carrière et al., 1995). Therefore, the function of Pax6 may heavily depend on the NLS in the PD domain. In this sense, it is crucial to localize the amino acid sequence comprising the NLS of the PD. Identifying that sequence would help us not only to infer the functional evolution of the PD domain, which has had a crucial role ever since the Tc1/mariner transposon was captured and domesticated in the ancient Pax family (Paixão-Côrtes et al., 2015), but also to design expression constructs, such as those harboring the nucleotide sequence encoding the NLS of the PD without any DNA-binding activity, to dissect the molecular function(s) of the PD in ontogeny.
According to Carrière et al. (1995), whose experiments were carried out in vitro using COS and quail embryonic cells, it is possible that all amino acid residues encoded by exon 5 of quail Pax6 can act as a NLS. If that is the case, an exceptionally long region of amino acid sequence would comprise the NLS, since 72 amino acid residues are encoded by exon 5 of quail Pax6 (gene ID: X70475.1; protein ID CAA49899.1), which is homologous to chick Pax6 exon 6 (Fig. 1A). NLSs are usually only seven to 10 residues in length: for example, the c-myc NLS contains only nine amino acid residues (PAAKRVKLD, Dang and Lee, 1988) and the p53 NLS contains only seven residues (PQPKKKP, Liang and Clarke, 1999). To clarify this issue, we now report our functional analysis in ovo to survey short amino acid sequences contributing to the PD-NLS in chick Pax6.
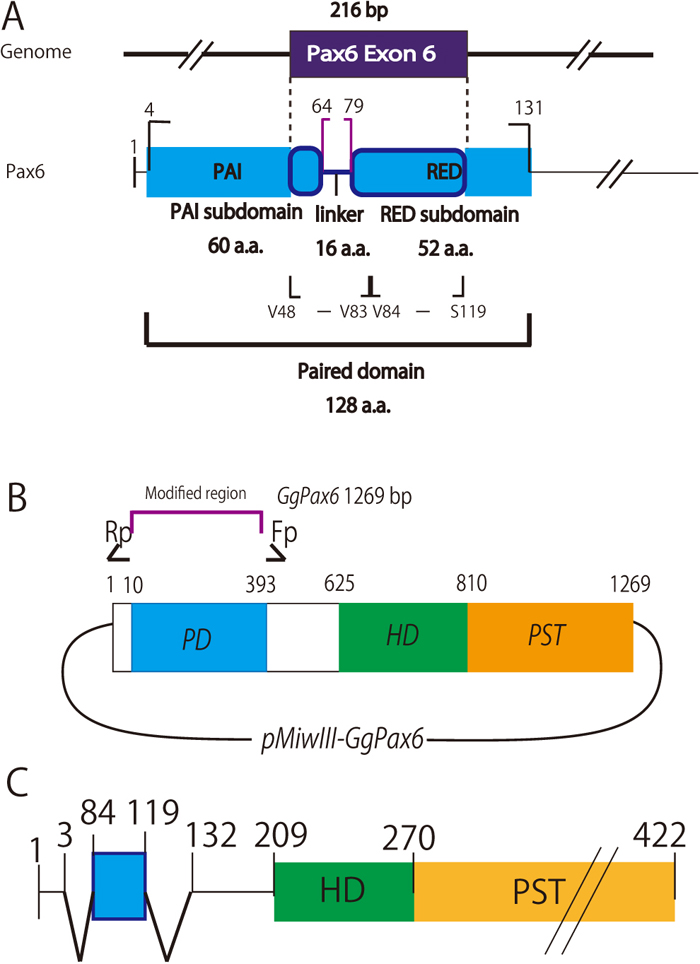
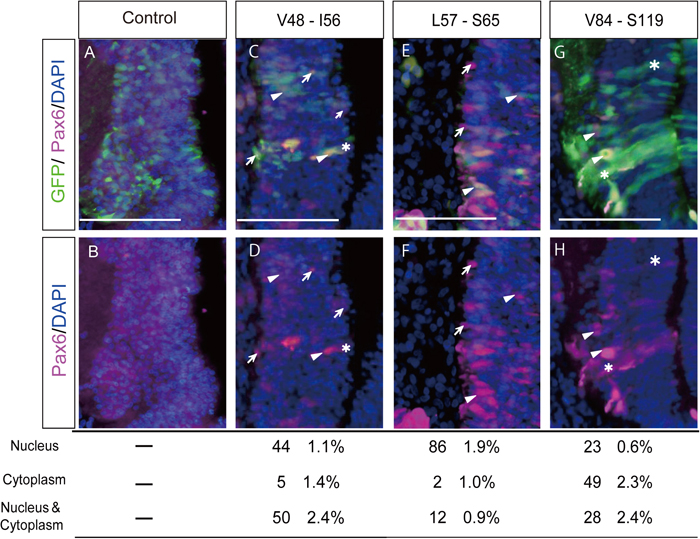
As noted above, exon 5 of Pax-QNR (Carrière et al., 1993, 1995) corresponds to exon 6 of chick (Gallus gallus domesticus) Pax6 (gene ID: XM_015286139; protein ID: XP_015141625). Full-length chick Pax6 cDNA was prepared using RT-PCR with mRNA obtained from E9 chick eyes and midbrains according to Mochii et al. (1998) and was inserted into the expression vector pMiwIII, a derivative of pMiwZ (Suemori et al., 1990; Wakamatsu et al., 1997; Matsunaga et al., 2000). This expression vector includes the nucleotide sequence that encodes virion protein 16 (Hirai et al., 2010), which has a robust transactivation activity that can be utilized in other experiments but does not affect the localization of the fused protein in transfected cells. From the resulting Pax6-Vp16 construct, plasmids expressing modified Pax6-VP16 transcription factors that are deficient in the PD sequence were prepared by inverse PCR (Fig. 1B). Next, exon 6 of chick Pax6 was inserted into the same position that the PD sequence had originally occupied. Using this resultant construct, new plasmids expressing modified Pax6-VP16 transcription factors that are deficient in the PD sequence except for the N-terminal half (V48 - V83) or the C-terminal half (V84 - S119) amino acid sequences encoded by exon 6, each as candidate NLS sequences from the PD, were prepared, also by inverse PCR, as shown in Fig. 1C. These modified Pax6s were termed (ΔPD+NLSV48-V83)-Pax6-VP16 (shortened to Pax6-NLSV48-V83) and (ΔPD+NLSV84-S119)-Pax6-VP16 (Pax6-NLSV84-S119), respectively. The primers used are summarized in Table 1.
Table 1. Primer sets used to produce modified Pax6
| Modified Pax6 | Template | Primer set |
|---|
| Pax6-ΔPD | pMiwIII-Pax6 | Fp - ACTGTTCTGCATGGTGGC |
| | Rp - AGCGAAAAGCAACAGATGG |
| Exon 6 N-terminal Pax6-NLSV48-V83 | pMiwIII-Pax6 (fragment inserted into pMiwIII-Pax6-ΔPD) | Fp - TCGAAGCTTGCCACCATGCAGAACAGTGTGTCGAATGGATGTGTGA GTAAAATTTTG |
| Rp - CTAGCTAGCAACTTCGGGAGTCGCTACTCTCG |
| Exon 6 C-terminal Pax6-NLSV84-S119 | pMiwIII-Pax6 (fragment inserted into pMiwIII-Pax6-ΔPD) | Fp - TCGAAGCTTGCCACCATGCAGAACAGTGTAAGCAAAATAGCGCAGT ATAAACGAGAGTG |
| Rp - CTAGCTAGCACTGGGTATGTTATCGTTGGTACAGACC |
| Pax6-NLSV48-S65 | pMiwIII-Pax6-NLSV48-V83 | Fp - AGCGAAAAGCAACAGATGGGTGCCGAC |
| | Rp - AGTAAAATTTTGGGCAGGTATTACGAAACTGGCTCC |
| Pax6-NLSI66-V83 | pMiwIII-Pax6-NLSV48-V83 | Fp - CCGCCACCATGCAGAACAGT |
| | Rp - ATCAGGCCCAGGGCGAT |
| Pax6-NLSV48-V53 | pMiwIII-Pax6-NLSV48-S65 | Fp - AGCGAAAAGCAACAGATGGGTGC |
| | Rp - CACACATCCATTCGACACACTGTTCT |
| Pax6-NLSV48-I56 | pMiwIII-Pax6-NLSV48-S65 | Fp - AGCGAAAAGCAACAGATGGG |
| | Rp - AATTTTACTCACACATCCATTCGA |
| Pax6-NLSL57-S65 | pMiwIII-Pax6-NLSV48-S65 | Fp - TTGGGCAGGTATTACGAAACTGGC |
| | Rp - ACTGTTCTGCATGGTGGCGG |
| Pax6-NLSI66-S74 | pMiwIII-Pax6-NLSI66-V83 | Fp - GCTAGCGAAAAGCAACAGATGGGT |
| | Rp - ACTACCTCCGATCGCCCTGG |
| Pax6-NLSK75-V83 | pMiwIII-Pax6-NLSI66-V83 | Fp - AAGCCGAGAGTAGCGACTCCC |
| | Rp - ACTGTTCTGCATGGTGGCGG |
| Pax6-NLSV78-V83 | pMiwIII-Pax6-NLSI66-V83 | Fp - GTAGCGACTCCCGAAGTTGCTAGC |
| | Rp - ACTGTTCTGCATGGTGGCGG |
| Pax6-NLSC52-Y61 | pMiwIII-Pax6-NLSV48-S65 | Fp1 - TGTGTGAGTAAAATTTTGGGCAGGT |
| | Rp1 - ACTGTTCTGCATGGTGGCGG |
| | Fp2 - AGCGAAAAGCAACAGATGGGTG |
| | Rp2 - GTAATACCTGCCCAAAATTTTACTCA |
| Pax6-NLSY61-A70 | pMiwIII-Pax6-NLSV48-V83 | Fp1 - TACGAAACTGGCTCCATCAGGC |
| | Rp1 - ACTGTTCTGCATGGTGGCGG |
| | Fp2 - GCTAGCGAAAAGCAACAGATGGGT |
| | Rp2 - CGCCCTGGGCCTGATGGA |
| Pax6-NLSA70-A79 | pMiwIII-Pax6-NLSI66-V83 | Fp1 - GCGATCGGAGGTAGTAAGCCGAG |
| | Rp1 - ACTGTTCTGCATGGTGGCGG |
| | Fp2 - GCTAGCGAAAAGCAACAGATGGGT |
| | Rp2 - CGCTACTCTCGGCTTACTACCTCCG |
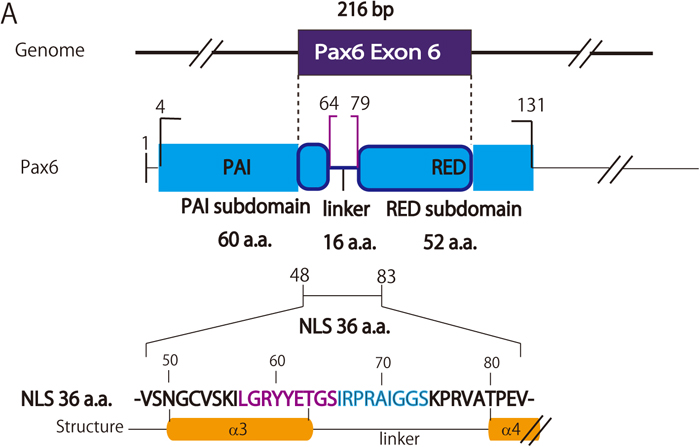
These constructs were electroporated into developing chick midbrains at HH 9-12 stages (Hamburger and Hamilton, 1992), at which the expression of endogenous Pax6 can hardly be detected (Fig. 2A and 2B). Localization of the modified Pax6 in nuclei was determined by observing co-localization of the signals of DAPI (to identify nuclei) and Pax6, which was detected by an antibody against that protein. Because we first found that the N-terminal half of the putative NLS (V48 - V83) had a higher activity to drive Pax6-NLSV48-V83 into nuclei, to obtain a more detailed map of the NLS of the PD, the N-terminal half (V48 - V83) encoded by exon 6 was scanned with 6-9-amino acid windows since NLSs are usually only seven to 10 residues in length, as mentioned above. Each expression vector was prepared by inverse PCR (Table 1). Representative examples are shown in Fig. 2. We found that the chick PD-NLS activity of Pax6 broadly resides over a 36-amino acid sequence between V48 and V83 (Fig. 3) compared to a lower NLS activity shown by the C-terminal half of the putative NLS (V84-S119). The amino acid sequence between V48 and V83 mainly corresponds to the linker region of the two subdomains in the PD, namely PAI and RED, and to the C- and N-terminal amino acid residues of the PAI and RED subdomains, respectively (Fig. 4). This linker region has been reported to have no specific function between these subdomains (Xu et al., 1999). According to the crystal structure of the human Pax6 PD-DNA complex reported by Xu et al. (1999), this region corresponds to the third α-helix in the PAI subdomain, followed by the linker and four amino acid residues of the fourth α-helix in the RED subdomain (Fig. 4). The central part of the linker region contacts the minor DNA groove according to crystallographic analysis (Xu et al., 1999). Within this relatively broader NLS activity region, L57 to S65 and I66 to S74 showed higher NLS activities (Fig. 4). This consecutive region corresponds to the C-terminal half of the third α-helix (L57 to T63) and 11 amino acid residues (G64 to S74) of the linker, which consists of a total of 16 amino acid residues (G64 to A79). We have not hitherto been able to detect a typical NLS (Kosugi et al., 2009) in this region, although positively charged amino acids needed to form a NLS are distributed within it.
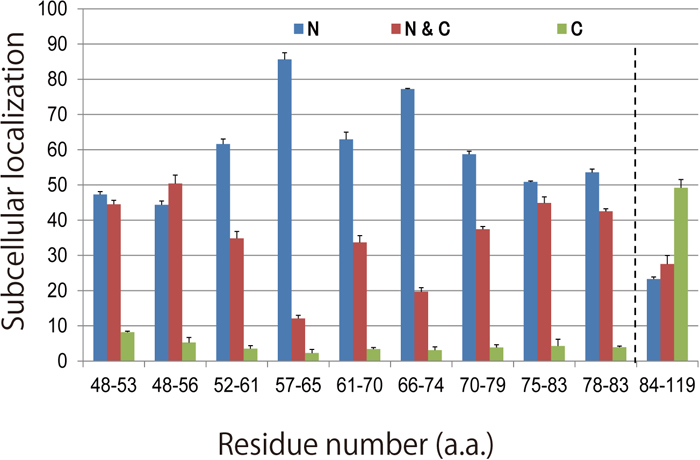
In the well-known NLS for the HD, the N-terminal amino acid segment LKRKLQR (HNLS5) adjacent to the HD and the Karyopherin 13 (importin 13)-binding site of the HD are required for the import of Pax6 and other paired-type homeodomain proteins into nuclei (Ploski et al., 2004). In contrast to the short HNLS5 with lower NLS activity, our results suggest that the two consecutive amino acid sequences (L57 to S65 and I66 to S74) with higher NLS activities and the surrounding amino acid residues with moderate NLS activities, together comprising the PD-NLS of Pax6, result in a higher activity of the NLS in the PD than that of HNLS5. We note that there may be two distinguishable ‘elements’ responsible for the higher PD-NLS activity between L57 and S74, because Y61 to A70 showed lower NLS activity (Fig. 3). In any case, because the range of highly active PD-NLS has been narrowed down in our study, it is now possible to further elucidate the molecular mechanism(s) by which the PD-NLS contributes to the nuclear localization of Pax6, by, for example, identifying where Karyopherin 13 binds and whether a nuclear import adaptor protein is required.
In summary, the NLS in the PD of chick Pax6 has been clarified in ovo to have a structure consisting of a relatively long amino acid sequence (36 residues, V48 through V83). This region spans the C- and N-terminal amino acid sequence of the PAI and RED subdomains in the PD, respectively, and the linker region between these subdomains (shown schematically in Fig. 4). Within this long NLS motif, the central area of 18 amino acids consisting of two consecutive nine-amino acid segments showed higher NLS activity; this area corresponds to the C-terminal half of the third α–helix of the PAI subdomain and the subsequent 11/16 amino acid residues of the linker between these subdomains (between the third and fourth α–helices). Therefore, when Pax6 is transported from the cytoplasm to the nucleus, it is possible that the PD-NLS works as a stronger NLS than the HD-NLS. Moreover, according to crystallographic analysis (Xu et al., 1999), it plays an important role to form the third α-helix of the PAI subdomain that binds to the major groove of the DNA double helix, and also consists of most of the linker between the PAI and RED subdomains that lie in the minor DNA groove and extensively contact the DNA.
Finally, concerning the 18 amino acid residues with high NLS activity, it should be noted that seven missense mutations of human PAX6 (T63P, G64V, I66N, P68S, G72S, G73D and S74G) associated with congenital eye diseases, such as aniridia, cataract, glaucoma, nystagmus, strabismus, morning glory disc anomaly and foveal hypoplasia, have been mapped within the corresponding human PD-NLS region (L57 to S65 and I66 to S74) of Pax6 (http://lsdb.hgu.mrc.ac.uk/variants.php?action=search_unique&select_db=PAX6). These amino acid residues are conserved in the PD-NLS of chick Pax6 and most of them reside in the linker region between the PAI and RED subdomains. Among these missense mutations, Azuma et al. (2003) reported that the P68S mutant human PAX6 protein (PAX6-P68S) failed to repress the promoter of PAX2, one of the target genes of PAX6. In addition, Chao et al. (2003) reported that the PAX6-G73D missense mutant protein possessed significantly reduced transcriptional activation ability. It will be intriguing to observe the efficiency of nuclear localization of these PAX6s. Interestingly, these seven amino acid residues are conserved in Pax1 and Pax9, both of which are devoid of the HD (Paixão-Côrtes et al., 2015). Therefore, it is possible that these seven residues also play, at least partly, an important functional role in these HD-deficient Pax proteins. These reports and our present results suggest the functional importance of the PD-NLS for Pax proteins to exert their biological functions.
Our results help us to further understand the molecular mechanism involved in ontogeny, where Pax6 plays a pivotal role.
