Abstract
To predict the seasonal vegetative growth of potted seedlings of Satsuma mandarin, the effects of initial plant conditions (age, shoot pruning), environment (greenhouse, greenhouse + shading, open-field culture), and pot size on growth were researched for 1.5–2.5 years. The growth pattern was evaluated using a curve fitting with the 4-parameter logistic (4L) model, biomass allocation, and classical (interval) growth analysis, and a generalized linear model analysis based on thermal time (tt). Growth delays from shoot pruning were confirmed by growth trajectories with the 4L model based on tt. Plant growth was positively affected by enlarging the pot size (from 20-L to 45-L), while shading significantly suppressed the growth of 45-L potted Satsuma mandarin seedlings in the greenhouse. In the growth analysis, the relative growth rate was not always determined by the net assimilation rate (NAR), which included both shoot-pruning and pot size effects. To predict the plant mass of Satsuma mandarin seedlings (MP), we proposed several generalized linear models using a log link function assuming that MP followed a Gamma-distribution. The best model to predict MP was selected based on Akaike information criterion (AIC) values, and contained explanatory variables for initial plant biomass, NAR, specific leaf area (SLA), leaf mass ratio (LMR), pot size, and tt. Some simpler models excluding NAR, but including SLA and/or LMR as explanatory variables, were more useful than a model lacking growth analysis parameters (NAR, SLA, and LMR).
Introduction
In Japan, most Satsuma mandarin trees growing in greenhouses are more than 20 years old. Citrus growers face a number of problems, including low fruit yield caused by weak tree vigor and/or by inferior light-intercepting characteristics of trees, and unstable income caused by fluctuations in fuel prices. To address these problems, a new cultivation method that shortens the vegetative growth period from planting to cropping is required for Satsuma mandarin.
Citrus growers commonly use an irrational practice to control and manipulate the vegetative growth of seedlings. Traditionally, many citrus growers prune the shoots of seedlings at transplanting to prevent water deficit and/or to decrease the number of flower buds in an open field. However, as pointed out in a few reports, unpruned trees show maximum growth (Alexander and Maggs, 1970, 1971). Unlike primitive open field culture, modern horticultural techniques have made it possible to irrigate large orchards and control flowering using gibberellin (Takahara et al., 1990). Therefore, the effect of shoot pruning at transplanting on plant growth must be evaluated objectively using a quantitative approach such as growth analysis (Hunt, 1982), and the growing practice should be rationalized to minimize the vegetative growth period from planting to cropping.
Growing seedlings in pots temporarily (for one or two years) before transplanting them into an orchard is one effective technique for shortening the vegetative growth period of fruit trees from planting to cropping (Iguchi et al., 1985). The advantages of this potting technique are that it is easy to move the plants and saves on the labor of digging plants out of the soil. However, as shown by many previous reports, pot size strongly restricts plant growth (Poorter et al., 2012a) by limiting nutrient availability (Bar-yosef et al., 1988; Mark and Marra, 1994; Yu et al., 2012) and leaf stomatal conductance (Ronchi et al., 2006). From a similar viewpoint, root restriction has been proposed to control vigorous vegetative growth of fruit trees (Hsu et al., 1996; Imai et al., 1987) and fruit quality (Mataa and Tominaga, 1998; Taniguchi, 1993). In this way, suppression of vegetative growth is inevitable for pot-grown plants because of their restricted root growth, but it is important to know the optimal pot size for new cultivation methods (e.g. high-density planting), and to predict the quantitative growth of seedlings in a given environment and field space.
The goal of this study was to predict the seasonal vegetative growth of potted Satsuma mandarin seedlings based on initial plant conditions (age, shoot pruning), environment, and pot size. Under equal pot-size conditions, a previous study indicated that the natural-log-transformed plant mass of Satsuma mandarin seedlings (mP) fitted a 4-parameter logistic (4L) model on a tt basis (Yano et al., 2018). However, even under equal tt conditions, different environmental conditions (e.g. light condition) and/or pot size will change the 4L coefficients; thus, the usefulness of a 4L model is limited to each specific case. Conversely, a general and progressive 4L model may more complicated. Here, to construct a simple prediction model for the growth of Satsuma mandarin seedlings, we first confirmed the effects of the initial plant conditions and pot size on their growth using a 4L model, biomass allocation, and growth analysis on a tt basis. Next, we tested several multiple regression models to predict plant mass not only by these variables (initial plant size, pot size, and environmental data as tt), but also growth analysis parameters. We used a generalized linear model with a log link function, assuming that plant mass followed a Gamma-distribution. Additionally, in this study, “growth” indicates vegetative growth.
Materials and Methods
Plant materials
In the experiment on the effect of shoot pruning, 1-year-old and 2-year-old ‘Miyagawa-Wase’ Satsuma mandarin seedlings grafted onto trifoliate orange [Poncirus trifoliata (L.) Raf.] rootstocks were planted in 15-L pots on March 25, 2014 and cultivated until August 2, 2015 in a field of the Fruit Tree Group, Oita Prefectural Agriculture, Forestry, and Fisheries Research Center (Kunisaki, Oita, Japan). Shoot pruning was performed on 2-year-old ‘Miyagawa-Wase’ Satsuma mandarin seedlings as shown in Fig. 1. The dry matter removed by pruning was 79% of the shoot (48% of the plant) compared with unpruned 2-year-old seedlings.
In the experiment on the effect of pot size, 1-year-old ‘Oita-Wase’ Satsuma mandarin seedlings grafted onto trifoliate orange rootstocks were planted in 20-L pots on March 29, 2013 and cultivated until October 5, 2015 in the field as described above. Transplanting from 20-L pots to 45-L pots was conducted on April 15, 2014.
In previous reports, the tree vigor of ‘Miyagawa-Wase’ was in between extreme early maturing Satsuma mandarins and late maturing Satsuma mandarins (Takahara, 2004). Similarly, the tree vigor of ‘Oita-Wase’ (nucellar seedlings of extreme early maturing Satsuma mandarins) was weaker than late maturing Satsuma mandarins, and was relatively strong among extreme early maturing Satsuma mandarins (Kawano, 2004). Therefore, during the vegetative growth period, we assumed that the difference in growth patterns between ‘Miyagawa-Wase’ and ‘Oita-Wase’ was negligible, and we defined both the ‘Miyagawa-Wase’ and ‘Oita-Wase’ seedlings as Satsuma mandarin seedlings.
Irrigation was supplied to the pots via a drip tube every day: 0.6 L was supplied to 15-L and 20-L pots, and 1.8 L was supplied to 45-L pots. Each Satsuma mandarin seedling was fertilized with 200 g coated granular fertilizer (N:P:K = 14:11:13) each year. All flower buds were detached during pre-anthesis. The sampling intervals were from 6 months to 1 year, with five replicates for each experiment (n = 5). Roots were all less than 2 cm in diameter.
Environmental conditions and measurements
From April 23, 2013 until the final sampling, the plants were grown under three different environmental conditions: (1) greenhouse; in an unheated greenhouse to promote the growth of Satsuma mandarin seedlings, (2) greenhouse + shading; in an unheated greenhouse under low light intensity (from shading), and (3) open field; in an open field. The greenhouse ceiling consisted of 0.15-mm polyester film, and the shade cloth was a knitted silver polyvinyl film.
Air temperature (Ta) was measured with a compact water proof temperature data logger (RTR-53, T&D Co., LTD., Japan) and solar radiation (Rs) was measured with a pyranometer (Solar Mini PCM-01, Prede Co., LTD., Japan) at 10-min intervals.
Estimating growth trajectories of shoot-pruned and unpruned Satsuma mandarin seedlings
We assumed that the mP of unpruned Satsuma mandarin seedlings would fit 4L functions as growth trajectories on a tt basis (Yano et al., 2018). In the fitting, the initial mP and tt for unpruned 2-year-old Satsuma mandarin seedlings were replaced as follows; tt in greenhouse was estimated by the initial linear relationship between mP (4.2–5.6) and tt (0–5,056 day·°C) of unpruned 1-year-old Satsuma mandarin seedlings (Fig. 3a), and tt in an open field was estimated using AMEDAS data for Asakura (Fukuoka, Japan), the nearest city to the field where the Satsuma mandarin seedlings were cultivated. Similarly, the initial biological tt (tt′) for shoot-pruned 2-year-old Satsuma mandarin seedlings was determined from the 4L parameters as follows:
|
tt'=-1rlnm0K-m0'(K-L-m0)(m0'-L) |
where m
0 is the natural-log-transformed initial biomass of unpruned 1-year-old Satsuma mandarin seedlings, K is the upper horizontal asymptote of 4L from unpruned 1-year-old and 2-year-old Satsuma mandarin seedlings, m
0′ is the natural-log-transformed initial biomass of shoot-pruned 2-year-old Satsuma mandarin seedlings, L is the lower horizontal asymptote of 4L from unpruned 1-year-old and 2-year-old Satsuma mandarin seedlings, and r is a constant determining the curvature of the growth pattern in 4L from unpruned 1-year-old and 2-year-old Satsuma mandarin seedlings.
Growth analysis by a classical (interval) approach
The growth analysis by the classical approach followed Hunt (1982). In this approach, the mean relative growth rate (RGR_) on a tt basis was calculated as follows:
where
mP2 and
mP1 are the natural-log-transformed plant masses of Satsuma mandarin seedlings at
tt2 and
tt1, respectively. Similarly, the derivative parameters net assimilation rate (
NAR_), specific leaf area (
SLA_), and leaf mass ratio (
LMR_) were calculated directly from the experimental data as follows:
|
NAR_=MP2-MP1tt2-tt1×logeAL2-logeAL1AL2-AL1
SLA_=AL1/ML1+AL2/ML22
LMR_=ML1/MP1+ML2/MP22
|
where M
P2 and M
P1 are the non-transformed plant masses of Satsuma mandarin seedlings, A
L2 and A
L1 are the plant leaf areas, and M
L2 and M
L1 are the plant leaf masses of Satsuma mandarin seedlings at
tt2 and
tt1, respectively.
Generalized linear model
Pooled sampling data (n = 37, shown in a supplemental data, Table S1) were tested for a generalized linear model. Each measurement value was derived from five trees (n = 5). Data analysis was performed using R-3.3.2 (R Core Team, 2016).
Results
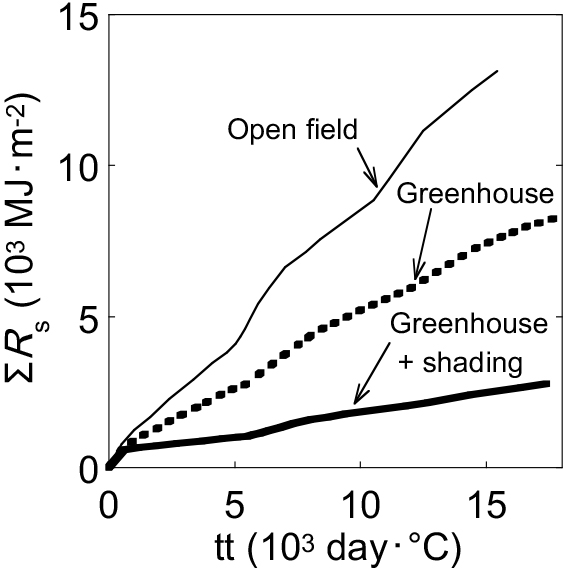
Figure 2 shows the seasonal changes in the integrated solar radiation (ΣRs) on a tt basis during the experimental period. At the same tt level, the ΣRs values of greenhouse and greenhouse + shading were almost 60% and 20% those of an open field, respectively (Fig. 2).
Growth trajectories of unpruned and shoot-pruned Satsuma mandarin seedlings
In evaluating the effects of initial plant conditions (age and shoot pruning) on the growth process of potted Satsuma mandarin seedlings, it was assumed that the mP fitted a 4L model on a tt basis (Yano et al., 2018). Both in a greenhouse and an open field, the growth patterns of unpruned potted Satsuma mandarin seedlings, including both 1-year-old Satsuma mandarin seedlings (1y) and 2-year-old Satsuma mandarin seedlings (2y), followed this assumption (Fig. 3, P < 0.001). However, the mP of shoot-pruned 2-year-old Satsuma mandarin seedlings (p2y) showed different growth trajectories, with trends below the 4L line estimated from the unpruned mP (Fig. 3).
Biomass allocation—environment and initial plant conditions
To understand the effects of initial plant conditions and the environment on the growth of Satsuma mandarin seedlings, growth parameters and mass distribution were compared at final sampling (August 2, 2015). The results of two-way ANOVA indicated that the environment significantly affected the tuber girth (TG), leaf mass (ML), stem mass (MS), root mass (MR), plant mass (MP), and leaf area of Satsuma mandarin seedlings (AL), and that initial plant conditions significantly affected ML, MS, MR, MP, and AL, and that there were no significant interactions of environment × initial plant conditions except for MR (Table 1).
Growth analysis—environment and initial plant conditions
In growth analysis based on tt, in both a greenhouse and an open field, the initial RGR_ and LMR_ values of shoot-pruned Satsuma mandarin seedlings were lower than those of unpruned Satsuma mandarin seedlings (Fig. 4a, b, g, h), but the initial NAR of shoot-pruned Satsuma mandarin seedlings were higher than that of unpruned Satsuma mandarin seedlings (Fig. 4c, d).
Biomass allocation—environment and pot size
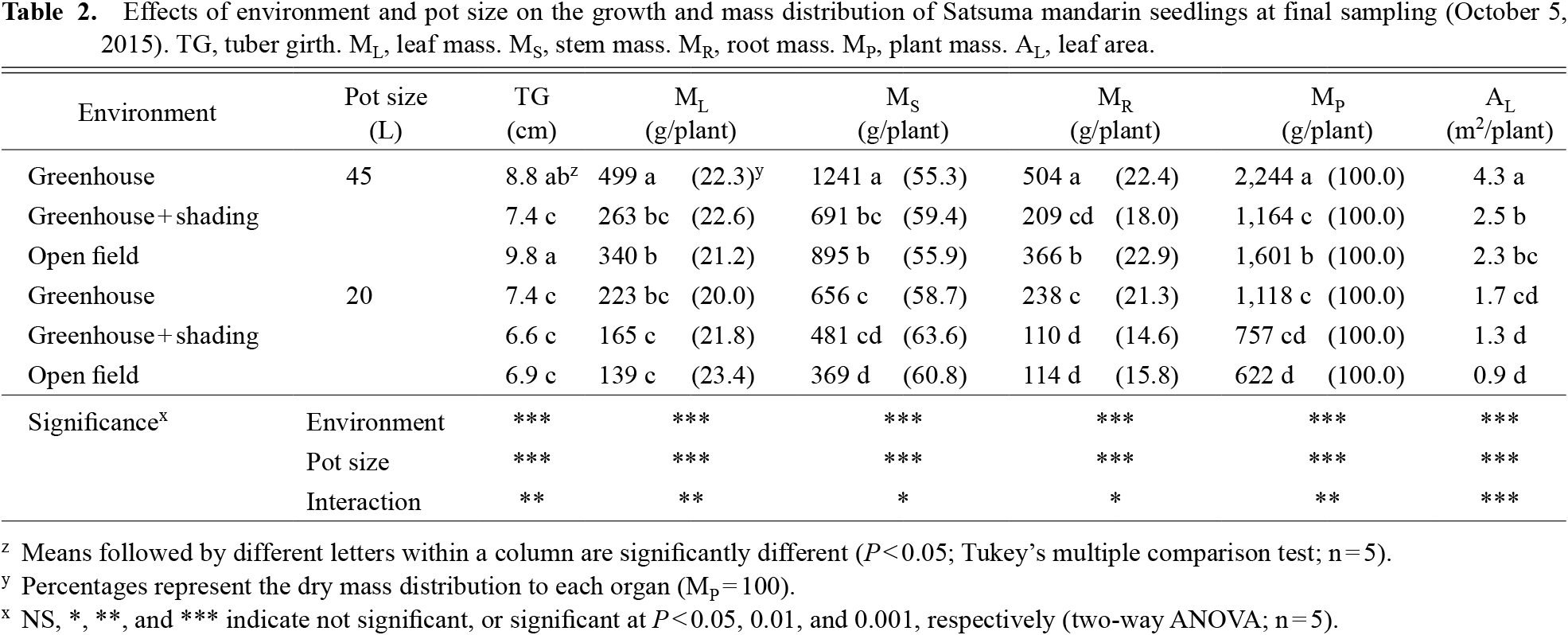
After transplanting, the mP values of plants transplanted into 45-L pots in a greenhouse and an open field were significantly larger than those of plants in 20-L pots in the same environments (Fig. 5a, c). The mP of plants transplanted into 45-L pots in the shaded greenhouse was not significantly different from the mP of plants in 20-L pots in greenhouse + shading (Fig. 5b). Two-way ANOVA of data collected at the final sampling (October 5, 2015) showed that both environment and pot size significantly affected the growth parameters of TG, ML, MS, MR, MP, and AL, and there were significant interactions of environment × pot size for them (Table 2). In all treatments, biomass allocations to the leaves showed relatively small differences (under 4%), and under equal pot-size conditions, shading tended to increase biomass allocation to the stem and decrease it to the roots (Table 2).
Growth analysis—environment and pot size
Generally, in the growth analysis, RGR_ and NAR_ showed similar patterns of change during both the first season after transplanting (from April 15, 2014 to November 14, 2014) and the second season after transplanting (from November 14, 2014 to August 2, 2015) (Fig. 6a, b). The RGR_ and NAR_ values were always larger for 45-L than 20-L pots under all three environments (Fig. 6a, b). However, unlike RGR_ and NAR_, no significant differences were observed between 45-L and 20-L pots in SLA_ and LMR_ under all three environments during either the first or second season (Fig. 6c, d).
Generalized linear model for MP based on growth components and environmental factors
To predict the seasonal growth of Satsuma mandarin seedlings, we estimated a generalized linear model for MP using growth components and environmental factors under the assumption that MP followed a Gamma-distribution. The pooled raw data of an initial plant biomass (M0) included not only the initial values for 1y, 2y, and p2y (Fig. 3a, b), but also the initial MP values just before transplanting (Fig. 5a–c). Selected models for MP containing the parameters M0, pot size, and tt showed a significant positive contribution (P < 0.0001) (Table 3). Significant negative correlations were detected between NAR_ and LMR_ (r = −0.78, P < 0.0001), and between SLA_ and ΣRs (r = −0.61, P < 0.0001). A stepwise AIC method was used to assess the model parameters, excluding ΣRs (Table 3). As shown in Model 1, adding NAR_, SLA_, and/or LMR_ as explanatory variables to Model 5 reduced the AIC (Table 3).
For example, in Model 1 MP could be estimated by the following equation:
where, mp1 is the value estimated by the following equation:
|
mp1=0.58+6.37×10−3M0+5.50NAR_+9.90SLA_
+8.74LMR_+1.57×10−2(Pot size)
+1.45×10−4tt.
|
Discussion
Poorter et al. (2012a) suggested that an appropriate pot size to avoid growth suppression is one in which the plant biomass does not exceed 1 g·L−1 (total plant biomass:pot size) from a meta-analysis including herbaceous and woody plants. In this study, the minimum and maximum values of total plant biomass of Satsuma mandarin seedlings:pot size ratios were 4 g·L−1 (tt = 0 day·°C) and 56 g·L−1 (tt = 17,698 day·°C). Therefore, our data were obtained under conditions in which considerable suppression of growth by pot size would have occurred as growth progressed.
Biomass allocation for active growth in potted Satsuma mandarin seedlings
Analysis of biomass allocation in the Satsuma mandarin seedlings indicated three characteristics: first, the shoot/root imbalance caused by shoot pruning was altered at the latest by the next season. Temporal decreases in LMR caused by shoot pruning occurred within the first season in a greenhouse (Fig. 4g, tt1′ = 2,831 day·°C, tt2′ = 7,419 day·°C) and an open field (Fig. 4h, tt1′ = 2,760 day·°C, tt2′ = 7,184 day·°C), and afterwards, the LMR of pruned Satsuma mandarin seedlings increased as these of unpruned Satsuma mandarin seedlings both in a greenhouse and an open field (Fig. 4g, h). In addition, at final sampling, not only the environment, but also the initial plant condition (size) significantly affected the leaf mass, stem mass, root mass, whole plant mass, and leaf area (Table 1). These results suggested that even though shoot pruning in Satsuma mandarin seedlings temporally affected the biomass allocation to the shoot and the roots, the initial plant size significantly affected the plant growth at least under the same environment and pot size.
The second characteristic was that biomass allocation to the leaves and leaf expansion were dominant regardless of the environment or pot size. Shading in the greenhouse resulted in small changes in biomass allocation to the leaves; those in 45-L and 20-L pots were only 0.3% and 1.8% (Table 2). Comparing among a greenhouse, greenhouse + shading, and an open field, LMR also showed a similar result (Fig. 6d). In contrast, environment and pot size tended to affect the biomass allocation to the stem and the roots. With shading in the greenhouse, the increases in biomass allocation to the stem in 45-L and 20-L pots were 4.1% and 4.9%, and the decreases in allocation to the roots in 45-L and 20-L pots were 4.4% and 6.7%, respectively (Table 2).
The third characteristic was that photosynthetic restriction predominantly suppressed root growth. Comparing a greenhouse and a greenhouse + shading, as indicated above, at both pot sizes (45-L and 20-L), a significant decrease was only observed in MR by shading (Table 2).
These characteristics suggest that even under low light conditions, a suitable temperature and large pot size could activate plant growth through the coordination of the shoot/root balance and the dominance of leaf growth at the expense of other organs in Satsuma mandarin seedlings.
Growth parameter determining RGR and MP
Growth is typically characterized by RGR; therefore, the contributions of its derivatives including NAR, SLA, and LMR have been studied for many plants (Shipley, 2002, 2006; Rees et al, 2010). Based on the results of many studies, including interspecific analysis of herbaceous and woody plants, RGR can be classified as SLA-dominant (Gibert et al., 2016; Lambers and Poorter, 1992; Poorter and Remkes, 1990; Rees et al., 2010; Reich et al., 1992, 1998; Walters et al., 1993; Wright and Westoby, 1999, 2000) or NAR-dominant (Huante and Rincon, 1998; Huante et al., 1995; Poorter, 1999; Ryser and Wahl, 2001; Saverimuttu and Westoby, 1996; Shipley, 2006; Taub, 2002; Yano et al., 2018), while LMR barely contributes to RGR (Shipley, 2006).
Essentially, the pot size effect must be similar to the grafting effect to control the shoot vigor of the rootstock. Under this assumption, the effects of both grafting and pot size on plant growth can be explained by variances in leaf photosynthesis, stomatal conductance, and hydraulic conductance originating from root growth (Iwasaki et al., 2011; Poorter et al., 2012a; Taylaran et al., 2011). This partly supports our result that enlarging the pot size increased both RGR and NAR (Fig. 6a, b). A meta-analysis also showed similar results; i.e., that pot size effects on plant growth were caused mainly by NAR rather than SLA or LMR (Poorter et al., 2012a). In this study, however, we detected some differences in the contributions of LMR and NAR to RGR between shoot pruning and enlargement of pot size. Shoot pruning reduced both initial RGR (Fig. 4a, b) and initial LMR (Fig. 4g, h), but increased initial NAR (Fig. 4c, d). The NAR increase may simply compensate for the LMR decrease, as a few studies reported that shoot pruning did not positively affect photosynthesis (Morais et al., 2011). However, other studies reported an increase in NAR or leaf photosynthesis after shoot pruning or cutting off an organ (Iglesias et al., 2002; Maggs, 1964; Thorne and Evans, 1964). In any case, an absolute decrease in the source capacity of a plant can never increase the RGR.
Only a few studies have reported a correlation between RGR and LMR (Shipley, 2006; Tomlinson et al., 2014). In a comparison of deciduous and evergreen savanna trees, deciduous trees showed a wide range of LMR values and a significant positive correlation between RGR and LMR, while evergreen trees did not (Tomlinson et al., 2014). Meta-analyses have shown that the natural variance in LMR must be smaller than the variance in SLA in evergreen trees (Poorter et al., 2012b). However, an artificial LMR decrease (i.e., shoot pruning) altered the maximum growth of sweet orange seedlings (Alexander and Maggs, 1971) and reduced the initial RGR of Satsuma mandarin seedlings in this study (Fig. 4a, b).
Multiple regression models to predict plant mass
Our results indicate that RGR is not always determined only by NAR or SLA, and in some cases, is possibly affected by LMR. Additionally, in the best generalized linear model (as selected based on the AIC), the growth parameters were all derivatives of RGR, and the components included initial plant size (M0), NAR, SLA, LMR, pot size, and tt (Table 3). A similar result was reported for seedlings of temperate tree species (Modrzynski et al., 2015).
As mentioned in the results, among the explanatory variables for MP, NAR and LMR were significantly negatively correlated (P < 0.0001) and SLA and integrated solar radiation (ΣRS) were significantly positively correlated (P < 0.0001) in this study. To construct a simpler model for MP and avoid multicollinearity, based on the former relationship, NAR can be excluded as an explanatory variable because LMR was the key parameter indicating the level of shoot pruning (Fig. 4; Table 3). In addition, NAR is difficult to measure because it requires the dry mass of the tree, including the roots. In the latter relationship, ΣRS was already excluded by the stepwise AIC, and SLA has been shown to be a suitable and precise parameter representing light conditions (Yano et al., 2018). Therefore, three of the short models [Model 2, consisting of Model 1 without NAR; Model 3, consisting of Model 1 without NAR and SLA; and Model 4, consisting of Model 1 without NAR and LMR] were not the best models, but would be useful for predicting growth because the models that included SLA and/or LMR as explanatory variables reduced the AIC as compared with Model 5 (Table 3).
Conclusion
The growth of potted Satsuma mandarin seedlings was regulated not only by initial plant size, but also by the shoot/root balance, as shown by LMR. Enlargement of pot size positively affected growth, while shading significantly suppressed the growth of 45-L potted Satsuma mandarin seedlings in a greenhouse. To predict the seasonal vegetative growth of Satsuma mandarin seedlings, we proposed several simple multiple regression models that considered shoot pruning and altered pot size, in which growth was primarily regulated by initial plant mass, pot size, tt, and growth analysis parameters such as SLA and/or LMR. Under these proposed models, a limited environmental resource such as a lower temperature under open field conditions than in a greenhouse or limited field space could be compensated for by initial plant size, enlarging the pot size, or prolonging the growth period. These proposed models should help to establish a new cultivation method for high-yield performance in greenhouse-grown Satsuma mandarins.
Literature Cited
- Alexander, D. M. and D. H. Maggs. 1970. Effects of initial size and of shading on the quantitative growth of seedlings of the citrus rootstock Carrizo sitrange. Ann. Bot. 1970: 67–73.
- Alexander, D. M. and D. H. Maggs. 1971. Growth responses of sweet orange seedlings to shoot and root pruning. Ann. Bot. 1971: 109–115.
- Bar-yosef, B., S. Schwartz, T. Markovich, B. Lucas and R. Assaf. 1988. Effect of root volume and nitrate solution concentration on growth, fruit yield, and temporal N and water uptake rates by apple trees. Plant and Soil 107: 49–56.
- Gibert, A., E. F. Gray, M. Westoby, I. J. Wright and D. S. Falster. 2016. On the link between functional traits and growth rate: meta-analysis shows effects change with plant size, as predicted. J. Ecol. 104: 1488–1503.
- Hsu, Y. M., M. J. Tseng and C. H. Lin. 1996. Container volume affects growth and development of wax-apple. HortScience 31: 1139–1142.
- Huante, P. and E. Rincon. 1998. Response to light changes in tropical deciduous woody seedlings with contrasting growth rates. Oecologia 113: 53–66.
- Huante, P., E. Rincon and I. Acosta. 1995. Nutrient availability and growth rate of 34 species from a tropical deciduous forest in Mexico. Funct. Ecol. 9: 849–858.
- Hunt, R. 1982. Plant growth curves: The functional approach to plant growth analysis. Edward Arnold, London.
- Iglesias, D. J., I. Lliso, F. R. Tadeo and M. Talon. 2002. Regulation of photosynthesis through source: sink imbalance in citrus is mediated by carbohydrate content in leaves. Physiol. Plant. 116: 563–572.
- Iguchi, I., T. Suzuki, K. Mochizuki and M. Konakahara. 1985. Improvement in rapid growing methods for nursery stock of citrus. (2) Grafting time, growth environments and transplanting time. Bull. Shizuoka Citrus Exp. Stn. 21: 19–25 (In Japanese with English abstract).
- Imai, S., G. Okamoto and M. Endo. 1987. Effects of dense planting and root-system control on attaining greater early production and fruit stability of tetraploid grapes. Bull. Hiroshima Fruit Tree Expt. Stn. 12: 1–9 (In Japanese with English abstract).
- Iwasaki, M., H. Fukamachi, K. Satoh, H. Nesumi and T. Yoshioka. 2011. Development of tree vigor prediction method at an early stage based on stem hydraulic conductance of seedlings in citrus rootstocks. J. Japan. Soc. Hort. Sci. 80: 390–395.
- Kawano, T. 2004. Citrus. The basics. Cultivation properties of Satsuma mandarins. p. 84. In: Nogyo-gijutsu-taikei (In Japanese) Fruit tree. No-san-gyoson-bunka-kyokai, Tokyo.
- Lambers, H. and H. Poorter. 1992. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv. Ecol. Res. 23: 187–261.
- Maggs, D. H. 1964. Growth-rates in relation to assimilate supply and demand I. leaves and roots as limiting regions. J. Exp. Bot. 15: 574–583.
- Mark, R. and F. Marra. 1994. Responses of young peach trees to root confinement. J. Amer. Soc. Hort. Sci. 119: 223–228.
- Mataa, M. and S. Tominaga. 1998. Effects of root restriction on tree development in ponkan mandarin (Citrus Reticulata Blanco). J. Amer. Hort. Sci. 123: 651–655.
- Modrzynski, J., D. J. Chmura and M. G. Tjoelker. 2015. Seedling growth and biomass allocation in relation to leaf habit and shade tolerance among 10 temperate tree species. Tree Physiol. 35: 879–893.
- Morais, L. E., P. C. Cavatte, E. F. Medina, P. E. M. Silva, S. C. V. Martins, P. S. Volpi, S. Andrade Junior, J. A. Machado, C. P. Ronchi and F. M. Damatta. 2011. The effect of pruning at different times on the growth, photosynthesis and yield of conilon coffee (coffea canephora) clones with varying patterns of fruit maturation in southeastern Brazil. Expl. Agric. 48: 210–221.
- Poorter, L. 1999. Growth responses of 15 rainforest tree species to a light gradient: the relative importance of morphological and physiological traits. Funct. Ecol. 13: 396–410.
- Poorter, H. and C. Remkes. 1990. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 83: 553–559.
- Poorter, H., J. Bühler, D. Dusschoten, J. Climent and J. A. Postma. 2012a. Pot size matters: a meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 39: 839–850.
- Poorter, H., K. J. Niklas, P. B. Reich, J. Oleksyn, P. Poot and L. Mommer. 2012b. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist 193: 30–50.
- R Core team. 2016. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/.
- Rees, M., C. P. Osborne, F. I. Woodward, S. P. Hulme, L. A. Turnbull and S. H. Taylor. 2010. Partitioning the components of relative growth rate: how important is plant size variation? Am. Nat. 176: E152–E161.
- Reich, P. B., M. G. Tjoelker, M. B. Walters, D. W. Vanderklein and C. Buschena. 1998. Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedling of nine boreal tree species grown in high and low light. Funct. Ecol. 12: 327–338.
- Reich, P. B., M. B. Walters and D. S. Ellsworth. 1992. Leaf lifespan in relation to leaf plant and stand characteristics among diverse ecosystem. Ecol. Monogr. 62: 365–392.
- Ronchi, C. P., F. M. DaMatta, K. D. Batista, G. A. B. K. Moraes, M. E. Loureiro and C. Ducatti. 2006. Growth and photosynthetic down-regulation in Coffea arabica in response to restricted root volume. Funct. Plant Biol. 33: 1013–1023.
- Ryser, P. and S. Wahl. 2001. Interspecific variation in RGR and underlying traits among 24 grass species grown in full daylight. Plant Biol. 3: 426–436.
- Saverimuttu, T. and M. Westby. 1996. Components of variation in seedling relative growth rate: phylogenetically independent contrasts. Oecologia 105: 281–285.
- Shipley, B. 2002. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: the relationship with daily irradiance. Funct. Ecol. 16: 682–689.
- Shipley, B. 2006. Net assimilation rate, specific leaf area and leaf mass ratio: which is most closely correlated with relative growth rate? A meta-analysis. Funct. Ecol. 20: 565–574.
- Takahara, T. 2004. Citrus. The basics. Cultivation properties of Satsuma mandarins. p. 88. In: Nogyo-gijutsu-taikei (In Japanese) Fruit tree. No-san-gyoson-bunka-kyokai, Tokyo.
- Takahara, T., K. Hirose, I. Iwagaki and S. Ono. 1990. Enhancement of suppression effect of flower-bud formation of citrus varieties by addition of machine oil emulsion to gibberellin. Bull. Fruit Tree Res. Stn. 18: 77–89 (In Japanese with English abstract).
- Taniguchi, T. 1993. Rooting-zone restriction for Satsuma mandarin. Agric. Hort. 68: 490–496 (In Japanese).
- Taub, D. R. 2002. Analysis of interspecific variation in plant growth responses to nitrogen. Can. J. Bot. 80: 34–41.
- Taylaran, R. D., S. Adachi, T. Ookawa, H. Usuda and T. Hirasawa. 2011. Hydraulic conductance as well as nitrogen accumulation plays a role in the higher rate of leaf photosynthesis of the most productive variety of rice in Japan. J. Exp. Bot. 62: 4067–4077.
- Thorne, G. N. and A. F. Evans. 1964. Influence of tops and roots on net assimilation rate of sugar-beet and spinach beet and grafts between them. Ann. Bot. 28: 499–508.
- Tomlinson, K. W., L. Poorter, F. Bongers, F. Borghetti, L. Jacobs and F. Van Langevelde. 2014. Relative growth rate variation of evergreen and deciduous savanna tree species is driven by different traits. Ann. Bot. 114: 315–324.
- Walters, M. B., E. L. Kruger and P. B. Reich. 1993. Growth, biomass distribution and CO2 exchange of northern hardwood seedlings in high and low light: relationships with successional status and shade tolerance. Oecologia 94: 7–16.
- Wright, I. J. and M. Westoby. 1999. Differences in seedling growth behavior among species: trait correlations across species, and trait shifts along nutrient compared to rainfall gradients. J. Ecol. 87: 85–97.
- Wright, I. J. and M. Westoby. 2000. Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs leaf function in 28 Australian woody species. Funct. Ecol. 14: 97–107.
- Yano, T., A. Morisaki, K. Matsubara, S. Ito and M. Kitano. 2018. Growth analysis of potted seedlings of Satsuma mandarin (Citrus unshiu Marc.) under different light conditions and air temperatures. Hort. J. 87: 34–42.
- Yu, X., B. Wang, C. Zhang, W. Xu, J. He, L. Zhu and S. Wang. 2012. Effect of root restriction on nitrogen levels and glutamine synthetase activity in ‘Kyoho’ grapevines. Sci. Hort. 137: 156–163.