原著論文
Blue LED Irradiation Induces Scoparone Production in Wounded Satsuma Mandarin ‘Aoshima Unshu’ and Reduces Fruit Decay during Long-term Storage
2018 年 87 巻 4 号 p. 474-480

詳細
2018 年 87 巻 4 号 p. 474-480
In the present study, we examined the effect of blue LED irradiation on production of phytoalexin scoparone in wounded satsuma mandarin fruit as a result of fungal infections, as well as the effect of tape-type blue LED on fruit decay during long-term storage (from December to April) in an actual storage room. The blue LED treatment reduced the rate of fruit decay compared to that of dark treatment in wounded fruits. The decay rate of LED irradiated fruit was 13.3%, while that of non-irradiated fruit was 51.1%. In wounded fruits, blue LED treatment resulted in significantly higher scoparone contents than dark treatment. Intact (non-wounded) fruits had low scoparone contents, irrespective of blue LED irradiation. In an examination of changes in total fruit decay rate during storage using tape type-blue LED, the total decay rate was not significantly different between the LED-treated and untreated fruits until 64 days after storage (36 days after starting LED irradiation). On the other hand, at 92 and 120 days after the start of storage, total fruit decay under blue LED treatment was significantly lower compared to the untreated control. The tape-type blue LED (10 μmol·m−2·s−1) did not affect either the fruit quality analyzed (soluble solid content, titratable acidity, specific gravity, percentage of flesh) or the rind color. These results indicate that blue light induces scoparone production in wounded satsuma mandarin, and because of this, along with other putative factors, tape-type blue LED irradiation reduces mandarin fruit decay during long-term storage.
Citrus fruits are cultivated around the world, including East Asia. Many cultivars with different harvest periods grow in various locations. In Japan, the satsuma mandarin (Citrus unshiu Marcow.) is mainly harvested from October to December and some of the produce is stored at normal or low temperatures (5–8°C) for three months. Improvements in storage conditions and postharvest management of this fruit during storage have widely studied (Fujisawa et al., 2001; Kitajima and Yamada, 1957; Murata and Yamawaki, 1992). As a practical result of such investigations, storage conditions have been improved, and high temperature treatment before storage, chemical fungicides, and other postharvest treatments aimed at preventing fruit decay during storage are routinely used. However, long-term storage has become more difficult because of global warming, which has made rind puffing of mandarin fruits more common nowadays. Puffy fruits are frequently damaged during harvest, transport, and storage when they can be easily infected by fungi and thus quickly decay. Major postharvest citrus pathogens causing severe economic loses during storage include Penicillium italicum (blue mold), Penicillium digitatum (green mold), and Botrytis cinerea (gray mold).
Synthetic chemical fungicides are widely used for the treatment of various diseases in fresh agricultural products, including citrus cultivars. However, the use of these fungicides has become restricted because of the threat they pose to human health and the environment, as well as the risk of development of pathogen resistance to these chemicals by their misapplication, which eventually turns them useless. Therefore, it is important to develop alternative methods for reducing the application of chemical fungicides. Previous studies have suggested application of salt, sodium bicarbonate, hydrogen peroxide, and essential oils, among other compounds, to control postharvest citrus diseases (Cerioni et al., 2012, 2013; Kouassi et al., 2012; Youssef et al., 2012). However, these methods have not been introduced in Japan because drying the surface of the fruits is time consuming and costly.
On the other hand, several studies have suggested that postharvest treatment using LED, could suppress some diseases, such as those caused by P. digitatum Sacc and P. italicum (Alférez et al., 2012; Lafuente and Alférez, 2015; Liao et al., 2013). The mechanism of induction of this resistance against pathogens has been documented for some plant models. For example, some studies have suggested that the antifungal response mechanism induced by blue light to control postharvest decay of citrus fruits is associated with increased phospholipase A2 activity (Alférez et al., 2012; Liao et al., 2013). One study has shown a relationship between exposure to blue light (at a photon flux of 630 μmol·m−2·s−1) and accumulation of scoparone in sweet oranges (Citrus sinensis L. Osbeck) (Ballester and Lafuente, 2017). In addition, irradiation with blue light can directly inhibit fungal growth in satsuma mandarin fruits and induce an antifungal response against blue mold (Liao et al., 2013; Yamaga et al., 2015a). Therefore, blue LED seems to be effective alternative technology to chemical treatment for citrus fruits in storage.
However, little information is available regarding the protective mechanism afforded by blue LED irradiation on satsuma mandarin fruits, a citrus variety with a thin peel. In addition, the use of blue LED for postharvest treatment or during fruit storage is limited only to the actual production sites. There are few reports on the use of postharvest LED for satsuma mandarin fruits in storage rooms. To verify the potential of blue LED to control fruit decay, we need to evaluate the effect of treating storage containers in the storage room with blue LED. One difficulty of this approach lies in the limited coverage of mandarins by the applied irradiation because several containers are piled up in the storage room, making it hard to effectively irradiate fruits under the prevailing storage conditions. Moreover, since LEDs are costly, we must try to use the least possible number of light sources for irradiation. Previously, we reported that irradiation with blue LED light at of 8 μmol·m−2·s−1 photon flux reduced blue mold sporulation in the satsuma mandarin ‘Aoshima unshu’ (Yamaga et al., 2015b). Therefore, it should be possible to reduce citrus fruit decay caused by postharvest fungal diseases in storage rooms by irradiation with relatively low intensities of blue LED light.
The objective of our study was to investigate the production of scoparone, which shows stronger inhibitory activities against fungi than other coumarins (Kuniga and Nesumi, 2011), by the flavedo tissue of satsuma mandarin fruits under blue LED irradiation. We also examined the effects of the application of tape-type blue LED at a photon flux of 10 μmol·m−2·s−1, which can be used in a narrow space, on fruit decay in an actual storage room from December to April.
The satsuma mandarin ‘Aoshima unshu’ fruits were harvested from an orchard at Shizuoka Prefectural Agriculture and Forestry Research Institute Fruit Tree Research Center, from December 1 to December 3, 2015. Fifteen incisions 1 mm deep and 0.5 mm in diameter were made between the pedicel and the lateral part of the fruit with a needle equipped with a stopper to ensure wound uniformity (Fig. 1). The wounded fruits were maintained under irradiation with blue LED (emission peak, 465 nm; photon flux, 80 μmol·m−2·s−1; Yamato Industrial, Shizuoka, Japan) for 7 days in a chamber at 20°C and over 85% relative humidity (RH). The same conditions were used for the dark treatment, except for the absence of any light source for the duration of the experiment. After 7 days, the number of decayed fruits due to natural fungal infection (as shown by water-soaked, soft rot spots) was counted for each treatment, which consisted of three replicates of 15 fruits each, kept in a polypropylene container (L410 mm × W300 mm × H60 mm).

The part wounded on satsuma mandarin ‘Aoshima unshu’ fruits.
Harvested mandarin fruits mentioned above were used. Thirty points of 1 mm deep × 0.5 mm diameter incisions were made by the method described above. The wounded fruits were maintained under blue LED (the same as described above) irradiation for 4 days at 20°C in a chamber with over 85% RH. The same number of non-wounded fruits was irradiated under the same conditions. Dark treatment was performed for wounded and non-wounded fruits under identical temperature and humidity conditions. Thereafter, scoparone contents of the flavedo were measured. Scoparone extraction was performed as described by Kuniga et al. (2005). The flavedo tissue (0.8 g) of the harvested fruits was excised with a knife and extracted using 80% ethanol. The tissue was then homogenized in a blender and centrifuged at 3000 rpm for 20 min. The extract was evaporated to the aqueous phase partitioned against dichloromethane. The dichloromethane fraction was dried and dissolved in 4 mL of 25% methanol then filtered. Samples of the solutions (20 μL) were injected into an HPLC system (Agilent1200; Agilent Technologies, Santa Clara, CA, USA) equipped with a C18 column (Inertsil ODS2 4.6 × 150 mm; GL Science, Tokyo, Japan) and an auto injection system. The column was eluted with a gradient of 20–70% methanol for 26 min at 40°C. The flow rate was 1.2 mL·min−1. For scoparone analysis, each treatment was repeated eight times (replications), using one fruit per replication.
Long-term storage methodThe satsuma mandarin ‘Aoshima unshu’ fruits were harvested from an orchard in the Shimizu area, Shizuoka city, December 1 to December 3, 2015. The harvested fruits were maintained at room temperature for two weeks, following which approximately 100 fruits were put in one flat plastic container (outer dimensions: L615 mm × W410 mm × H190 mm, inner dimensions: L565 mm × W375 mm × H180 mm). Two hundred containers were stored for 120 days in a storage room (30 m3) at 8°C. Temperature and RH in the containers under both the blue LED treated and non-treated storage conditions were measured using the thermorecorder RS-13 (Espec, Aichi, Japan).
Two tape-type (3.0 m) blue LEDs (Hamamatsu Puls, Shizuoka, Japan) with a maximum emission wavelength of 465 nm were inserted into the upper gap of the containers (Fig. 2), and the fruits were irradiated for 90 days (from January 22 to April 19; the fruits were in storage from December 22 to April 19). The photon flux of this tape-type LED is 10 μmol·m−2·s−1, and they were used for 12 h each day, a duration chosen considering the fact that blue LED with an emission peak of 465 nm and photon flux of 8 μmol·m−2·s−1 significantly reduces the growth of blue mold (P. italicum) (Yamaga et al., 2015b), while the phospholipase A2 gene expression in Citrus sinensis exposed to a 12/12 h light/dark cycle of blue light (photon flux 3 μmol·m−2·s−1) was higher than that in plants under dark treatment (Liao and Burns, 2010).

Experimental setup of blue light-emitting diodes (LEDs) (left) and blue LED irradiation on the fruits during storage (right).
The decayed fruits from each treatment were counted at 29, 64, 92, and 120 days after the start of storage, and the decayed fruits were removed on the date of each counting. The total decay rate was calculated according to the following formula: Total decay rate = (total number of decayed fruits/total number of examined fruits) × 100. Thirteen and thirty replicates were made for LED and dark treatments respectively, using approximately 100 fruits per replicate (container).
Internal fruit quality and rind color after long-term storageAfter storage, internal fruit quality and rind color were examined. Specific gravity was determined by weighing fruits in air and water. The percentage of flesh was the ratio of flesh weight to whole fruit weight. The soluble solid content (SSC) and titratable acidity (TA) were determined using freshly extracted juice. The SSC was determined using a digital refractometer (DBX-55A; Atago, Tokyo, Japan). The TA was determined by titrating 5 mL of the juice to pH 8.1, using 0.156 mol·L−1 NaOH. The acidity of the juice is expressed as grams of citric acid per 100 mL of juice. Rind color was measured using a color analyzer (TC-1500SX; Tokyo Denshoku, Tokyo, Japan). The CIELAB (L* = lightness, a* = bluish-green/red purple hue component, b* = yellow/blue hue component) of the rinds was measured. Five fruits constituted a single replicate (container) and each treatment was repeated ten times.
Statistical analysisStatistical analysis was performed using statistical functions in Microsoft Office Excel 2007 for Windows (Microsoft Corporation, Redmond, WA, USA). The difference between the means was evaluated by Student’s t-test.
The effect of blue LED irradiation on natural infection of wounded fruit is shown in Figure 3. The mean decay rate of LED irradiated fruit was 13.3%. In contrast, decay of non-irradiated fruit was 51.1%. Fruits irradiated with blue LED showed significantly reduced decay rates, compared to those in the dark treatment. Natural incidence of postharvest diseases under the experimental conditions included blue and green molds under both treatments.

Effect of blue LED irradiation on natural disease infection of wounded fruits. Data are expressed as the mean ± SE (n = 3). Data were transformed to the arcsine of the square root before analysis. ** indicates significant difference at P < 0.01 by t-test.
Scoparone accumulation in the flavedo tissue was determined in the wounded and intact fruits, both exposed and unexposed to blue LED irradiation. The results are shown in Figure 4. The blue LED treatment resulted in a significantly higher production of scoparone in wounded fruit compared with the dark treatment. The scoparone content in wounded and blue LED irradiated fruit was 2.3 μg·g−1 FW. In contrast, wounded, but non-irradiated fruits accumulated only a small amount of scoparone (0.5 μg·g−1 FW). Intact fruit accumulated even smaller amounts of scoparone (0–0.08 μg·g−1 FW), irrespective of blue LED irradiation.

Effect of blue LED irradiation on scoparone production in the flavedo of satsuma mandarin fruits. Data are expressed as the mean ± SE (n = 8). ** indicates significant difference at P < 0.01 by t-test.
Temperature and RH in containers with or without blue LED irradiation during storage are shown in Figure 5. The temperature of LED treatment remained slightly higher compared to the dark treatment. Average temperatures of LED and dark treatments from February 24 to April 15 were 8.2°C and 7.8°C, respectively; RH under the LED treatment remained low compared to the dark treatment. Average RH values in LED and dark treatments for the same period mentioned above were 89.0% and 92.7%, respectively. Changes in total fruit decay rate during storage are illustrated in Figure 6. After up to 64 days in storage (36 days after starting LED irradiation), the total decay rate was not significantly different between LED-treated and untreated fruits. On the other hand, from 91 to 120 days of storage, total fruit decay in the blue LED treatment was significantly lower compared to that of the control group. After 120 days of storage, the decay rate in the blue LED treatment was 13.7%, whereas that of the control treatment reached 18.4%.
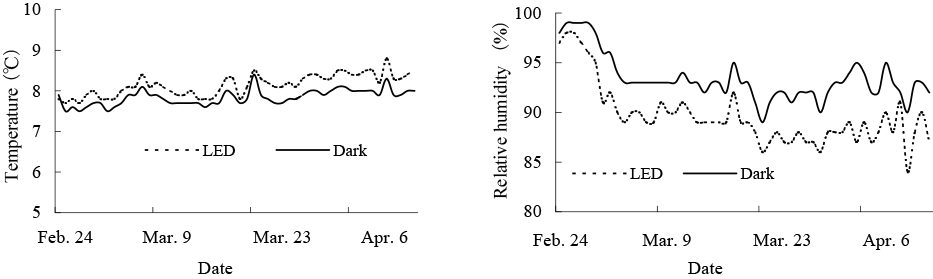
Temperature and relative humidity in storage containers during long-term storage.

Changes in total decay rate of satsuma mandarin fruits during long-term storage. Data are expressed as the means of 13 (LED) and 30 (Dark) replicates. Data were transformed to the arcsine of the square root before analysis. * and ** indicate significant difference at P < 0.05 and 0.01, respectively, by t-test.
The influence of blue LED irradiation on internal fruit quality and rind color of satsuma mandarin ‘Aoshima unshu’ fruits during long-term storage is shown in Table 1. Irradiation with tape-type blue LED (10 μmol·m−2·s−1) did not affect either fruit quality as analyzed in this study (SSC, TA, specific gravity, percentage of flesh) or rind color, as evaluated by coefficient (L*), a* value, and b* value.

Influence of blue LED irradiation on the fruit quality of satsuma mandarin fruits (Citrus unshiu Marcow. ‘Aoshima unshu’) after long-term storage.
Previous studies have revealed that fungal colonization and development of disease areas in fruits inoculated with P. digitatum, P. italicum, and Phomopsis citri were reduced by exposure to blue light (Alférez et al., 2012; Liao et al., 2013; Yamaga et al., 2015a). In contrast, we report that blue LED irradiation reduced the natural infection by Penicillium in wounded fruits (non-inoculated). The results of the experiment (Fig. 3) show that although the wounded parts of the fruits are prone to being infected with Penicillium, blue light exposure significantly reduces the chance of pathogenesis.
We examined the effect of blue LED at a photon flux of 80 μmol·m−2·s−1 on scoparone production and showed that only wounded fruits showed significantly increased scoparone levels upon exposure to blue LED irradiation (Fig. 4). On the other hand, Ballester and Lafuente (2017) reported that scoparone content increased mainly in response to blue LED irradiation at a photon flux of 630 μmol·m−2·s−1 applied for 18 h, however, they did not rule out the possibility that the scoparone increase may have reflected an oxidative stress condition in citrus fruit peels exposed to excess light. The photon flux used in our experiments was lower than that used in their study; therefore, oxidative stress in citrus fruit peels was lower in our experiment. Instead, our results suggest a synergistic effect between blue LED and the stress caused by wounding because blue LED-irradiated intact fruits accumulated little scoparone. Phenylalanine ammonia-lyase (PAL) activity is widely known to be activated by wounding stress (Dan et al., 1999). It is increased by blue light irradiation in some plants (Engelsma, 1974) and by blue light induced accumulation of reactive oxygen species in vivo (Jourdan et al., 2015). Considering that PAL activity is increased by stress conditions other than wounding, the scoparone production observed in this study could also have been a reaction to multiple stresses as mentioned above. Although we demonstrated that the scoparone content increased significantly in artificially wounded fruit when irradiated with blue LED at a photon flux of 80 μmol·m−2·s−1 (Fig. 4), the production levels were still very low compared to those induced by UV-C or UV-B treatment (Kuniga et al., 2015; Yamaga et al., 2016). These results led us to conclude, in agreement with Ballester and Lafuente (2017), that low scoparone production induced by blue LED irradiation was not the main factor involved in the induced resistance observed. Therefore, we need to study enzyme activities and phytoalexins other than coumarins in satsuma mandarin fruits. The increase in phospholipase A2 activity in flavedo tissue in response to blue light exposure was associated with a reduction in postharvest citrus decay (Alférez et al., 2012). Additionally, Liao et al. (2013) also showed that the aldehyde octanal, which plays an important role in the antifungal response, was markedly induced by blue light exposure in tangerines. PAL and scoparone have been related to UV irradiation-induced resistance in this fruit crop (D’hallewin et al., 1999). PAL has been linked to the elicitation of resistance in citrus fruit peel against P. digitatum (Ballester et al., 2013). The relation between PAL activity and blue LED irradiation to postharvest mandarin fruit resistance to decay caused by fungal infection also needs to be further characterized.
Blue LED irradiation could directly inhibit fungal growth in satsuma mandarin fruits (Yamaga et al., 2015b). Blue light exposure was also able to stop colony growth of P. digitatum, which had already grown a mycelium, and no further growth occurred if three-day LED treated colonies were transferred to darkness (Lafuente and Alférez, 2015). This indicates that the effect of blue light exposure lasts for a certain period after removal of the light stimulus. Further, we showed that intermittent blue LED irradiation effectively controlled the incidence of the disease caused by P. italicum (Yamaga et al., 2015a). Considering our findings in this study, we suggest that intermittent blue LED irradiation at a rate of 12 h per day would be enough to suppress mycelial growth, even in a storage room. In the early stages of storage (storage time: 0–29 days), the fruit decay rate was low, increasing towards the later period of storage (storage time: 92–120 days) (Fig. 6). Blue LED irradiation treatment was started 29 days after storage initiation in this experiment; thus, the protocol used here offers the additional benefit of an energy saving effect.
Temperature in the containers irradiated with blue LED tended to remain slightly higher compared to that in containers in the dark treatment (Fig. 5). This difference could play a role in reducing fruit decay, including chilling injury and rind disorder. Blue light is known to induce the activation of the plasma membrane and stomatal opening (Shimazaki, 1994), which suggest that disease resistance in the flavedo tissue was increased by transpiration. Investigating the effect of blue LED on transpiration in mandarin fruits may be used to identify the factors that suppress fruit decay. Humidity control during low temperature storage of satsuma mandarin fruits is more difficult during and after March because the outside ambient air humidity becomes higher then. Moisture enhances rind puffing and fruit decay due to weakening of the peel. Light irradiation by the tape-type LED may cause convection and air circulation in a small space inside the container. Tape-type LED is believed to reduce the retention of moisture on the fruit surface, assisting its release to the ambient air. LED has higher energy efficiency than conventional fluorescent lamps and generates less heat, but in closed environments, the temperature becomes slightly higher as mentioned above. Therefore, blue LED may be recommended for use in storage rooms with cooling facilities. Although the beneficial effect of tape-type LED on stored fruit may be limited because of incomplete fruit coverage by the irradiation, blue LED (e.g., irradiation on wall surfaces in a storage room) would be a useful tool for reducing contamination, even if the light does not reach the entire surface of every fruit in the storage room. In a previous study, we showed that blue LED (fluorescent tube type) irradiation on the wall surfaces in a storage room significantly reduced mandarin fruit decay caused by blue and green molds (data not shown).
Tape-type blue LED irradiation at 10 μmol·m−2·s−1 did not affect fruit quality characteristics (Table 1). In our previous study, we reported that fruits treated with blue LED irradiation at 80 μmol·m−2·s−1 for 6 days in a chamber (15°C) showed different citric acid concentrations from those of the control treatment (Yamaga et al., 2015a). It is possible that blue LED irradiation at 10 μmol·m−2·s−1 at low temperature (8°C) has little effect on characteristics related to fruit quality. Future research will undertake examination of the influence of blue LED irradiation on moisture loss by fruits in order to gain further understanding of the protective mechanism at play.
In conclusion, we showed that blue LED irradiation reduced fruit decay in a storage room without affecting internal fruit quality or rind color. These results seem to be due to the combined effect of fungal growth inhibition by LED irradiation and phytoalexin production (scoparone) synergistically induced by wounding stress. Reduction of RH due to LED irradiation is also considered a factor involved in the reduction of fruit decay under the experimental conditions used here. Additionally, we propose that a sequential preprogrammed use of tape-type blue LED devices, followed by intermittent LED irradiation and finally constant irradiation beginning 29 days after storage initiation, would lead to a cost reduction compared to irradiation with normal LED lamps or fluorescent lamps during the total storage period. Efficient blue light irradiation in the storage room will require the use of reflecting materials on the container walls or spacing racks within each storage room to maximize fruit coverage by the irradiation treatment. Successful introduction of blue LED irradiation systems in actual storage rooms at production sites requires the maximum reduction of the initial cost of the LED.