Abstract
Hydrangea (Hydrangea spp.) flower buds usually differentiate from the end of summer through autumn, but some cultivars can also produce flower buds in spring. In the present study, we selected cultivars with high potential for such unseasonable flower bud production in Japan by evaluating the flower bud production on pinch-treatment-induced axillary shoots. We also examined the pinching time and the difference between the two shoot types: previously formed shoots that developed from buds formed in the previous autumn and basal shoots that formed in the current spring. We found that ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ had the highest frequencies of unseasonable flower bud production on the axillary shoots of previously formed shoots (90.0%, 80.0%, and 90.0%, respectively) and basal shoots (90.0% for all three cultivars). When the buds were pinched in mid-April, the axillary shoots that formed on both shoot types flowered from early July to mid-August, which was approximately 1 month later than for seasonably produced flower buds in the same cultivars. In 2011, 98.0%, 90.5%, and 84.0% of the axillary shoots that developed on previously formed shoots flowered in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively, while in 2012, 63.2%, 54.9%, and 75.3% flowered, respectively. Similar values were seen for the basal shoots, with the exception of ‘Christmas’ in 2011 and ‘Endless Summer’ in 2012, which both had lower flowering rates than previously formed shoots. In ‘Rosea’, flower bud differentiation started just after the pinch treatment, and the frequency of flower bud production decreased when the pinch treatment was conducted after late April. Thus, ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ were considered to have high potential for unseasonable flowering following pinch treatment in early spring. However, unseasonable flower bud production may often be suppressed in pinched basal shoots.
Introduction
Hydrangeas, which include Hydrangea macrophylla, H. serrata and their hybrids, are common ornamental plants cultivated as pot flowers, shrubs, and cut flowers in many countries (Mallet, 2008; van Gelderen and van Gelderen, 2004). Hydrangea plants develop two types of shoots in spring: current shoots that develop from terminal or axial buds that formed in the previous autumn (previously formed shoots) and vigorous shoots that develop from the basal part of the plant (basal shoots) (Fig. 1).

Floral differentiation in hydrangeas usually occurs from the end of summer through autumn on previously formed shoots or basal shoots, and the resulting flower buds then develop as previously formed shoots in the following spring once winter dormancy has been broken, with anthesis occurring until summer (Zhou and Hara, 1988). When flower buds that differentiated in the previous autumn are removed after spring, the shoots rarely produce more flower buds. Many studies have investigated the regulation of this type of flower bud differentiation and anthesis (Adkins et al., 2002; Eid et al., 2016; Nordli et al., 2011; Orozco-Obando et al., 2005).
Some hydrangea cultivars can produce flower buds and flower unseasonably–for example, Kudo et al. (2008) produced an interspecific hybrid between H. scandens ssp. chinensis and H. macrophylla, that flowered during winter and did not show dormancy. Adkins and Dirr (2003) examined the frequency of flowering in the vegetative cuttings of several hydrangea cultivars that had been obtained after spring and called those cultivars that flowered at a high frequency (remontant) cultivars, which means flowering twice in a single growing season. Such unseasonable flower bud production has several advantages in the horticultural industry such as expansion of the harvest period and an increase in the harvest.
In hydrangeas, unseasonably produced apical flower buds of basal shoots are used for cut flower production and are essential for increasing the cut flower harvest (Kitamura et al., 2014). Furthermore, although the unseasonable flower bud production trait is rarely used in pot hydrangea flower production in Japan, the stable induction of this trait could lead to the establishment of a novel production system that does not require heating of the cultivation area to protect the flower buds from freezing winter temperatures. However, it is often difficult to exhibit the unseasonable flower bud production trait stably (Kitamura, 2016). Unfortunately, Adkins and Dirr (2003) did not provide detailed information about the factors that affect flower bud production after spring and included several cultivars that are not common in Japan in their study. Therefore, further information on the factors that affect this trait and the cultivars that are common in Japan is required before it can be applied to pot or cut hydrangea flower production.
In this study, we selected cultivars with high potential for unseasonable flower bud production from 12 hydrangea cultivars that were readily available in Japan. We then examined the optimum shoot type and pinching time for the application of unseasonable flower bud production to hydrangea production in Japan.
Materials and Methods
Plant materials
In this study, 12 hydrangea cultivars were examined: ‘Benton’, ‘Christmas’, ‘Endless Summer’, ‘Flambeau’, ‘Green Glove’, ‘Grünherz’, ‘Libelle’, ‘Madame Plumecoq’, ‘Masja’, ‘Rosea’, ‘Sekka Yae’, and ‘Temariezo’. Adkins and Dirr (2003) previously reported that ‘Endless Summer’ produced flower buds after spring but did not examine any of the other cultivars which were selected as candidate cultivars in this study because they were readily available and were not patented (i.e., could be used for production without any permission). ‘Benton’, ‘Christmas’, ‘Endless Summer’, ‘Flambeau’, ‘Green Glove’, ‘Grünherz’, ‘Libelle’, ‘Madame Plumecoq’, ‘Masja’, ‘Rosea’, and ‘Sekka Yae’ are hybrids between or among H. macrophylla and H. serrata. ‘Temariezo’ is H. serrata and was introduced from a wild population in Fukushima Prefecture, Japan (Uemachi et al., 2014). Origin, inflorescence type, and flower color of used cultivars are shown in Table 1.

Plant cuttings were prepared in 2010 and used in all of the experiments. In all cultivars except ‘Endless Summer’ and ‘Temariezo’, vegetative shoots were obtained as cuttings from Kyoto University (Kyoto, Japan) in July 2010. In addition, 3 mother plants of ‘Endless Summer’ were purchased from a nursery in June 2010, while one mother plant of ‘Temariezo’ was obtained from the University of Shiga Prefecture (Hikone, Japan) in 2007. All of the cuttings with one node were rooted at the Shinshu University experimental farm (Minamiminowa, Japan) under periodic misting in June 2010. The plants were then transplanted into 19.5 cm-diameter pots filled with 3 L of medium consisting of 75% Metro Mix 250 (SunGro Horticulture Distribution, USA) and 25% vermiculite (v/v) (Asahi Kogyo, Japan) in early September 2010. Plants were fertilized every 2 weeks using 1 L of 1:1000 diluted Hanakoujyo (N:P:K = 50:100:50 mg·mL−1; Sumitomo Chemical Garden Products Inc., Japan). Plants were grown from December to March in a greenhouse heated above 0°C and were grown in a greenhouse without heating from April to November. The plants were grown under full sunlight from October to May and under 50% sunlight using Dio Cool White (Dio Kasei, Japan) 620SW (50% shading) from June to September.
Pinch treatment and evaluation of flower bud production
The potential for unseasonable flower bud production was examined for the pinch-treatment-induced axillary shoots of previously formed shoots in all 12 cultivars and basal shoots in all cultivars except ‘Benton’, ‘Flambeau’, ‘Green Glove’, and ‘Grünherz’ (Fig. 1). From April 6 to 14, 2011, the shoots were pinched to 2–3 nodes bearing vegetative axillary buds, and all other shoots were removed. This treatment removed any floral buds that had been produced up to the previous autumn. From May 24 to 26, 2011, 10 apical buds of the developed axillary shoots were dissected under a stereomicroscope (SZ60; Olympus Co., Ltd., Japan), and the developmental stage of each apical bud was determined according to Kosugi and Arai (1960; Fig. 2). The number of nodes with fully expanded leaves was recorded. Ten plants per cultivar were used. For each shoot type, 7–10 shoots which had 3–4 fully expanded leaves with vegetative axillary buds were used.
Flowering time of the unseasonably produced flower buds
‘Christmas’, ‘Endless Summer’, and ‘Rosea’ produced flower buds at a high frequency, and the developmental stages of their flower buds were higher than those of any other cultivars. Thus, these cultivars were studied for the flowering time of the unseasonably produced flower buds that developed on previously formed shoots and basal shoots. On April 18, 2012, pinch treatment was conducted as above. The number of axillary shoots that flowered every week until the end of August 2012 was counted. When approximately 80% of the decorative florets in the inflorescence had developed the cultivar-specific coloration of the decorative sepals, the inflorescence was regarded as flowered (Fig. 3A). In total, 12, 14, and 18 plants of ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively, were used. For previously formed shoots, 68, 51, and 126 axillary shoots developed from 27, 25, and 50 pinched shoots were examined, while for basal shoots, 73, 151, and 114 axillary shoots developed from 28, 60, and 45 pinched shoots were studied for each cultivar, respectively.

The flowering rates of unseasonably produced flower buds on previously formed shoots and basal shoots of ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ were studied in 2011 and 2012. From April 6 to 14, 2011 and on April 18, 2012, pinch treatment was conducted as outlined above. Numbers of flowered axillary shoots, shoots with aborted flower buds, and vegetative shoots were recorded in October 2011 and 2012 (Fig. 3B–D). In 2011, 27, 30, and 30 plants were used for ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively. For previously formed shoots, 53, 95, and 94 axillary shoots developed from 28, 38, and 38 pinched shoots were studied for each cultivar, respectively, while for basal shoots, 85, 156, and 205 axillary shoots developed from 28, 45, and 59 pinched shoots were studied for each cultivar, respectively. In 2012, 12, 14, and 18 plants were used for ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively. For previously formed shoots, 68, 51, and 126 axillary shoots developed from 27, 25, and 50 pinched shoots were studied, respectively, while for basal shoots, 73, 151, and 114 axillary shoots developed from 28, 60, and 45 pinched shoots were studied for each cultivar, respectively.
Flower bud initiation in the axillary shoots
Twenty ‘Rosea’ plants were used to obtain details of flower bud initiation in the axillary shoots of pinched previously formed shoots. On April 18, 2012, pinch treatment was conducted on 100 shoots as stated above. Then 6–10 apical meristems of the axillary shoots were randomly collected twice per week from the day of the pinch treatment until May 14, 2012. Collected samples were fixed in 40% formaldehyde/acetic acid/ethanol/water = 1:1:9:9 (volume basis). Then, samples were dehydrated in a graded ethanol series (30%, 50%, 70%, 80%, 90%, 95%, 99.5%, and 100%) and the samples were substituted with xylene. Next, the segments were embedded in paraffin at 60°C (Paraplast Plus; Sigma-Aldrich Co., Ltd., USA). The embedded samples were sectioned on a microtome at a thickness of 10 μm. Sections were extended and deparaffinized according to standard procedures and stained with 1:1000 diluted safranin O (Muto Pure Chemicals Co., Ltd., Japan). The cross sections were then observed under an optical microscope (BH2; Olympus Co., Ltd.), and the developmental stage of each apical meristem was determined according to Kosugi and Arai (1960; Fig. 2).
Effect of pinching time on unseasonable flower bud production
The effect of pinching time on unseasonable flower bud production on the axillary shoots developed from previously formed shoots was studied using 7 plants from each of ‘Flambeau’ and ‘Rosea’. ‘Flambeau’ was used as a representative of cultivars with low potential for unseasonable flower bud production. ‘Rosea’ exhibited high potential for unseasonable flower bud production stably during 2011 and 2012, so this cultivar was used as a representative of cultivars with high potential for unseasonable flower bud production. From March 31 to May 26, 2014, pinch treatment was conducted every two weeks on 10 previously formed shoots from each cultivar as stated above. On October 18, 2014, the numbers of flowered axillary shoots, shoots with aborted flower buds, and vegetative shoots were recorded for 8–10 axillary shoots that had developed in each pinch treatment (Fig. 3B–D).
Statistical analysis
Data collected for the cultivar difference in flower bud production were analyzed using the Steel-Dwass test. Data collected for the number of nodes that were produced on axillary shoots were analyzed using Tukey’s multiple range test.
Results
Frequency of unseasonable flower bud production on axillary shoots differed among the cultivars
In both shoot types, the median developmental stage of unseasonably produced flower buds on axillary shoots was significantly higher in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ than in all other cultivars except ‘Libelle’ (Table 2). Furthermore, ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ also had a higher rate of axillary shoots with flower buds for both previously formed shoots (90%, 80%, and 90%, respectively) and basal shoots (90% for all three cultivars). In ‘Libelle’, 30% and 40% of the axillary shoots from previously formed shoots and basal shoots, respectively, produced flower buds. Similarly, 30% and 40% of the axillary shoots of previously formed shoots developed flower buds in ‘Green Glove’ and ‘Masja’, respectively, but flower bud development was slower than in ‘Libelle’. The percentage of axillary shoots with flower buds was lower on the basal shoots than on the previously formed shoots in ‘Madame Plumecoq’ and ‘Masja’. Flower bud production was rarely or never observed on the axillary shoots of either shoot type in any of the remaining cultivars. Although the number of nodes on the axillary shoots of both shoot types significantly differed among the cultivars examined, no common trend was observed in the development of unseasonably produced flower buds.
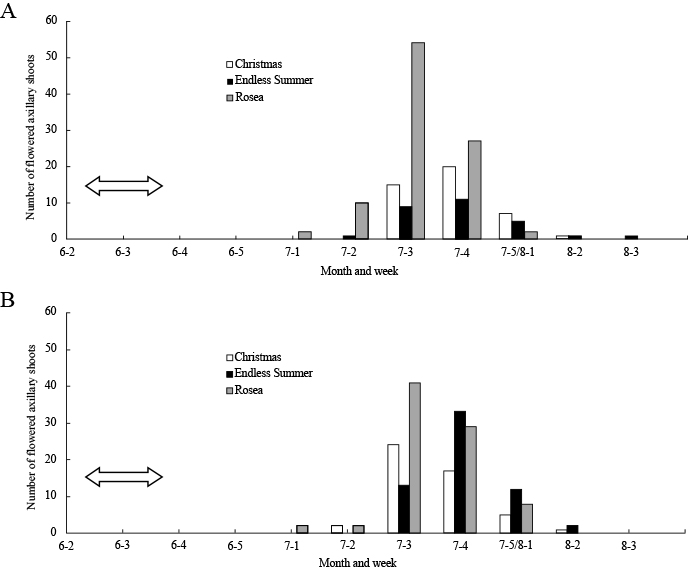
Unseasonably produced flower buds on the axillary shoots of both shoot types in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ flowered from early July to mid-August, which was approximately 1 month later than the flowering periods of seasonably produced flower buds in the same cultivars (Figs. 4 and 5). In all three cultivars, the flowering period of the unseasonably produced flower buds was 3–6 weeks and did not differ between the shoot types.
Flowering rate of unseasonably produced flower buds differed between years
In 2011, 98.0%, 90.5%, and 84.0% of the axillary shoots of previously formed shoots, and 80.0%, 82.7%, and 89.8% of those of basal shoots flowered in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively (Table 3). No axillary shoots with aborted flower buds were observed for either shoot type in 2011, with the remaining axillary shoots exhibiting vegetative growth during the study period. By contrast, in 2012, 63.2%, 54.9%, and 75.4% of the axillary shoots of previously formed shoots, and 67.1%, 38.4%, and 71.9% of those of basal shoots flowered in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, respectively. Furthermore, aborted flower buds were observed on 20.6%, 13.7%, and 11.9% of the axillary shoots of previously formed shoots, and 8.2%, 10.6%, and 12.3% of those of basal shoots in each cultivar, respectively. The remaining axillary shoots exhibited vegetative growth during the study period.
Flower bud development was observed in the apical meristems of the axillary shoots 8 days after the pinch treatment. Flower buds continuously developed on the axillary shoots during the study period and developed partial inflorescence primordia (Fig. 6).
Frequency of flowering of axillary shoots decreased as the pinch treatment was delayed
In ‘Rosea’, 80% of the axillary shoots that developed after the pinch treatments on March 31 and April 15 flowered until mid-October (Table 4). However, the percentage of flowered axillary shoots decreased as the pinch treatment was delayed, with no flowering being observed in the axillary shoots that developed after the pinch treatment on May 26. By contrast, the axillary shoots of ‘Flambeau’ did not flower following any of the pinch treatments, with 10% of the axillary shoots that were formed by the pinch treatment on March 31 and April 15 exhibiting aborted flower buds, and the remaining shoots in all treatments exhibiting vegetative growth.
Discussion
Adkins and Dirr (2003) reported that ‘Endless Summer’ had high potential as a remontant cultivar and the result of our study supported this. In addition, ‘Christmas’ and ‘Rosea’, which were not studied by Adkins and Dirr (2003), were newly identified as additional cultivars with high potential for unseasonable flower bud production in the present study. Katayama et al. (2017) conducted phylogenetic analysis of some hydrangea cultivars and lines using nuclear SSR marker and the nucleotide sequence of internal transcribed spacer (ITS) region and reported that ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ belonged to the same clade. Thus, the high potential for unseasonable flower bud production observed in those three cultivars may originate from the same ancestor. A relatively high percentage of the axillary shoots of previously formed shoots had flower buds in ‘Green Glove’, ‘Libelle’, and ‘Masja’, but these developed to a much lower stage than those of ‘Christmas’, ‘Endless Summer’, and ‘Rosea’, reducing their potential for unseasonable flower bud production in the present study.
The number of nodes is considered to have a major effect on flower bud production on axillary shoots (Dennin and McDaniel, 1985). Adkins and Dirr (2003) reported a negative correlation between the number of nodes and the developmental stage of unseasonably produced flower buds in several hydrangea cultivars, whereas we observed no relationship between them. This difference may have resulted from the different data collection methods used: Adkins and Dirr (2003) used combined data of number of nodes obtained from three different harvest dates, whereas we evaluated the number of nodes approximately 45 days after the pinch treatment in both shoot types.
In both shoot types, the unseasonably produced flower buds of ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ flowered from early July to mid-August (Fig. 5). We previously reported that some cultivars that had been bred for cut flower production produced unseasonable flowers in the apical buds of unpinched basal shoots from July to November, and that the difference in the time of floral induction may be related to the variation in the flowering time (Kitamura et al., 2014). It appears that the flowering period of unseasonably produced flower buds observed in the present study was shorter than that of apical buds of the unpinched basal shoots observed in our previous report. In ‘Rosea’, unseasonable flower bud production on the axillary shoots of previously formed shoots started just after the pinch treatment (Fig. 6). Also, in ‘Christmas’ and ‘Endless Summer’, unseasonable flower bud production would start simultaneously in the axillary shoots just after the pinch treatment, and the flowering periods may have become shorter than those on the apical buds of unpinched basal shoots.
The percentage of axillary shoots that flowered in ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ was higher in 2011 than in 2012 for both shoot types. The pinch treatment was conducted 4–12 days earlier in 2011 than in 2012 (April 6 to 14 vs. April 18, respectively) and our investigation of ‘Rosea’ showed that a delayed pinch treatment decreased the percentage of flowered axillary shoots (Table 3). The difference in the time of pinch treatment could be a factor for the difference in the percentage of flowered axillary shoots between those two years.
Unseasonable flower bud production in the axillary shoots started just after the pinch treatment (Fig. 6), suggesting that some factor in pinch treatment accelerated the unseasonable flower bud production. Decreases in temperature and day length in autumn trigger seasonable floral induction in hydrangeas (Struckmeyer, 1950). In particular, temperatures of around 18°C cause floral induction regardless of the photoperiod, while a photoperiod of up to 12 h causes floral induction when the temperature is above 22°C (Bailey and Weiler, 1984; Litlere and Strømme, 1975). Adkins and Dirr (2003) reported that the remontant cultivars they selected could produce flower buds even under a 24 h photoperiod at 24°C. In mid-April and mid-May 2014, the mean temperatures in our experimental condition were approximately 12.0°C and 16.3°C, and the day lengths were approximately 13 h and 14 h, respectively (data not shown), both of which were inductive for seasonable floral bud production of hydrangeas due to the relatively low temperatures. Moreover, the temperatures were lower and the day lengths were shorter than in Adkins and Dirr’s (2003) study, in which they selected ‘Endless Summer’ as a remontant cultivar. Despite these appropriate conditions for flower bud production, the axillary shoots that were developed by the pinch treatment on May 13, 2014 rarely flowered in ‘Rosea’ (Table 3), suggesting that this cultivar may have a lower potential for unseasonable flower bud production than ‘Endless Summer’. However, the percentage of flowered axillary shoots was higher in ‘Rosea’ than in ‘Endless Summer’, with the exception of the axillary shoots of previously formed shoots in 2011 (Table 3), indicating that there is little difference in the potential for unseasonable flower bud production between these two cultivars. There could be a factor other than temperature or daylength that regulates the occurrence of unseasonable flower bud production. This factor may be optimal in early spring in Japan and caused a high frequency of unseasonable flower bud production in the present study.
There was no difference in unseasonable flower bud production between previously formed shoots and basal shoots with two exceptions: the percentage of axillary shoots with flower buds was lower in the basal shoots of ‘Madame Plumecoq’ and ‘Masja’ while the percentage of flowered axillary shoots was lower in the basal shoots of ‘Christmas’ in 2011 and ‘Endless Summer’ in 2012 (Table 3). Thus, it appears that unseasonable flower bud production may often be suppressed in the basal shoots.
In conclusion, ‘Christmas’, ‘Endless Summer’, and ‘Rosea’ were selected as cultivars with high potential for unseasonable flower bud production. However, application of pinch treatment in early spring would be necessary for unseasonable flower bud initiation, and unseasonable flower bud production may often be suppressed in pinched basal shoots. Further research is required to clarify the relationship between the unseasonable flowering potential evaluated in this study and that of the apical buds of unpinched basal shoots, with a particular focus on the flowering period.
Acknowledgement
We are very grateful to Dr. Munetaka Hosokawa (Kyoto University) for providing us several constructive opinions on the study.
Literature Cited
- Adkins, J. A. and M. A. Dirr. 2003. Remontant flowering potential of ten Hydrangea macrophylla (Thunb.) Ser. cultivars. Hort. Sci. 38: 1337–1340.
- Adkins, J. A., M. A. Dirr and O. M. Lindstrom. 2002. Cold hardiness potential of ten hydrangea taxa. J. Environ. Hort. 20: 171–174.
- Bailey, D. A. and T. Weiler. 1984. Control of floral initiation in florist’s hydrangea. J. Amer. Soc. Hort. Sci. 109: 785–791.
- Dennin, K. A. and C. N. McDaniel. 1985. Floral determination in axillary buds of Nicotiana silvestris. Dev. Biol. 112: 377–382.
- Eid, G. M., N. Albatal and S. Haddad. 2016. Effect of photoperiod on the flowering of some cultivars of hydrangea (Hydrangea macrophylla). Int. J. Hort. 6: 1–7.
- Katayama, C., T. Fudano, Y. Kitamura, T. Uemachi, E. Kesmawati and M. Hosokawa. 2017. Phylogenetic analysis of Hydrangea spp. in Indonesia by nuclear SSR marker and ITS region. Hort. Res. (Japan) 16 (Suppl. 2): 276 (In Japanese).
- Kitamura, Y. 2016. Production of cut hydrangea flower. p. 93–102. In: Nosangyosonbunkakyoukai (ed.). Resent Techniques in Floriculture. Series 8 (In Japanese). Nousangyosonbunkakyoukai, Tokyo.
- Kitamura, Y., K. Shimoyama and T. Hattori. 2014. Floral bud differentiation and flowering time of basal-shoot in cut flower hydrangea. Hort. Res. (Japan) 13 (Suppl. 1): 188 (In Japanese).
- Kosugi, K. and N. Arai. 1960. Studies on the flower bud differentiation and development in some ornamental trees and shrubs VII. On the date of flower bud differentiation and flower bud development in Hydrangea macrophylla. Tech. Bull. Fac. Agr. Kagawa Univ. 12: 78–83 (In Japanese with English abstract).
- Kudo, N., T. Matsui and T. Okada. 2008. A novel interspecific hybrid plant between Hydrangea scandens ssp. chinensis and H. macrophylla via ovule culture. Plant Biotechnol. 25: 529–533.
- Litlere, B. and E. Strømme. 1975. The influence of temperature, daylength, and light intensity on flowering in Hydrangea macrophylla (Thunb.) Ser. Acta Hortic. 51: 285–298.
- Mallet, C. 2008. Hydrangea. Eugen Ulmer, Paris.
- Nordli, E. F., M. Strøm and S. Torre. 2011. Temperature and photoperiod control of morphology and flowering time in two greenhouse grown Hydrangea macrophylla cultivars. Sci. Hortic. 127: 372–377.
- Orozco-Obando, W., G. N. Hirsch and H. Y. Wetzstein. 2005. Genotypic variation in flower induction and development in Hydrangea macrophylla. Hort. Sci. 40: 1695–1698.
- Struckmeyer, E. B. 1950. Blossom bud induction and differentiation in hydrangea. Proc. Amer. Soc. Hort. Sci. 56: 410–414.
- Uemachi, T., Y. Mizuhara, K. Deguchi, Y. Shinjo, E. Kajino and H. Ohba. 2014. Phylogenetic relationship of Hydrangea macrophylla (Thunb.) Ser. and H. serrata (Thunb.) Ser. evaluated using RAPD markers and plastid DNA sequences. J. Japan. Soc. Hort. Sci. 83: 163–171.
- van Gelderen, C. J. and D. M. van Gelderen. 2004. Encyclopedia of Hydrangeas. Timber Press Portland, Oregon.
- Zhou, T. S. and N. Hara. 1988. Development of shoot in Hydrangea macrophylla. I. Terminal and axillary buds. Bot. Mag. Tokyo 101: 281–291.