原著論文
Effects of the pat-2 Gene and Auxin Biosynthesis Inhibitor on Seed Production in Parthenocarpic Tomatoes (Solanum lycopersicum L.)
2019 年 88 巻 4 号 p. 481-487

詳細
2019 年 88 巻 4 号 p. 481-487
In pat-2 parthenocarpic tomatoes (Solanum lycopersicum L.), inhibition of seed production prevents their breeding and development. Low seed set is thought to be caused by a pleiotropic effect of the pat-2 gene because the number of seeds from pat-2 parthenocarpic tomato cultivars is generally low; however, research on the relationship between low seed set and the pat-2 gene is lacking. In this study, to clarify the effect of the pat-2 gene on seed production and develop a method for promoting seed production in pat-2 parthenocarpic tomatoes, we examined the relationship between the pat-2 gene and low seed set and the effect of aminoethoxyvinylglycine (AVG), an indole-3-acetic acid (IAA) biosynthesis inhibitor, on seed production in ‘Renaissance’, a pat-2 parthenocarpic tomato cultivar. In an F2 population derived from a cross between ‘Micro-Tom’ and ‘Renaissance’, the number of seeds in plants homozygous for the pat-2 allele was significantly lower than that in heterozygous plants and that homozygous for the Pat-2 allele. This indicated that low seed set was genetically associated with the pat-2 allele, and that low seed set was unavoidable when using the pat-2 gene for breeding of parthenocarpic tomato cultivars. Therefore, it is important to develop a method to promote seed production in pat-2 parthenocarpic tomatoes. In this study, we examined the effect of AVG on seed production in ‘Renaissance’. We found that treatment with 30 μM AVG significantly increased the number of seeds in ‘Renaissance’. In addition, when we treated flower clusters with AVG solution at the first flower anthesis, the effect of treatment with 30 μM AVG on increasing seed set was highest for the flower in the second position in a cluster, which suggests the effect of AVG on promoting seed production changed depending on the stage and/or position. Our results showed that low seed set was genetically associated with the pat-2 allele and that AVG is an effective tool to promote seed production in pat-2 parthenocarpic tomatoes. Based on these results, research progress on improving seed production in parthenocarpic tomatoes with the pat-2 gene is expected to accelerate in the future.
Parthenocarpy induces fruit development either naturally or artificially without fertilization (Gustafson, 1942). In tomato cultivation, parthenocarpy is considered an attractive trait because it reduces the financial and labor costs of fruit setting and can prevent reductions in yield caused by unfavorable conditions, such as low or high temperatures and low or high humidity (George et al., 1984; Picken, 1984; Takisawa et al., 2017). In the tomato, there are nine parthenocarpic genes (name of line or cultivar): pat (‘Soressi’ and Montfavet191), pat-2 (‘Severianin’), pat-3/pat-4 (RP75/59), pat4.1/pat5.1 (IL5-1), pat4.2/pat9.1 (IVT-line1), and pat-k (‘MPK-1’) (Gorguet et al., 2005, 2008; Takisawa et al., 2017). Among these parthenocarpic lines, ‘Severianin’, possessing the pat-2 gene, produces fruits at high and low temperatures (Lin et al., 1982; Philouze and Maisonneuve, 1978a; Vardy et al., 1989) and its fruits are not puffy (Philouze and Maisonneuve, 1978b). Because of these advantages, the pat-2 gene is used as a breeding source for parthenocarpic tomato cultivars in Japan.
A parthenocarpic tomato cultivar, ‘Renaissance’, possessing the pat-2 gene was released in 2000 (Sugahara et al., 2002). It shows stable parthenocarpy, enabling high fruit setting and normal fruit development in spring, early summer, autumn, and winter, which are the blooming seasons of the main cropping system in Aichi Prefecture in Japan (Ohkawa et al., 2006). In addition, the percentage of fruit setting in ‘Renaissance’ is very high at high and low temperatures compared with that of non-parthenocarpic tomato cultivars (Ohkawa et al., 2007). However, seed production of pat-2 parthenocarpic tomatoes, including ‘Renaissance’, is low and unstable compared with that of non-parthenocarpic tomato cultivars (Ohkawa et al., 2008), and this inhibits the breeding of new pat-2 parthenocarpic tomato cultivars and their introduction. The low seed set is thought to be caused by a pleiotropic effect of the pat-2 gene because the number of seeds in pat-2 parthenocarpic tomato cultivars is generally low; however, research on the relationship between low seed set and the pat-2 gene is lacking. Identifying the genes that inhibit seed formation is important in order to understand the mechanism, and will support development of a method to promote seed production in pat-2 parthenocarpic tomatoes.
Although genes related to low seed set in pat-2 parthenocarpic tomato cultivars remain elusive, previous reports suggested two reasons for the low seed production: formation of a pseudoembryo and inhibition of pollen tube elongation. A pseudoembryo is formed from the innermost layer of the integument in the embryo sac cavity by treatment with synthetic auxin or gibberellin (GA) (Asahira et al., 1967; Kataoka et al., 2003; Serrani et al., 2007). Pseudoembryos are also observed in the embryo sacs of ‘Severianin’, and are thought to inhibit fertilization and seed production (Kataoka et al., 2008). Uniconazole, an inhibitor of GA biosynthesis, suppressed pseudoembryo development in ‘Severianin’ (Kataoka et al., 2003), and treatment with paclobutrazol, another inhibitor of GA biosynthesis, promoted seed production in ‘Renaissance’ (Ohkawa et al., 2012). In contrast, Johkan et al. (2010) suggested that a high level of IAA inhibited the elongation of pollen tubes at the style base in ‘Renaissance’, resulting in poor fertilization. They also reported that p-chlorophenoxyisobutyric acid (PCIB), an auxin action inhibitor, promoted seed production. These reports showed that plant growth regulators that inhibit GA biosynthesis or IAA action could promote seed production in pat-2 parthenocarpic tomato cultivars.
The indole-3-pyruvic acid (IPyA) pathway is thought to be primarily involved in IAA biosynthesis (Korasick et al., 2013; Zhao, 2012). In the IPyA pathway, Trp is converted to IPyA by tryptophan aminotransferase (TAA/TAR1), and IPyA is subsequently converted to IAA by the YUCCA family of flavin monooxygenases (Korasick et al., 2013). Although no inhibitors have been established for auxin biosynthesis, Soeno et al. (2010) found that aminoethoxyvinylglycine (AVG) and L-amino-oxyphenylpropionic acid (AOPP) directly blocked auxin biosynthesis through inhibition of TAA/TAR1, reducing the endogenous IAA level in tomato seedlings. In tomato ovaries, IAA levels increase after pollination, coincident with the upregulation of ToFZY2 and ToFZY5, YUCCA-like genes of tomato (Matsuo et al., 2018; Takisawa et al., 2019). These reports suggested that the IPyA pathway is the main IAA biosynthesis pathway in tomato ovaries and that AVG could reduce endogenous IAA levels in the ovaries of parthenocarpic tomatoes.
In this study, two experiments were conducted; we first examined the number of seeds in an F2 population from a cross between ‘Micro-Tom’ and ‘Renaissance’ to clarify the effect of the pat-2 gene on seed production. Subsequently, we investigated the effect of AVG on seed production in ‘Renaissance’ to develop a method to promote seed production in pat-2 parthenocarpic tomatoes.
‘Micro-Tom’, a non-parthenocarpic tomato cultivar, and ‘Renaissance’ (Sakata Seed Corporation, Yokohama, Japan), a parthenocarpic tomato cultivar that is homozygous for the pat-2 allele, were used. We developed a population of F1 (n = 3) and F2 plants (n = 89) derived from a cross between ‘Micro-Tom’ and ‘Renaissance’.
Plant growth conditionsTo investigate the effect of the pat-2 gene on seed production, we examined the seed number in F2 plants derived from a cross between ‘Micro-Tom’ and ‘Renaissance’ in spring and fall. In the spring examination, seeds of ‘Micro-Tom’, ‘Renaissance’, and F1 and F2 populations were sown on March 24, 2014. The seedlings were transplanted into containers in a greenhouse on April 19, 2014. In the fall examination, cuttings of F1 and F2 plants and ‘Renaissance’ were planted in a 128-cell tray filled with vermiculite, and ‘Micro-Tom’ seeds were sown on August 25 or 26, 2014. The seedlings were transplanted into containers in a greenhouse in September 15, 2014. To examine the effect of AVG on seed production, ‘Renaissance’ seeds were sown on March 21, 2013, and the seedlings were transplanted on April 20, 2013. All plants were grown in a greenhouse on an experimental farm at Kyoto University, Takatsuki (34°51' N, 135°37' E) in 2013 and 2014, respectively.
Effect of pat-2 on fruit weight and number of seeds, ovules, and loculesIn the spring and fall examinations, we pollinated from four to ten flowers within one or two flower clusters at anthesis. After maturation, we weighed the fruit and counted the number of seeds in spring and fall. In addition, in the spring cultivation, we sampled more than three young fruits from 13 to 44 days after anthesis, removed the pericarp of each young fruit, and counted the number of ovules. In the fall cultivation, we cut fruit in half along the equatorial plane and counted the number of locules.
Effect of AVG on seed productionWe used AVG (Sigma-Aldrich, St. Louis, MO, USA) for the experiment. At the first flower anthesis, we dipped a flower cluster in 0, 30, or 60 μM AVG solution. Between 39 and 43 flowers of ‘Renaissance’ were treated. We pollinated four flowers in each flower cluster at anthesis. After maturation, we weighed the fruit and cut them in half along the equatorial plane and counted the number of seeds in the distal and proximal parts of the fruits.
DNA extractionYoung leaves were sampled from the F2 plants and stored in a freezer at −40°C. DNA was extracted from the frozen leaves using a Nucleon PhytoPure kit (GE Healthcare Japan, Tokyo, Japan) according to the manufacturer’s instructions with some minor modifications.
PCR-based marker analysis to differentiate pat-2 allelesWe used primers for the marker that differentiates pat-2 alleles, according to Nunome et al. (2013) and Takisawa et al. (2017). The PCR reactions were performed in a total volume of 10 μL using BIOTAQTM DNA polymerase (Bioline, London, UK) according to the manufacturer’s instructions with some minor modifications. Amplification was carried out under the following conditions: 94°C for 10 min; 30 cycles of 94°C for 30 s, 58°C for 45 s, and 72°C for 30 s; and a final extension step at 72°C for 7 min. The PCR products were run on 1.0% agarose gels and stained with ethidium bromide.
Statistical analysisMean comparisons were performed using the Student’s t-test and Tukey’s test to examine differences among treatments at the 5% level of significance.
Nunome et al. (2013) reported that pat-2 is a zinc finger homeodomain (ZHD) protein gene. ZHD proteins function as transcription factors, possessing a homeodomain and a zinc-finger motif that contains cysteine and histidine residues (Windhovel et al., 2001). Although it has been reported that ZHD proteins are involved in regulating floral development in Arabidopsis (Tan and Irish, 2006), the function of pat-2 remains elusive. However, Fos et al. (2000) suggested that the parthenocarpy of pat-2 ovaries may be caused by a higher concentration of GA20, the precursor of an active GA. In addition, a study of MINI ZINC FINGER 1 (MIF1), a putative zinc finger protein from Arabidopsis, suggested that it may be involved in integrating signals from multiple plant hormones (Hu and Ma, 2006). These reports indicate the possibility that pat-2 is involved in fruit development by regulating metabolism of plant hormones.
In the F2 plants homozygous for the Pat-2 allele, heterozygous, and homozygous for the pat-2 allele, pollinated fruit weights were 19.7, 48.5, and 32.0 g in the spring examination and 17.1, 59.0, and 44.3 g in the fall examination, respectively (Table 1). The pollinated fruit weight of the plants homozygous for the Pat-2 allele was significantly lower than that of heterozygous plants in the spring examination, and lower than that of heterozygous plants and plants homozygous for the pat-2 allele in the fall examination (Table 1). This result suggested that pat-2 was involved in regulating fruit weight. However, it was reported that in an F2 population from ‘Micro-Tom’ and ‘MPK-1’, a quantitative trait locus (QTL) for pollinated fruit weight was detected on chromosome 4, where pat-2 is positioned (Takisawa et al., 2018). Although both ‘Micro-Tom’ and ‘MPK-1’ alleles at the pat-2 locus are homozygous for the Pat-2 allele, the QTL was detected. In addition, the fruit weight of plants homozygous for the ‘Micro-Tom’ allele at the QTL was significantly lower than that of plants with the other alleles, which is consistent with the result in the current study. These results suggested that the QTL for fruit weight was linked to the pat-2 locus, affecting pollinated fruit weight of plants with each allele at the pat-2 locus. Although we found that the position of the QTL was very close to that of the pat-2 locus in another experiment (unpublished data), further research is needed to clarify the reason for the pollinated fruit weight being lower in plants homozygous for the Pat-2 allele.

Pollinated fruit weight, number of seeds, ovules, and locules of each genotype at the pat-2 locus in the F2 population.
In the spring examination, the average seed numbers of ‘Micro-Tom’, ‘Renaissance’, and F1 plants were 21.8, 35.5, and 118.3, respectively (Fig. 1). In addition, the F2 population showed a continuous distribution and transgressive segregation in the number of seeds from 30.2 to 101.3 in plants homozygous for the Pat-2 allele, from 30.3 to 147.9 in heterozygous plants, and from 0.5 to 57 in plants homozygous for the pat-2 allele (Fig. 1). In the fall examination, distribution of the number of seeds in each genotype was similar to that of the spring examination. These results suggested that the number of seeds in the F2 population was regulated by multiple loci and that both parents harbored genes that regulate seed number.

Frequency distribution of number of seeds in the ‘Micro-Tom’, ‘Renaissance’, F1, and F2 populations distinguished by alleles of the pat-2 gene in the spring examination (n = 89).
The number of seeds was much lower in pat-2 parthenocarpic tomato cultivars than that in non-parthenocarpic tomato cultivars (Kataoka et al., 2008; Ohkawa et al., 2008). In this study, the number of seeds in plants homozygous for the pat-2 allele was significantly lower than that in heterozygous plants in the spring and fall examinations and that in Pat-2 homozygous plants in the spring examination (Table 1). These results showed that low seed set was genetically associated with the pat-2 allele. In addition, we detected no significant difference in the number of ovules and locules among plants with different genotypes (Table 1), suggesting that they were not related to low seed number in plants homozygous for the pat-2 allele. Furthermore, the number of seeds in heterozygous plants was significantly higher than not only that in pat-2 homozygous plants, but also that in Pat-2 homozygous plants. As described above, fruit weights of heterozygous plants were heavier than those of Pat-2 homozygous plants, suggesting that the increase in the number of seeds in heterozygous plants was caused by an increase in the fruit size.
AVG promotes seed production in ‘Renaissance’In the previous experiment, we showed that low seed set was genetically associated with the pat-2 allele. This indicated that low seed set was unavoidable when using the pat-2 gene to breed parthenocarpic tomato cultivars; therefore, it is important to develop a method to promote seed production in pat-2 parthenocarpic tomatoes.
In this study, we attempted to increase the number of seeds in ‘Renaissance’ using AVG. The average fruit weights following treatment with AVG at 0, 30, and 60 μM were 196.2, 177.2, and 169.3 g; treatment with AVG reduced the fruit weights, but there was no statistically significant difference among treatments (Fig. 2A). On the other hand, the average numbers of seeds following treatment with AVG at 0, 30, and 60 μM were 40.1, 81.3, and 43.3; treatment with AVG at 30 μM significantly increased the number of seeds compared with treatment at 0 or 60 μM AVG (Fig. 2B). Ohkawa et al. (2012) reported that 5 mg treatment with paclobutrazol, an inhibitor of GA biosynthesis, yielded significantly more seeds than the control in ‘Renaissance’. However, the number of seeds per fruit in the 25 mg treatment was significantly lower than that in the 5 mg treatment. This result suggests that there is an ideal concentration for the paclobutrazol treatment. In this study, although 30 μM AVG treatment increased the number of seeds compared with 0 μM AVG, 60 μM AVG treatment did not, which suggests that there is also an ideal concentration for AVG treatment.
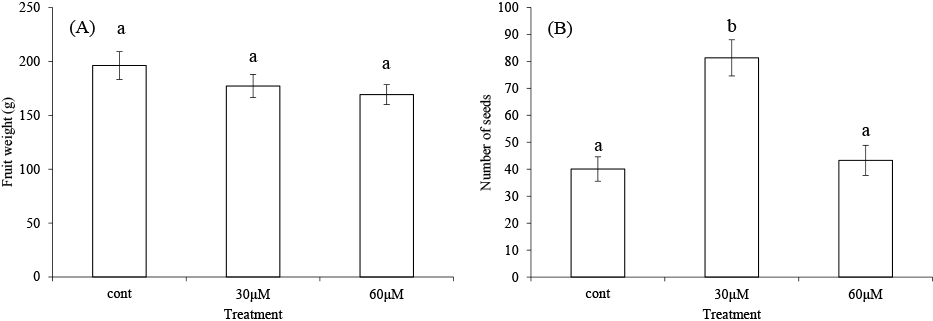
Effect of aminoethoxyvinylglycine (AVG) treatment (at 0, 30, or 60 μM) on (A) fruit weight and (B) number of seeds in ‘Renaissance’. Data show the mean ± SE (standard error). Different letters indicated significant differences by Tukey’s test at the 5% level.
In addition, the number of seeds was significantly lower in the proximal parts than in the distal parts of fruits in both the 0 and 60 μM AVG treatment groups (Fig. 3). In contrast, there was no significant difference in the number of seeds between the proximal and distal parts with the 30 μM AVG treatment. Takisawa et al. (2012) reported that pollen tubes did not reach the proximal half of fruits in ‘Renaissance’, causing low seed set in the proximal part of the fruit. In this study, although the 30 μM AVG treatment increased the seed number in both parts, it was greater in the proximal part; i.e., a 1.5-fold increase in the distal part was observed as compared to a 3.3-fold increase in the proximal one. Johkan et al. (2010) suggested that a high level of IAA inhibited the elongation of pollen tubes. Based on the results of Takisawa et al. (2012) and Johkan et al. (2010), the substantial increase in seed number in the proximal part with the 30 μM AVG treatment could be explained by reduced inhibition of pollen tube elongation due to the lower level of IAA in ‘Renaissance’ ovaries.

Effect of aminoethoxyvinylglycine (AVG) treatment (at 0, 30, or 60 μM) on the number of seeds in the distal and proximal parts of pollinated fruits in ‘Renaissance’. Data show the mean ± SE (standard error). Asterisks indicated significant differences between the distal and proximal parts by Student’s t-test at the 5% level.
Furthermore, we compared the effect of AVG on seed production at each flower position. There was almost no difference in seed number among flower positions following treatment with 0 or 60 μM AVG (Fig. 4). In contrast, with 30 μM AVG, the seed number of the flowers in second position in clusters was highest among all positions, although the difference was not statistically significant. The stages of second, third, and fourth-position flowers were about one, three, and five days before anthesis when we treated the flower cluster with AVG solution, respectively. Although the stages when AVG treatments were performed were different for each flower position, we did not observe any morphological changes in flowers at any position. In addition, the effect of AVG treatment was highest about one day before anthesis, just before pollen tubes elongated in the pistil. Therefore, it is possible that the AVG treatment about one day before anthesis reduced the concentration of IAA in the ovaries and promoted seed production by alleviating the inhibition of pollen tube elongation caused by a high concentration IAA.

Effect of aminoethoxyvinylglycine (AVG) treatment (at 0, 30, or 60 μM) on the number of seeds in pollinated fruits of ‘Renaissance’ at each flower position.
Recently, in addition to AVG and AOPP, several inhibitors of IAA biosynthesis have been discovered: L-kynurenine, 5-(4-chlorophenyl)-4H-1,2,4-triazole-3-thiol (yucasin), yucasin difluorinated analog, 4-biphenylboronic acid, and 4-phenoxyphenylboronic acid (He et al., 2011; Kakei et al., 2015; Tsugafune et al., 2017). It is anticipated that these chemicals will also promote seed production and we hope that future research will clarify the effect of these chemicals on the seed number in pat-2 parthenocarpic tomatoes.
In this study, we showed that low seed set was associated with the pat-2 allele. In addition, we did not detect a significant difference in the number of ovules or locules among plants with different alleles at the pat-2 locus, suggesting that they were not related to the low seed number in pat-2 parthenocarpic tomatoes. Moreover, we showed that treatment with 30 μM AVG could increase the number of seeds in the ‘Renaissance’ cultivar. Parthenocarpic tomato cultivars are beneficial in terms of reducing the labor needed for fruit production and maintaining stable fruit production under unfavorable conditions. Our research will contribute to the further development of pat-2 parthenocarpic tomatoes by identifying ways to increase seed production.