原著論文
Characterization of Parthenocarpic Fruit of ‘Miyazaki-wase No. 1’, a Tropical Squash (Cucurbita moschata L.) Cultivar
2021 年 90 巻 1 号 p. 68-74

詳細
2021 年 90 巻 1 号 p. 68-74
Parthenocarpy is a trait in which fruit set and development are induced without fertilization, thereby contributing to labor saving in fruit production. In Cucurbita species, parthenocarpic cultivars have been reported in Cucurbita pepo, but not in Cucurbita maxima and Cucurbita moschata. In this regard, the identification of parthenocarpic cultivars and clarification of their characteristics will contribute greatly to the research and breeding of the parthenocarpic trait in Cucurbita species. In this study, we aimed to genetically screen diverse Cucurbita accessions for high parthenocarpic ability and to elucidate the characteristics of the parthenocarpic cultivars discovered by the screening. We screened 33 accessions from squash genetic resources and found that a Cucurbita moschata cultivar, ‘Miyazaki-wase No. 1’, exhibited parthenocarpy. In order to clarify the characteristics, we examined the fruit set rate, fruit expansion, and fruit quality of ‘Miyazaki-wase No. 1’. In the spring and fall experiments, the fruit set rate of parthenocarpic fruit was lower than that of pollinated fruit. On the other hand, there was no significant difference in the transverse diameter and weight of parthenocarpic and pollinated fruits at harvest. These results indicated that the fruit set ability of parthenocarpic fruit is inferior to that of pollinated fruit, although the fruit expansion ability of parthenocarpic fruit was almost the same as that of pollinated fruit. In addition, we compared the composition of parthenocarpic fruit with that of pollinated fruit at harvest and after storage in the fall experiment. The results demonstrated that there was no significant difference in all the examined compositions between the parthenocarpic and pollinated fruits at harvest. On the other hand, the β-carotene concentration of parthenocarpic fruit was significantly lower than that of pollinated fruit one month and two months after harvest. These results indicated that parthenocarpic fruit development did not affect fruit quality at harvest, but had an effect on β-carotene metabolism during storage. We showed that ‘Miyazaki-wase No. 1’ is a parthenocarpic cultivar and clarified its characteristics. Based on the results, progress in research and breeding of parthenocarpic cultivars in Cucurbita species is anticipated in the future.
Cucurbita species are important fruit vegetables that supply carbohydrates, minerals, and vitamins. There are five domesticated species: Cucurbita argyrosperma, C. ficifolia, C. maxima, C. moschata, and C. pepo (Nee, 1990). Among them, C. maxima, C. moschata, and C. pepo are cultivated in fields or greenhouses in Japan according to regional characteristics and the season. Cucurbita species are monoecious crops that form male and female organs in the same individual. For fruit production, the female flower needs to be pollinated with pollen from the male flower via flower-visiting insects or artificial pollination. However, the pollen viability of C. maxima, C. moschata, and C. pepo drops sharply after anther dehiscence (Hayase, 1956; Nepi and Pacini, 1993). Therefore, farmers need to carry out artificial pollination in the early morning, which is very labor-intensive. In addition, pollination is inhibited when the stigma is wet by rain in field cultivation, lengthening the cultivation period and reducing fruit quality because of delayed harvest.
A promising way to overcome these problems is to take advantage of parthenocarpy. Parthenocarpy is a trait in which fruit set and development are triggered without pollination and fertilization, which reduces the financial and labor costs of fruit setting. In plants of the family Cucurbitaceae, use of the parthenocarpic trait is most prevalent in cucumber (Cucumis sativus L.) and it is considered a good model for the study of parthenocarpy. There are many genetic studies of parthenocarpy and they have suggested that the mode of genetic inheritance for parthenocarpy is quantitative in cucumber (Sun et al., 2006; Lietzow et al., 2016; Wu et al., 2016). In addition, Yoshioka et al. (2018) explored 172 accessions from an East Asian melon collection and identified accessions that exhibited strong parthenocarpic ability throughout the year. In Cucurbita species, although parthenocarpic cultivars of C. pepo were reported (den Nijs and Balder, 1983; Robinson, 1993; Robinson and Reiners, 1999; de Menezes et al., 2005; Martínez et al., 2014), there are no reports on the parthenocarpic cultivars of C. maxima and C. moschata. In addition, only a few studies focused on C. moschata, which could provide useful new traits. Identification of parthenocarpic cultivars and clarification of their characteristics will make a great contribution to the research and breeding of the parthenocarpic trait in Cucurbita species.
In this study, we firstly genetically screened diverse squash accessions, especially focusing on C. moschata, for high parthenocarpic ability. As a result, we found that ‘Miyazaki-wase No. 1’ (C. moschata L.), a local F1 cultivar in Miyazaki Prefecture, Japan, exhibited parthenocarpy. To elucidate the characteristics of parthenocarpy, we compared the fruit set rate, fruit expansion, and fruit morphology between pollinated fruit and parthenocarpic fruit. In addition, to clarify the fruit quality and storage traits of parthenocarpic fruit, we analyzed the composition of fruit at harvest and after storage.
A total of 33 squash accessions preserved in the Genebank of the National Agriculture and Food Research Organization (NARO) were used to screen parthenocarpic cultivars (Table S1). The seeds of all accessions were sown on April 25–27, 2016, and two seedlings from each accession were transplanted at 200 cm row length, spaced 80 cm, in a field located at the Kizu Experimental Farm of Kyoto University at Kizugawa, Japan (34°73' N, 135°84' E) on May 25, 2016. Compound chemical fertilizer (10 kg N, 21 kg P2O5, and 8 kg K2O per 10a) was applied as basal dressing.
The results of screening showed that a C. moschata F1 cultivar ‘Miyazaki-wase No. 1’ (JP83577), which was derived from a cross between the selected lines of ‘Jiki’ and ‘Futtsu-wase’ exhibited parthenocarpy. In order to clarify its characteristics, we performed investigations in fall 2018 and spring 2019 using seeds of the same cultivar that were stored at the Miyazaki Agricultural Experiment Station. Seeds were sown in 10.5 cm plastic pots filled with a mixture of bark compost, decomposed granite soil, and smoked rice hulls (2:2:1, v/v/v) on August 29, 2018 in the fall experiment and April 3, 2019 in the spring experiment. In the fall and spring experiments, 49 and 20 seedlings were transplanted in a greenhouse at the Kizu Experimental Farm, respectively. The plants were planted at 300 cm row length, spaced 30 cm, on September 20 and 21, 2018 in the fall experiment and on April 30, 2019 in the spring experiment. We carried out irrigation, fertilization, and pest control according to standard procedures. In the experiments, the main shoot was trained horizontally without topping, and lateral buds on the lower nodes that bore fruit were removed. During the experimental period in fall 2018 and spring 2019, the temperatures were measured by a data logger (TR-71wb; T&D Corporation, Matsumoto, Japan).
Evaluation of parthenocarpyIn the screening, we used two plants from each accession. One or more female flowers of each plant were tied with hemp twine to prevent pollination one day before anthesis in June 2016. The fruits were harvested in early August 2016 and checked for the presence of seeds. We judged accessions that formed at least one seedless fruit as parthenocarpic cultivars.
No pollination and pollination treatmentsIn the experiments, we tied the first female flower on the main shoot with hemp twine to prevent pollination one day before anthesis. This was followed by pollination and no pollination treatments. In the case of pollination treatment, hemp twine was removed at anthesis and the female flower was pollinated artificially early in the morning. In the no pollination treatment, the flower was kept tied with hemp twine after anthesis to avoid pollination. In addition, in plants whose first female flower did not set, we performed the same treatment on the second female flower. In the fall experiment, first female flowers and second female flowers were formed from the 13th to the 20th node and from the 19th to the 28th node of the main stem, respectively. In the spring experiment, first female flowers and second female flowers were formed from the 13th to the 15th node and from the 17th to the 21st node of the main stem, respectively.
Characterization of parthenocarpy in ‘Miyazaki-wase No. 1’To characterize parthenocarpy in ‘Miyazaki-wase No. 1’, we used 9–10 plants in which pollination or no pollination treatment was performed in October 2018 in the fall experiment and on May 2019 in the spring experiment. Forty days after anthesis, we examined the fruit set rates of non-pollinated and pollinated flowers and harvested the fruits. After harvest, we measured fruit weight and transverse diameter and flesh (mesocarp) thickness at the equatorial plane in both experiments, and fruit height in the spring experiment. In addition, we weighed the seeds and placenta, and calculated their percentage to total fruit weight in the fall experiment. We also compared the size of the parthenocarpic and pollinated fruits by measuring fruit transverse diameter with electronic calipers at 0, 10, 20, 30, and 40 days post-anthesis in both experiments.
Preparation for fruit component analysis and measurement of water content and total soluble solids (TSS)To evaluate parthenocarpic fruit quality at harvest and after storage, we used eight or nine pollinated and eight parthenocarpic fruits at each storage stage: at harvest and after 1-month and 2-month storage in the fall experiment. The storage conditions were 10°C at 30% relative humidity. We examined water content, TSS, sugar content (fructose, glucose, and sucrose), β-carotene, and ascorbic acid at harvest, and TSS and β-carotene after storage. We cut the fruit into small pieces. After approximately 5 g of sample was dried at 80°C for four days, the dry weight was measured. From the fresh weight and the dry weight, we calculated the water content in the fruit. TSS was measured using a digital refractometer (PAL-1; Atago Co., Ltd., Tokyo, Japan) after grating the sample.
Sugar analysisAn approximately 5 g sample was frozen using liquid nitrogen and disrupted with a MULTI-BEADS SHOCKER® (Yasui Kikai Corporation, Osaka, Japan). After disruption, sugars were extracted with 10 mL of 80% (v/v) ethanol at 80°C for 10 min, followed by centrifugation at 4000 rpm at 4°C for 10 min. After repeating the extraction procedure four times, the volume was fixed at 50 mL with measuring flasks. The extracted solution was filtered through a 0.2 μm Minisart Syringe Filter (Sartorius AG, Goettingen, Germany). Glucose, fructose, and sucrose were determined by high-performance liquid chromatography (HPLC) using a Shodex Asahipak NH2P-50 4E column (Showa Denko K. K., Tokyo, Japan). The mobile phase was 75% (v/v) acetonitrile/water at the flow rate of 1 mL/min at 40°C. Quantification of sugars was performed by comparison with external standards.
β-carotene analysisApproximately 5 g of frozen sample was disrupted with a MULTI-BEADS SHOCKER® (Yasui Kikai). After disruption, β-carotene was extracted with 7.5 mL of 100% acetone at room temperature for 30 min using a shaker (R-10 mini; TAITEC CORPORATION, Saitama, Japan), followed by centrifugation at 4000 rpm at 4°C for 10 min. After repeating the extraction procedure five times, the volume was fixed at 50 mL with measuring flasks. The extracted solution was filtered through a 0.2 μm Minisart Syringe Filter (Sartorius AG, Goettingen, Germany). β-Carotene was determined by HPLC. Separation of carotenoids was performed in a Kinetex C18 column (150 × 4.5 mm) (Phenomenex, Le Pecq, France) by gradient elution of acetonitrile and ethanol. The elution started with a mixture of 90% acetonitrile and 10% ethanol, which was changed to 10% acetonitrile and 90% ethanol at 10 min and returned to the initial condition at 14 min. The flow rate was 1 mL·min−1. Quantification of β-carotene was performed by comparison with external standards.
Ascorbic acid analysisAscorbic acid content was determined with a reflectometer (RQflex® 20; Merck KGaA, Darmstadt, Germany) and an ascorbic acid test (Reflectoquant ascorbic acid test; Kanto Chemical Co., Inc., Tokyo, Japan) according to the manufacturer’s instructions.
Statistical analysisStatistical analysis of the data was performed using Welch’s t-test to determine differences between treatments at 1% and 5% levels of significance.
We genetically screened 33 squash accessions for high parthenocarpic ability to identify parthenocarpic cultivars of C. maxima and/or C. moschata. As a result, we found that a C. moschata cultivar, ‘Miyazaki-wase No. 1’, formed seedless fruits among the examined cultivars and judged this cultivar to be a parthenocarpic cultivar.
Parthenocarpic ability of ‘Miyazaki-wase No. 1’To clarify the parthenocarpic ability of ‘Miyazaki-wase No. 1’, we examined the fruit set rate and development of parthenocarpic fruit in fall and spring experiments. In the fall experiment, the fruit set rates of the first flowers were 3/9 and 10/10 in the no pollination and pollination treatments, respectively (Table 1). In the no pollination treatment, six plants did not bear fruits, so we performed no pollination treatment on the second flowers of those plants. As a result, all the plants bore parthenocarpic fruit. In the spring experiment, the fruit set rates of the first flowers were 0/10 and 9/10 in the no pollination and pollination treatments, respectively (Table 1). We performed pollination or no pollination treatment with the second flowers of the plants that did not bear fruit. In the pollination treatment, one plant bore pollinated fruit. On the other hand, in the no pollination treatment, four of 10 plants bore parthenocarpic fruit. The fruit set rates of the first flowers in the pollination treatment were 10/10 and 9/10 in the fall and spring experiments, respectively. The fruit set rates were much higher than those of parthenocarpic fruit, which indicated that the ability to set parthenocarpic fruits was lower than that to set pollinated fruits.
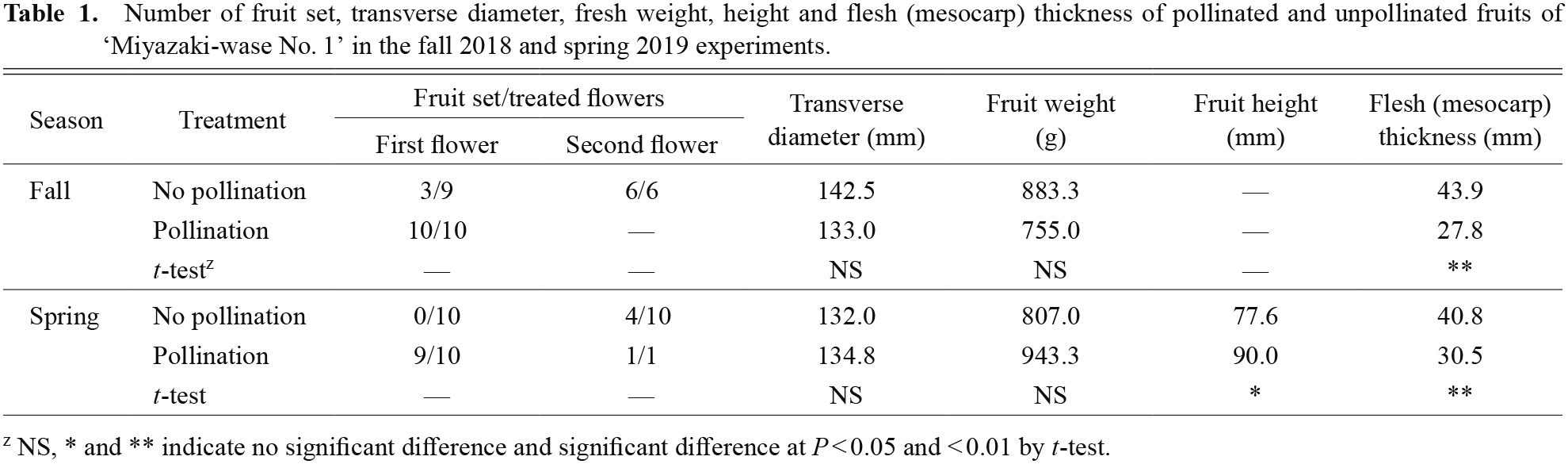
Number of fruit set, transverse diameter, fresh weight, height and flesh (mesocarp) thickness of pollinated and unpollinated fruits of ‘Miyazaki-wase No. 1’ in the fall 2018 and spring 2019 experiments.
The fruit set rate of the first flowers in the no pollination treatment was higher in the fall experiment than the spring experiment. It is known that the low temperatures in winter promote parthenocarpy in C. pepo (Globerson, 1971; Rylski, 1974; den Nijs and Balder, 1983; Rylski and Aloni, 1990; Robinson and Reiners, 1999). Figure 1 shows the daily mean temperature from transplanting to the end of harvest in the 2018 fall and 2019 spring experiments. The daily mean temperature decreased after transplanting in 2018 fall, while it increased in 2019 spring. From 20 days after transplanting, the daily mean temperature was always lower in 2018 fall than 2019 spring. These results showed that the low temperature in fall may increase the ability of ‘Miyazaki-wase No. 1’ to set parthenocarpic fruits. Furthermore, in both the fall and spring experiments, the fruit set rates of the first flowers were lower than those of the second flowers in the no pollination treatment. Fruit set at low nodes only has a few leaves that produce photosynthetic products, which can lead to nutrient competition between fruit formation and vegetative growth. In cucumber (Cucumis sativus), fruits and vegetative organs competed for photoassimilates and the unequal distribution of photoassimilates among fruit and vegetative organs caused fruit abortion (Schapendonk and Brouwer, 1984; Marcelis, 1992; Hikosaka and Sugiyama, 2015). These reports supported the hypothesis that nutrient competition reduced the fruit set rates in the first flowers of ‘Miyazaki-wase No. 1’.

Daily mean temperature in the fall 2018 and spring 2019 experiments.
Next, the characteristics of fruit expansion and fruit morphology between pollinated and parthenocarpic fruits were investigated. During fruit development, there was no significant difference in the transverse diameter of the parthenocarpic fruit and the pollinated fruit in the fall and spring experiments (Fig. 2). In addition, the transverse diameter and fruit weight did not show significant difference between parthenocarpic and pollinated fruits at harvest (Table 1). These results showed that the ability to enlarge of parthenocarpic fruits was the same as that of pollinated fruits. On the other hand, fruit height in the spring experiment was significantly higher in pollinated fruit than parthenocarpic fruit, and flesh (mesocarp) thickness in both the spring and fall experiments was significantly greater in parthenocarpic fruit than pollinated fruit (Table 1). Furthermore, Figure 3 shows that the seed and placenta parts of parthenocarpic fruit were smaller than those of pollinated fruit. These results demonstrate that parthenocarpy affected fruit morphology. In watermelon (Citrullus lanatus), parthenocarpic fruits produced by pollination with bottle gourd pollen affected fruit morphology, and there was a significant difference in the ratio of fruit length to fruit width (Sugiyama et al., 2014). Although the reason remains unclear, fruit development depends on an increase in cell number and cell expansion. In the future, histological analysis is expected to clarify the cause of the difference in fruit morphology between pollinated and parthenocarpic fruits.

Development of pollinated and parthenocarpic fruits of ‘Miyazaki-wase No. 1’ in the fall (A) and spring (B) experiments. Values are means of 4–10 fruits ± SE.
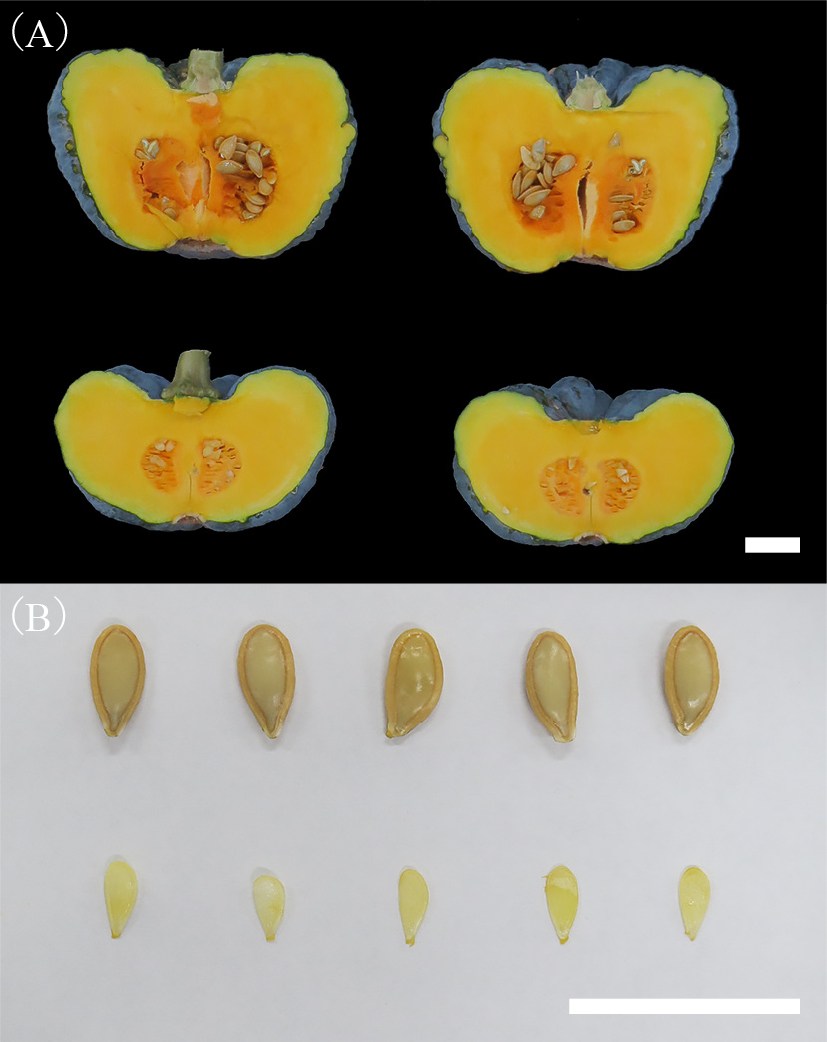
Fruits and seeds of ‘Miyazaki-wase No. 1’ in the 2019 spring experiment. (A) Longitudinal sections of pollinated fruit (upper) and parthenocarpic fruit (lower), and (B) seeds of pollinated fruit (upper) and empty seeds of parthenocarpic fruit (lower). Scale bars: 3 cm (A and B).
Parthenocarpy can produce seedless fruit, which is preferred by consumers and processors, because it reduces labor and processing costs. In this study, we showed that empty seeds composed of only thin seed coats were produced in parthenocarpic fruit of ‘Miyazaki-wase No. 1’ (Fig. 3). The empty seeds were smaller than seeds of pollinated fruit, and the empty seeds are less troublesome than normal seeds. In addition, the percentage weight of seeds and placenta to the total fruit weight was 9.1% in pollinated fruits, while it was only 1.8% in parthenocarpic fruits. This result showed that edible part was larger in parthenocapric fruits than pollinated fruits when they were the same weight.
Nutritional composition of ‘Miyazaki-wase No. 1’ parthenocarpic fruitCucurbita species are important vegetables containing carbohydrates, minerals, and vitamins. In addition, although many studies investigated parthenocarpy in various crops, few studies compared the quality of pollinated fruits and parthenocarpic fruits. Therefore, we examined water content, TSS, sugar, β-carotene, and ascorbic acid contents in pollinated and parthenocarpic fruits of ‘Miyazaki-wase No. 1’ in the fall experiment. We found no significant difference in composition between pollinated and parthenocarpic fruits (Table 2), suggesting that parthenocarpic fruit development did not affect nutritional composition at harvest. In cucumber, naphthaleneacetic acid (NAA) and N-(2-chloro-4-pyridyl)-N′-phenylurea (CPPU) treatment induced parthenocarpic fruits. NAA treatment did not affect the nutritional characteristics, while CPPU treatment decreased the levels of phenolic acid and ascorbic acid (Qian et al., 2018). This report suggested that the factor that induced parthenocarpy affected the nutritional characteristics of parthenocarpic fruits. An investigation into the relationship between plant hormones and parthenocarpy in ‘Miyazaki-wase No. 1’ and a clarification of the effect of plant hormones on nutritional characteristics of parthenocarpic fruits in C. moschata could promote an understanding of the mechanism regulating the nutritional characteristics of parthenocarpic fruits.
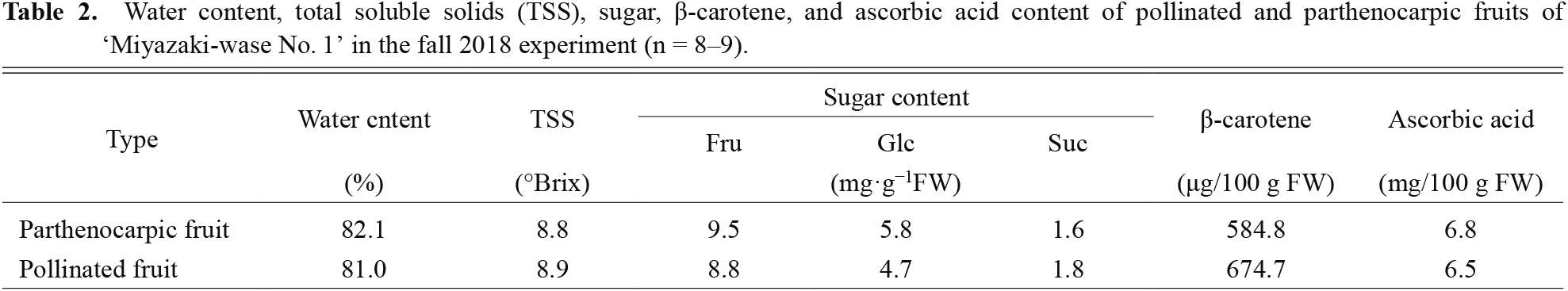
Water content, total soluble solids (TSS), sugar, β-carotene, and ascorbic acid content of pollinated and parthenocarpic fruits of ‘Miyazaki-wase No. 1’ in the fall 2018 experiment (n = 8–9).
To examine the change in fruit composition during storage, we examined β-carotene and TSS in pollinated and parthenocarpic fruits one month and two months after harvest. It is known that sugar and β-carotene contents increased in Cucurbita species during storage (Phillips, 1946; Kami et al., 2011). The increase in sugar content was caused by the conversion of starch into sugar. In this study, TSS increased in pollinated and parthenocarpic fruits during storage, and there was no significant difference in TSS between them. On the other hand, although the β-carotene content also increased in both fruits during storage, the content in parthenocarpic fruit was significantly lower than that in pollinated fruit one month and two months after harvest (Fig. 4). Zhang et al. (2014) reported that carotenoid accumulation is promoted by the conversion of amyloplasts into chromoplasts in winter squash, and the carotenoid enhancement seems to be due to continuous biosynthesis along with reduced turnover and/or enhanced sequestration. Therefore, a study of the conversion process of amyloplasts to chromoplasts or comparing the structure of amyloplasts and chromoplasts in parthenocarpic fruits will clarify the reason for the difference in β-carotene content between the parthenocarpic and pollinated fruits of ‘Miyazaki-wase No. 1’.

Total soluble solids (TSS) and β-carotene contents in pollinated and parthenocarpic fruits of ‘Miyazaki-wase No. 1’ during storage in the 2018 fall experiment. We used eight or nine pollinated fruits and eight parthenocarpic fruits in each storage stage. Values are means of eight–nine fruits ± SE. Asterisk indicates significant difference between pollinated and parthenocarpic fruits at each time point. (t-test; P < 0.05).
In conclusion, we found that ‘Miyazaki-wase No. 1’ exhibited parthenocarpy. As far as we know, ‘Miyazaki-wase No. 1’ is the first parthenocarpic cultivar reported in C. moschata. In addition, we clarified the characteristics of the parthenocarpic fruit. Parthenocarpic fruit development does not have an adverse effect on fruit expansion or quality. Therefore, ‘Miyazaki-wase No. 1’ is an attractive source for breeding parthenocarpic cultivars of the Cucurbita species. In Japan, C. maxima is the most important Cucurbita species and the introduction of the parthenocarpic trait into C. maxima is desirable. Further research on the mechanism of parthenocarpy and the identification of parthenocarpic genes is necessary to promote the development of parthenocarpic cultivars.
We thank the vegetable department of Miyazaki Agricultural Experiment Station and the Genebank of NARO for providing ‘Miyazaki-wase No. 1’ seeds and germplasm of squashes, respectively.