原著論文
Effect of Cold Exposure on Brix Value of Borecole Leaves Grown in an Unheated Greenhouse in Hokkaido, Subarctic Region
2023 年 92 巻 2 号 p. 171-177

詳細
2023 年 92 巻 2 号 p. 171-177
Borecole (curly kale) was cultivated without heating during autumn and winter in a multi-film-covered greenhouse in Hokkaido, Japan. Borecole survived even when the greenhouse air temperature dropped to −6.2°C with an accompanying drastic increase in Brix value. This rapid increase of Brix value was observed under conditions where the minimum air temperature in the greenhouse was less than 0°C. Cold temperatures caused the plant to freeze, inducing extracellular freezing. Although the Brix value increased at low temperatures irrespective of leaf order, the Brix values for leaves of lower order, at the top of the plant, were much larger than the higher order leaves, closer to the base of the plant. Leaf Brix values started to increase when the average minimum temperature was 1.3°C or less for 10 days before harvest. Moreover, the nitrate concentration, reported to correlate with bitterness intensity, decreased at low temperatures. These results revealed that leaves with reduced bitterness and improved sweetness can be produced in low temperatures from the upper portions of borecole plants.
Most of Hokkaido, the northernmost region of Japan, is categorized as subarctic (Köppen, 1936) with close to the heaviest snowfall in the world. Few leafy vegetables are cultivated there in winter (Hokkaido Central Union of Agricultural Corporations and HOKUREN Federation of Agricultural Cooperatives, 2021). During the Hokkaido mid-winter, Jishi and Takahama (2021) cultivated komatsuna (Japanese mustard spinach, Brassica rapa var. perviridis) in an unheated greenhouse of multiple polyolefin (PO) film layers. Their results showed that despite a lowest outdoor air temperature of −26.8°C, the minimum greenhouse temperature was −9.1°C, demonstrating komatsuna cultivation in the Hokkaido winter without heating. Unheated greenhouses in Hokkaido can also support the cultivation of lettuce (Takahama, 2021), turnip, green pak choi (Takahama et al., 2021a), purple mizuna, and leaf mustards (Takahama et al., 2021b). Identifying additional leafy vegetables for winter cultivation without heating is economically and nutritionally beneficial. As such, borecole is expected to be a suitable winter production vegetable.
Among the Brassica vegetables, Kale (Brassica oleracea var. acephala L.) has the greatest cold stress tolerance (Altinok and Karakaya, 2003). It is often cultivated in open fields from autumn to winter in northern Europe. Borecole, a type of kale, is also called curly kale for its unique curled leaf appearance (Dixon, 2006). Borecole is commonly eaten in Europe (Laska and Zelenkova, 1988) and is recognized as a “super-food” for its high functional nutrient content (Sikorska-Zimny and Beneduce, 2021). Even in Japan, borecole cultivation is increasing due to its excellent nutritional benefits and good taste.
Jurkow et al. (2019) cultivated borecole plants in an unheated greenhouse in Poland from autumn to winter and reported a temperature of approximately −5°C increased sugar and nutritional component content. As sweet vegetables are popular in Japan, the sugar concentration of borecole leaves is an important indicator of commercial value. Therefore, borecole cultivation methods in Hokkaido yielding high sugar content crops have great economic potential in Japan.
We specifically examined the Brix value of borecole to assess the sugar content in vegetables, grown in an unheated greenhouse in Hokkaido from autumn to winter. We also measured changes in nitrate concentration, which is reported to correlate with bitterness intensity (Noda and Makuta, 2015). As borecole is often shipped as individual leaves, it is also necessary to study the sugar concentration as a function of leaf position on the plant. There are currently, no reports describing this relationship.
Therefore, we investigated Brix value as a function of leaf position. Also, we evaluated the relationship between temperature and leaf Brix value variation in the individual plant to evaluate the impact of cold exposure during winter.
Experiments were conducted in an unheated plastic greenhouse (235.1 m2 area, 3.3 m building height, 365.4 m3 volume) located at the Hokkaido Research Organization, Pippu, Hokkaido, Japan (43.9°N, 142.5°E). The greenhouse is depicted in Fig. 1. To improve heat retention, an air-inflated double-layer plastic greenhouse was used. The roof film was double stretched with a constant inter-layer airflow supplied by a blower (Iwasaki et al., 2011; Roberts, 1978). The research conducted over winter 2017 and 2018, and in spring 2018, investigated Brix value changes in each leaf on a plant. In winter 2018 and 2019, changes in the leaf dry matter ratio were assessed. A movable PO film curtain was installed on November 4, 2017, November 23, 2018, and November 15, 2019. The curtain opened at 8:30 and closed at 16:30. The side window opened with an automatic ventilation system (Kurufami Ace III; Seiwa Co., Ltd., Shimotsuke, Japan) when the air temperature was over 25°C. The air temperature at 20 cm above the ground in the greenhouse was recorded hourly using a temperature data logger (RTR-502; T&D Co., Nagano, Japan). Additionally, in winter 2019, the greenhouse soil temperature at 10 cm below the ground surface was recorded hourly. The outdoor air temperature recorded by Automated Meteorological Data Acquisition System (Pippu, 43.5°N, 142.3°E) of the Japan Meteorological Agency was used for this study.

Unheated greenhouse used in the experiments.
Borecole ‘winterbor’ (Bejo Japan Co., Sapporo, Japan) plants were used for the study. The sowing dates were August 26, 2017, April 6, 2018, August 24, 2018, and August 24, 2019, for the winter 2017 experiment, spring 2018, winter 2018, and winter 2019 experiments, respectively. Seeds were sown in 128-cell plug trays with a commercial substrate (Pottobaido; Hokusan Co., Ltd., Kitahiroshima, Japan). Seedlings were grown for 19, 21, 21, and 25 days, in each experiment described above respectively, and transplanted to the greenhouse. The planting density was 6.3 plants·m−2. Fertilizer (N, P, K = 18, 6, 13 g·m−2) was applied before planting. Irrigation was conducted according to the method established by Jishi and Takahama (2021).
Harvest and Brix measurementIn winter 2017, six harvests were conducted from October 31 to December 20, 2017. In spring 2018, the same investigation was performed four times from May 22 to June 11. In each harvest, six plants were randomly sampled from the greenhouse, and the individual plant investigated for plant growth and Brix values as follows. Non-yellowing and undamaged fresh leaves 10 cm or longer in length were collected from each sampled plant (Fig. 2). The number of leaves was counted simultaneously. Fresh weights of all leaves were measured. To measure the Brix value, individual fresh leaves were placed in a polyethylene bag, frozen in a freezer at −30°C, then after thawing at 20°C crushed with a mallet to obtain juice; the juice was used to measure Brix value with a refractometer (PR-101α; Atago Co., Ltd., Tokyo, Japan). The average leaf Brix value was calculated by multiplying the Brix value by fresh leaf weight for each leaf order, then dividing the sum of the obtained values by the total leaf fresh weight.

Borecole plant separated by leaf order.
In winter 2018, on each sample day, six plants were randomly sampled on October 29, November 28, and December 26, with the individual plant growth, Brix values, and nitrate concentration evaluated as follows. After counting the number of leaves 10 cm or longer in length in each plant, all leaves were cut longitudinally in half. Half of each leaf was used for the Brix value measurement. The other half was placed in a paper bag for evaluation of the fresh leaf dry matter ratio and nitrate concentration. The Brix values were evaluated as per the 2017 experiment. To measure the dry matter ratio, the leaves were placed in an automatic oven set at 70°C. After 72 h or more, the leaf samples were weighed. The dry matter ratio was calculated by dividing the dried leaf weight by the fresh leaf weight. Nitrate concentration was measured using samples prepared for the dry matter ratio measurements. The nitrate concentration measurement method followed that used for forage (Okamoto et al., 2007). To 2 g of the dried and powdered sample was added 100 mL of distilled water. This mixture was shaken for 30 min to extract nitrates. Then the nitrate concentration in the filtrate was measured using RQ flex (Merck KGaA Co., Darmstadt, Germany).
In winter 2019, six plants were randomly sampled from the greenhouse seven times in all from October 25, 2019, to January 10, 2020, and the individual plant with the number of fresh leaves, leaf fresh weight, nitrate concentration, and dry matter ratio evaluated using the same method as that used for the winter 2018 experiment.
To identify optimal environmental conditions to increase Brix value, the relationship between Brix value and average minimum air temperature for 10 days before harvest was investigated using the winter-cultivated plants in 2017 and 2018.
StatisticsThe Tukey-Kramer test and Dunnett’s test were used for statistical analysis. Significant difference was inferred for Ρ < 0.05. Statistical software was used (Excel statistics ver. 7.0; Esumi Co., Ltd., Tokyo, Japan). The dry matter ratio was evaluated using the Tukey-Kramer test after performing an arcsine transformation.
Each year, the winter air temperature inside the greenhouse decreased (Fig. 3A, C, D). After November 30 in 2017, November 20 in 2018, and December 3 in 2019, the daily minimum greenhouse air temperature was below 0°C on most days. The minimum greenhouse air temperatures during the cultivation period for each year were −2.2°C, −2.9°C, and −6.2°C in 2017, 2018, and 2019, respectively. During the cultivation period, the maximum greenhouse air temperatures exceeded 0°C on most days. Over winter the soil temperature in 2019 decreased, with each day after November 7 falling to below 10°C. During the winter cultivation periods of 2017, 2018, and 2019, the freezing of plants was observed in the early morning when the daily minimum air temperature in the greenhouse fell below 0°C. However, the frozen leaves recovered with the daytime rise in air temperature (Fig. 4). During the spring cultivation period of 2018, the greenhouse air temperature rose over time (Fig. 3B). For this period, the minimum greenhouse temperature was 4.5°C on May 20, 2018.
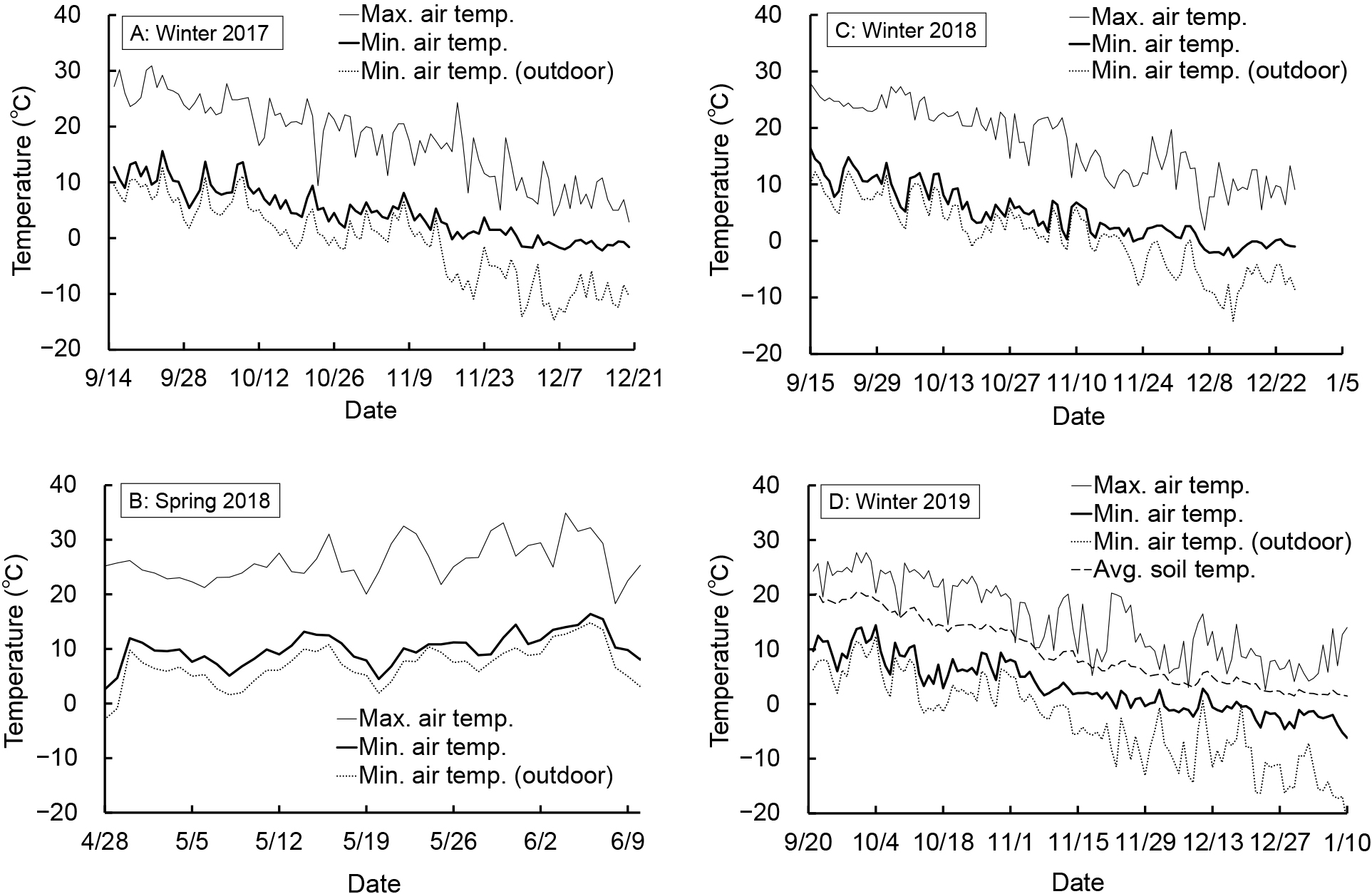
Transition of greenhouse daily maximum and minimum air temperature (A, B, C, and D), average soil temperature (D), and the minimum outdoor air temperature (A, B, C, and D) for each cultivation.

Borecole leaf freezing and recovery after frost exposure on December 20, 2019. A, early morning (7:00 AM); B, noon.
During the winter 2017 cultivation period, the average borecole leaf Brix value significantly increased after November 14. The Brix value on December 20 was 11.2 °Bx, 204% the November 14 value (Fig. 5A). By November 28 there was a significant increase in the number of fresh leaves. After this point the number of leaves remained steady at approximately 16. Strong negative correlations were observed between the leaf order from the top to the bottom and the leaf Brix value on November 14, 2017 and December 20, 2017 (Fig. 6A). The leaf Brix values of the lower order leaves, from the top of the plant, were greater than those of higher leaf order, closer to the ground. The leaf Brix values on December 20, 2017 were all larger than those on November 14, 2017, irrespective of leaf order.
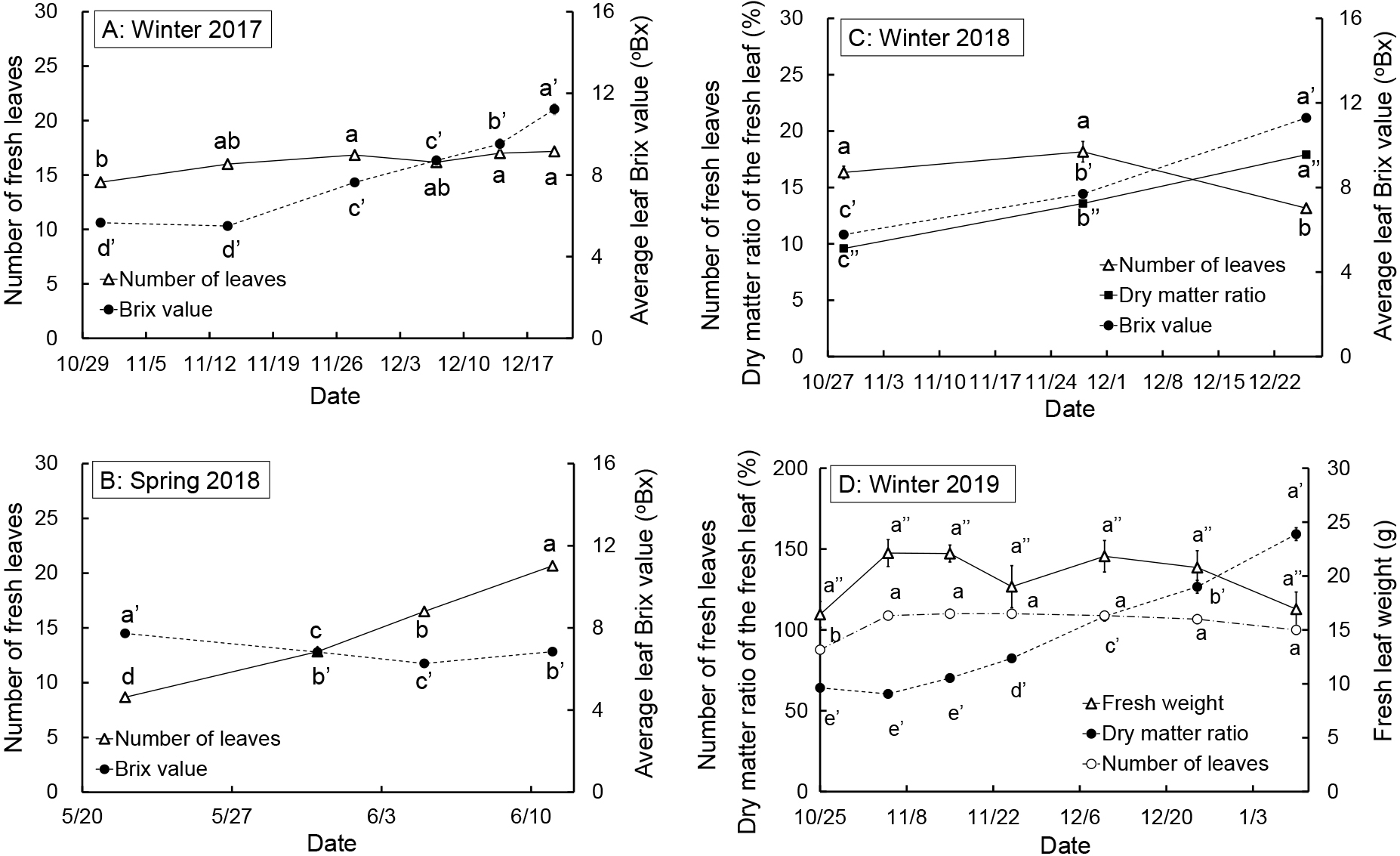
Change in the number of fresh leaves, average Brix values (A, B, and C), dry matter ratios (C, D), and fresh leaf weight (D) for borecole plants in each cultivation. Different letters represent significant differences among dates per item investigated at P < 0.05 by Tukey-Kramer’s test (n = 6); a–e: number of leaves, a′–d′: Brix value, a″–c″: dry matter ratio (C) or fresh weight (D). Vertical bars represent standard error.
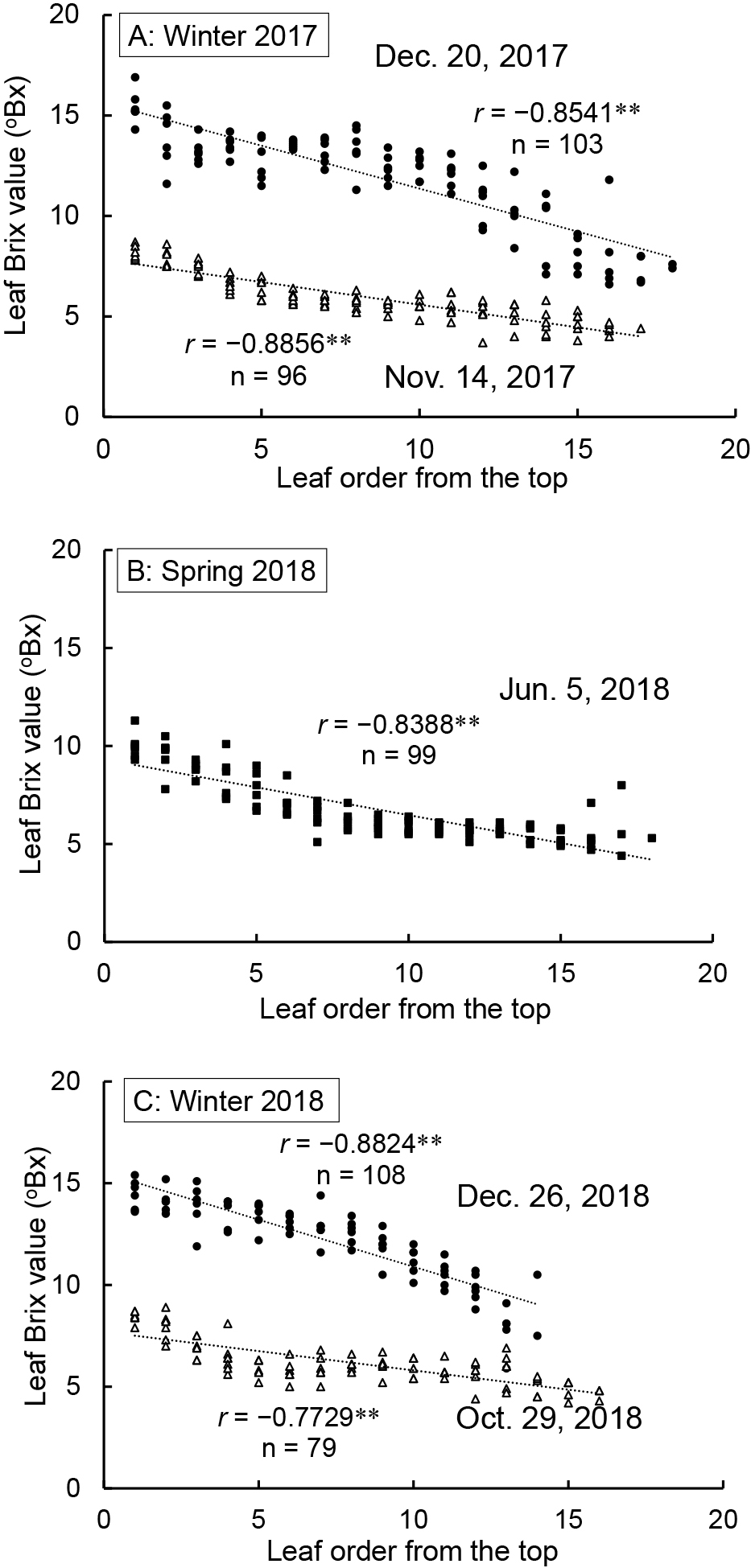
Relationship between leaf order from the top and Brix value in borecole plants of each cultivation. Smaller leaf order corresponds to closer to the plant top, while larger numbers are closer to the base of the plant. ** denotes a significant negative correlation between leaf order and Brix value (P < 0.01).
The average leaf Brix value in spring 2018 slightly decreased over time (Fig. 5B). The lowest leaf Brix value was 6.5 °Bx on June 5 during cultivation. In contrast, the number of fresh leaves increased, reaching 16.5 on June 5, 2018, roughly the same number of fresh leaves as from November 28, 2017 to December 20, 2017 (Fig. 5A, B). The leaf order showed similar behavior as that observed during winter cultivation. A strong negative correlation was also identified between the leaf order and the leaf Brix value on June 5, 2018 (Fig. 6B). The Brix values of the lower order leaves, from the top of the plant, were greater than those with a higher leaf order, from the bottom of the plant.
Winter 2018In winter 2018, there were significantly fewer fresh leaves on December 26 than on October 29 and November 28 as the higher order leaves, from the lower section of the plant, were wilted or yellowed (Fig. 5C). Both the average leaf Brix value and dry matter ratio of fresh leaves significantly increased over time. The Brix value and dry matter ratio on December 26 were, respectively, 205% and 187% of those on October 28. As in the 2017 experiment, the leaf Brix values for leaves of lower order, from the top of the plant, were greater than those for leaves of larger orders, from the lower plant section, on October 29 and December 26 (Fig. 6C). The fresh leaf nitrate concentrations decreased considerably over time (Fig. 7) with the nitrate concentration on December 26 only 24% of the October 28 value.
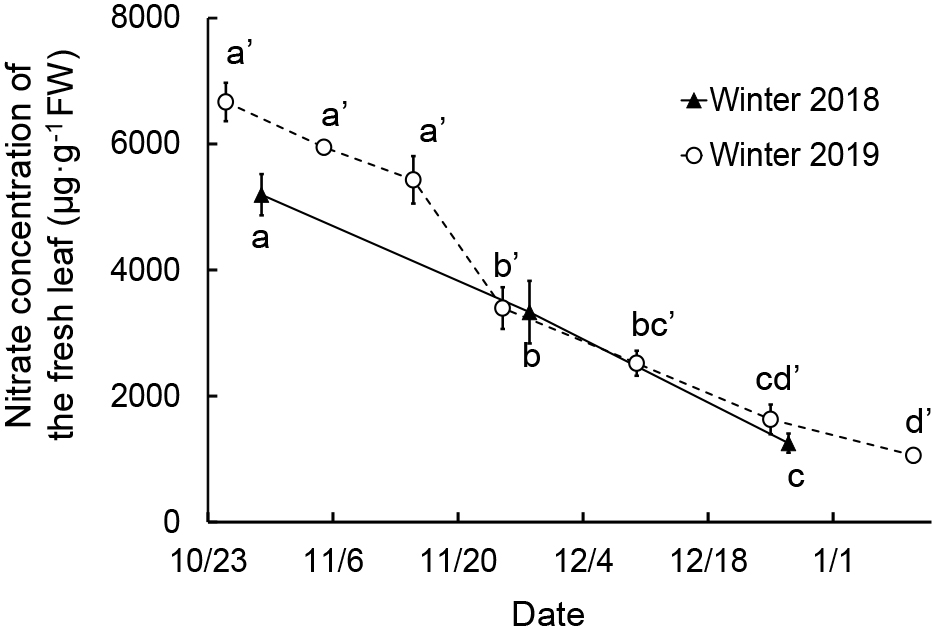
Change in leaf nitrate concentration of borecole plants in each cultivation. Different letters represent significant differences among dates per year investigated at P < 0.05 by Tukey-Kramer’s test (n = 6); a–c: winter 2018, a′–d′: winter 2019. Vertical bars represent the standard error.
No significant difference was found in the number of fresh leaves and the fresh leaf weight after November 5, 2019. The number of fresh leaves was approximately 16, as in the experiment of winter 2017 (Fig. 5D). The dry matter ratio significantly increased after November 14. The nitrate concentration of fresh leaves decreased significantly over time as in the 2018 experiment. The nitrate concentration on January 10 was 16% of that on October 25 (Fig. 7).
Relation between the leaf Brix value and the average minimum air temperatureUntil the middle of November, the Brix values in leaves harvested in 2017 and 2018 were relatively low, less than 6, however, after late November the Brix values of harvested leaves increased (Fig. 5). The leaf Brix values did not increase with an average minimum air temperature of 2.5°C (arrow in Fig. 8, 5.8 in Brix value). However, a dramatic increase occurred coinciding with a drop in the average minimum air temperature to below 1.3°C. All plots (harvests) yielded significantly higher than 5.8 °Bx at 2.5°C by Dunnett’s test, with some harvests yielding Brix values close to 12 °Bx.
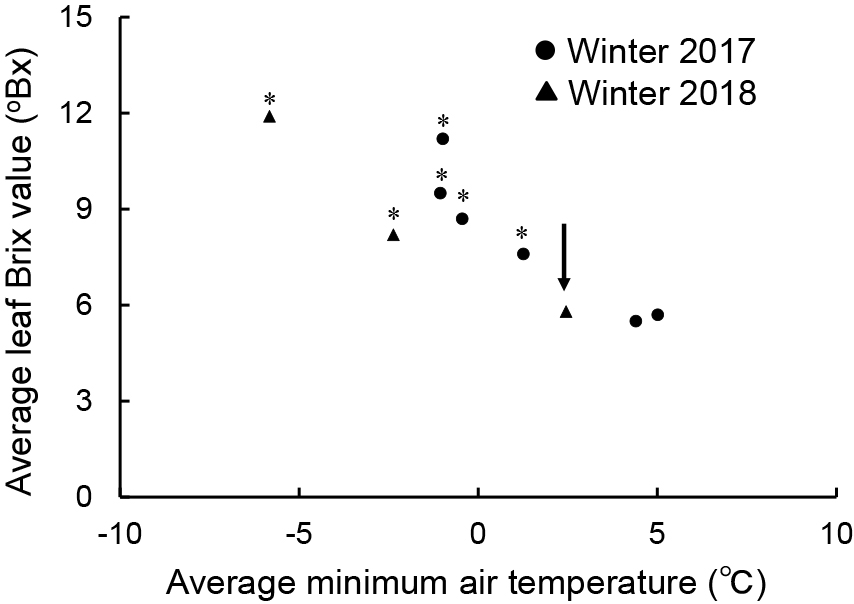
Relationship between leaf Brix value and average minimum air temperature for 10 days before harvest day. * represents the significant difference of the value indicated by the arrowed point at P < 0.05 by Dunnett’s test (n = 9). At the allowed point, the average minimum air temperature and leaf Brix value were 2.5°C and 5.8 °Bx, respectively. The average leaf Brix value (vertical axis) in each plot includes data from 6 individual plants.
In our study, borecole survived in the greenhouse without heating, even in midwinter. The Brix value increased over time (Figs. 5A, C, D, and 6A, C) and behaved consistently with that reported by Jurkow et al. (2019). Jurkow et al. (2019) reported the necessity of exposure to air temperatures of −5°C or lower to increase the sugar concentration and dry matter ratio of borecole plants. The greenhouse in this study was equipped with better heat insulation than that used by Jurkow et al. (2019). During this study period, for only two days did the daily minimum greenhouse air temperature fall below −5°C (Fig. 3). Examining the Brix value and dry matter ratio identified increases even when the minimum temperature was higher than −5°C. Tamura (2005) demonstrated, in spinach and komatsuna, the close relationship between sugar concentration and average minimum air temperature over the 10 days before harvest. These findings indicated that the sugar concentration increased when the average daily minimum air temperature, over 10 days before harvest, was 5°C or less. In the current study, the relationship between average daily minimum air temperature over the 10 days before harvest and the borecole leaf Brix value was evaluated (Fig. 8). From this examination, the borecole leaf Brix value increased significantly when the daily minimum air temperature over 10 days before harvest was less than 1.3°C.
The daily minimum air temperature in the greenhouse fell below 0°C almost every day after November 30, 2017, November 20, 2018, and December 3, 2019 (Fig. 3A, C, D). On days when the air temperature in the greenhouse dropped to less than 0°C, freezing of plants was observed in the early morning (Fig. 4A). Freezing of the cytoplasm during frost will normally kill the cell. However, some plants with freeze-tolerance traits can survive as the ice formation occurs outside the cell. This extracellular freezing then draws water from within the cells, leading to their dehydration (Ashworth and Pearce, 2002; Mazur, 1969). In our study, as the daily maximum air temperature was higher than 0°C on most days (Fig. 3A, C, D), the extracellular ice in leaves melted during the daytime. Therefore, the inference is that this dehydration caused by extracellular freezing occurs repeatedly. In addition, Okada et al. (2005) demonstrated in a spinach experiment that when the soil temperature was 10°C or less, the dry matter ratio and sugar concentration of plants increased, suggesting that plant dehydration may result from the low soil temperature, which decreases root activity. In the 2019 winter experiment, daily average soil temperatures after November 25 were less than 10°C (Fig. 3D), with a corresponding sharp increase in dry matter ratio. This mirrors the findings of Okada et al. (2005) for spinach. In our study, the soil temperature range that weakens borecole root activity towards water uptake could not be specified. However, the root activity was still presumed to decrease in response to low soil temperature.
Moreover, the leaf Brix value increased from November 14, 2017, with no difference in the number of fresh leaves (Fig. 5A). In 2019, the Brix value was not measured. However, there was no difference in behavior in either fresh leaf weight or dry matter ratio increase (Fig. 5D). The development of new leaves is well-known to be slowed by low temperature (Cao and Moss, 1989; De Koning, 1994), which suggests preferential accumulation of photosynthetic products in the existing tissues. This accumulation contributed to increases in leaf Brix value and dry matter ratio. To elucidate the detailed mechanisms of the phenomenon identified in this study, more precise experiments using a phytotron will be necessary.
This study specifically examined the leaf Brix values as a function of leaf order. The leaf Brix values of borecole in lower leaf orders, from the top of the plant, were greater than those from higher leaf orders, closer to the bottom of the plant, irrespective of harvest day. The Brix values of all leaves increased over winter, irrespective of leaf order (Fig. 6). Tamura (2005), after investigating spinach and komatsuna sugar concentrations by leaf order, reported that the leaf sugar concentrations of higher order leaves, from the lower section of the plant, were less than those of the lower order leaves from the top of the plant. A similar relationship between Brix value and leaf order was observed in our borecole experiment, supporting the results reported by Tamura (2005). Therefore, borecole with optimal Brix values can be obtained by exposure to low temperatures and selecting the lower leaf order leaves that are closer to the top of the plant.
Nitrate concentrations decreased considerably during the winters of 2018 and 2019 (Fig. 7). Nitrate ions are absorbed by the roots into the plant and are reduced to nitrite and ammonia by enzymatic action, followed by conversion to various amino acids with excess nitrate ions accumulating in the vacuole (Kiba et al., 2021). As earlier described, in the winter cultivations of this study we assumed a decreased root activity due to the low soil temperature. Consequently, it was speculated that the accumulated nitrate ions were converted into various amino acids resulting in a gradual decrease in leaf nitrate concentration. Given the correlation between nitrate concentration and leafy vegetable bitterness intensity (Noda and Makuta, 2015) with the poor popularity of bitter vegetables in Japan, these findings suggest that the low temperature cultivated borecole is less bitter, thus the cold exposure can improve borecole taste and market quality.
In conclusion, leaves with decreased bitterness and improved sweetness are produced under low temperatures in borecole leaves from the upper sections of the plant. The borecole, with a large leaf Brix value and low nitrate concentration due to low temperature exposure, is a strong candidate as an attractive, value-added product in Japan. Further study is necessary to optimize borecole cultivation methods for enhanced functionality.
We are indebted to M. Ishizaki, M. Sasaki, S. Nakagawa, H. Sato, M. Fujii, S. Nishizuka, S. Toshima, H. Ohta and E. Nishiyama of HRO for their plant and field management and generous assistance. We are also deeply grateful to Dr. M. Takahama of HRO for his willingness to review this report.