Abstract
Genetic improvement of flower vase life is an important breeding target for ornamental plants. As the vase life of cut dahlia (Dahlia variabilis) flowers is very short, we initiated a conventional crossbreeding research program in 2014 to improve it. We evaluated the vase life of dahlia seedlings during summer (from July to early September) to develop dahlias that grow and bloom under high temperatures and have excellent vase life for Japanese summer conditions. Crossing and selection over five generations greatly improved vase life. The mean vase life increased from 4.4 days in the 1st generation, derived from 22 parental cultivars, to 8.0 days in the 5th generation, a net increase of 3.6 days. Mean vase life increased significantly by 1.6 days from the 4th to the 5th generation, indicating continued improvement. The mean vase life of the 12 lines selected from the 3rd and 4th generations ranged between 6.0 and 15.9 days in distilled water and from 7.8 to 14.6 days in GLA solution (10 g·L−1 glucose, 0.5 mL·L−1 CMIT/MIT [isothiazolinone derivatives], and 50 mg·L−1 aluminum sulfate). Vase life was further extended by 0.5 to 4.0 days using 6-benzylaminopurine (BA) sprays compared to GLA alone. In particular, the 4th-generation line 003-15 vase life was 13.8 to 15.9 days (2.5× that of ‘Kamakura’, a leading white dahlia cultivar in Japan) in distilled water, 12.0 to 14.6 days (1.8× to 2.2×) in GLA, and 13.9 to 15.3 days (1.7× to 1.9×) in GLA+BA in winter and spring. Cut flowers of line 003-15 harvested under high temperatures in July–August and at 28°C with GLA treatment also had long vase life. The pedigree of line 003-15 suggests that genes related to long vase life derived from ‘Micchan’ (with a long vase life) may have accumulated or duplicated in line 003-15. Finally, four selected lines, including 003-15, had high ethylene sensitivity, as 10 μL·L−1 ethylene treatment caused petal abscission in 2.0 to 2.8 days.
Introduction
Long flower vase life is an essential trait in ornamental plants, influencing quality and consumer appeal (Onozaki, 2018a). Long vase life is a key factor for consumer repurchase of cut flowers (Giovannini et al., 2021). Unlike ornamental traits such as flower color, size, and type, flower vase life is a difficult genetic character to assess accurately, being a complex quantitative trait involving multiple genes with additive effects (Boxriker et al., 2017, 2018), and often-unclear modes of inheritance. Also, the measurement of flower vase life across many individuals is exceptionally labor-intensive.
Dahlia (Dahlia variabilis), in the Asteraceae family, is an important crop sold as cut flowers, garden ornamentals, and potted plants in many countries, with increasing popularity in Japan (Onozaki and Azuma, 2019; Yang et al., 2021). Dahlias are not surveyed in the nationwide Statistics of Agriculture, Forestry and Fisheries in Japan, but the total transactional amount of cut dahlia flowers handled in the Tokyo metropolitan market increased from 391 million JPY in 2009 to 697 million JPY in 2019 (a 78% increase over 10 years; http://www.shijou-tokei.metro.tokyo.jp/). Demand for cut dahlia flowers is increasing year by year, and dahlia has the highest transaction value rate increase among the 30 major flower items in the Tokyo metropolitan market.
A major weakness of dahlia is its short vase life, which limits widespread use and general household consumption. Dahlia cultivars’ vase life is usually only 3–7 days at room temperature (23°C) without preservative treatment (Onozaki and Azuma, 2019). Dahlias were previously used mainly for business purposes, such as weddings and events. However, recently numbers of products handled at retail stores and household demand are increased. To encourage the purchase of cut flowers for home ornamental use in England, volume sellers created a sales system guaranteeing the vase life of cut flowers leading to remarkable sales increases. Since implementation, this practice spread to France, Germany, the Netherlands, and Japan (Shimizu-Yumoto, 2018). However, for dahlia, it is difficult to guarantee a 1-week vase life at room temperature (Ichimura et al., 2011). To address the limited consumer appeal and marketability of short-lived cut flowers such as dahlias, cultivars are being selected with attention to vase life. Excellent vase life is essential to expand and meet demands with a large distribution volume, thus it is important to develop cultivars with long vase life in dahlia breeding.
The Institute of Vegetable and Floriculture Science (NIVFS), NARO, Japan, initiated a breeding research program in 2014 to improve dahlia vase life (Onozaki and Azuma, 2019). It aims producing commercially successful dahlia cultivars with a long vase life and clarify the underlying mechanism. Three cultivars—‘Eternity Torch’, ‘Eternity Romance’, and ‘Eternity Rouge’ (called the “Eternity series”)—with a genetically determined long vase life were developed in 2020 (Onozaki and Azuma, 2022). The flower vase life of these cultivars was 6.3 to 12.0 days (1.4× to 2.2× that of a standard cultivar, ‘Kamakura’) in distilled water and 9.0 to 13.4 days (1.1× to 2.0×) in “GLA” (10 g·L−1 glucose, 0.5 mL·L−1 CMIT/MIT [isothiazolinone derivatives], and 50 mg·L−1 aluminum sulfate) at 23°C, a 12-h photoperiod, and 70% relative humidity (RH). The trial cultivation test results of Eternity series in four prefectural research institutes (Akita Agricultural Experiment Station, Nara Agricultural Research Center, Kochi Agricultural Research Center, and Miyazaki Agricultural Research Institute) in Japan from 2018 to 2019 demonstrated that cultivation is possible in dahlia production areas nationwide (Onozaki and Azuma, 2022). Four experienced dahlia growers in Japan conducted field demonstration tests from 2020 to 2022 and encountered no major cultivation problem.
Commercial cut flower production of the Eternity series, based on plug cuttings sold by two seed and nursery companies, began in 2022 all over Japan. These cultivars are expected to further expand dahlia cut flowers for home use and export. With the impact of covid-19, the primary demand for cut flowers is shifting from business to home use. Therefore, dahlia growers and retailers are requesting new cultivars with additional colors and longer vase life.
Previously, we reported increased dahlia vase life in many dahlia lines by repeated crossing and progeny selection for long vase life over three generations, from 2014 to 2018 (Onozaki and Azuma, 2019). Here, we report the results of further crossing and selection over two more generations, using our previously selected lines as parental material. We additionally extended dahlia flower vase life and identified the new line 003-15, with ultra-long vase life.
We previously reported that exogenous ethylene exposure accelerated petal abscission or wilting in most dahlia cultivars tested (Azuma et al., 2020). Particularly, petals of ‘Carnelian’, ‘Garnet’, ‘Namahage Cute’, ‘Namahage Magic’, and ‘Saisetsu’ abscised within 2 days in 10 μL·L−1 ethylene. Yang et al. (2021) reported that exogenous ethylene treatment of ‘Saisetsu’ cut flowers accelerated the decline of petal drawing resistance, shortening vase life.
As dahlia cultivars with high ethylene sensitivity suffer from petal abscission during transport, especially in summer, it is necessary to select ethylene-resistant cultivars (Yamagata, 2022). Thus, we also investigated ethylene sensitivity in our lines with long vase life.
Materials and Methods
Crossing and selection
We chose 22 commercial dahlia cultivars with large differences in vase life as initial breeding materials (Onozaki and Azuma, 2019). In autumn 2014, crosses were made among these cultivars. Lines exhibiting a long vase life were then selected as parents of the next generation for five generations, from 2015 to 2021 (Onozaki and Azuma, 2019).
Crosses among selected lines were made in October of each year, mainly in the open field and partly in a vinyl house, using the standard dahlia crossing method (Konishi, 2009; Onozaki, 2018b) as previously described (Onozaki and Azuma, 2019).
We obtained 261 seeds from 4 crosses (Cross nos. 914–917) made in October 2018 among 7 selected 2nd-generation lines. On 29 March 2019, 168 seeds were sown, and 137 plants germinated by 24 May 2019; 123 of these plants were grown in the open field at NIVFS. Plants that did not flower until 6 September 2019 (the last day of evaluation) were discarded, and the remaining 74 plants were considered the 3rd generation, in addition to the 155 seedlings in the previous report (Onozaki and Azuma, 2019) (Table 1).

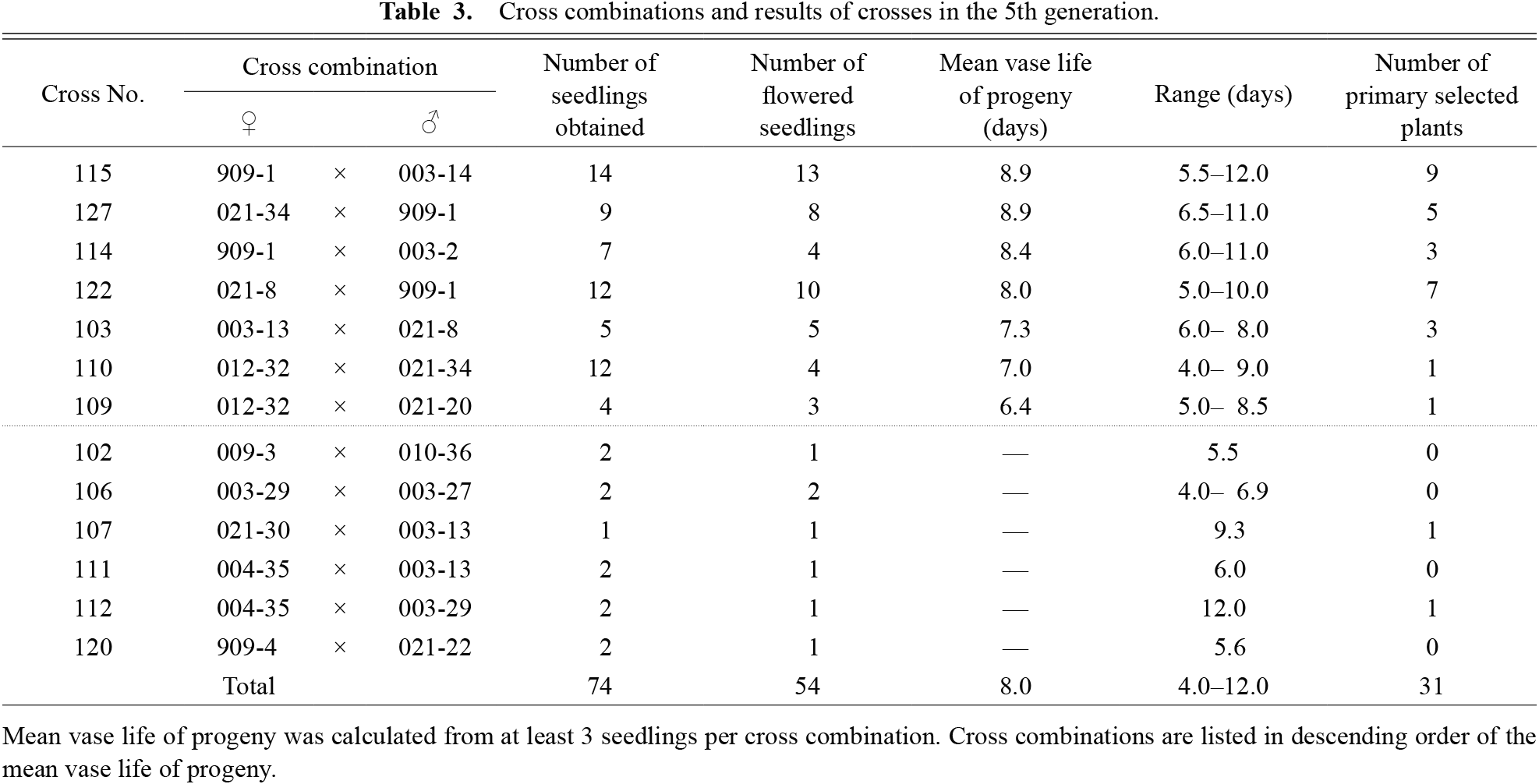
In October 2017, 2018, and 2019, a total of 80 plants with the longest mean vase life (> 6.1, > 6.2, and > 6.0 days, respectively) were selected and used for the next crosses. Seeds obtained from the crosses were sown on 29 March 2019, 30 March 2020, or 30 March 2021, and 284 plants were grown in the open field. Progeny that did not flower until 6 September 2019, 11 September 2020, or 10 September 2021 (the last day of evaluation) were discarded, and the remaining 233 plants were considered the 4th generation (Table 2). In October 2019, 2020, and 2021, a total of 58 plants with the longest mean vase life (> 6.5, > 6.4, and > 7.5 days, respectively) were selected and used for the next crosses. The seeds obtained from the crosses were sown on 30 March 2021, and 74 plants were grown in the open field. Progeny that did not flower until 10 September 2021 (the last day of evaluation) were discarded, and the remaining 54 plants were considered the 5th generation (Table 3). In October 2021, 31 plants with the longest mean vase life (> 7.5 days) were selected.

The initially selected 3rd- and 4th-generation lines were propagated vegetatively, and 10 out of 80 lines and 15 out of 58 lines were further selected. We examined the vase life of the cut flowers of the first 10 selected lines in detail in two seasons and three cultivation styles (winter–spring in greenhouse heated above 10°C, daylength extension to 14.5 h using artificial lighting from 05:00 to 07:00 and from 16:00 to 19:30) and summer–autumn in the open field or in the vinyl house (no heating, natural daylength). The latter 15 lines with a vase life of ≥ 10 days in GLA solution in any situation were selected. The latter selection was based primarily on vase life but also partly on other important traits (e.g., flowers are fully double with no open center at any time, productivity, earliness of flowering).
Vase life evaluation in seedlings and selected lines
As an antibacterial agent, we used CMIT/MIT (Kathon CG, formerly Legend MK; Rohm and Haas Japan K.K., Tokyo, Japan) solution containing 11.3 g·L−1 5-chloro-2-methyl-4-isothiazolin-3-one (CMIT) and 3.9 g·L−1 2-methyl-4-isothiazolin-3-one (MIT). Flowers were harvested when the first row of petals from the outside opened at right angles to the stem. The stem was cut to 40 cm, and all leaves except the top ones were removed. One or two cut flowers were placed in 300- or 500-mL conical beakers containing distilled water (DW), 0.5 mL·L−1 CMIT/MIT, or GLA solution (10 g·L−1 Glucose, 0.5 mL·L−1 CMIT/MIT [Legend MK], 50 mg·L−1 Aluminum sulfate; Ichimura et al., 2006).
A spray treatment with 6-benzylaminopurine (BA) extends the vase life of cut dahlia ‘Kokucho’ (Shimizu-Yumoto and Ichimura, 2013). Combining BA spray with immersion in sugar solution is very effective at extending vase life (Shimizu-Yumoto, 2018). The use of commercial BA products is now common for cut dahlia flowers in Japan (Shimizu-Yumoto et al., 2020). Therefore, we also investigated combined BA spraying and GLA solution, using a commercial preservative (Miracle Mist FF; Chrysal Japan, Osaka, Japan) as the BA agent, sprayed on day 0 (harvest day).
Flowers were maintained at 23°C or 28°C (with GLA), 70% RH, under a 12-h photoperiod provided by cool fluorescent lamps (10 μmol·m−2·s−1 irradiance) and were evaluated daily. The solution was not exchanged, but if its level decreased to less than half of the initial level, more was added.
The vase life of dahlia seedlings for each generation was evaluated from July to early September in 0.5 mL·L−1 CMIT/MIT solution. The petal thickness of each harvested flower was measured with a Digimatic thickness gauge (547-301; Mitutoyo Corp., Kawasaki, Kanagawa, Japan). The outermost three petals were measured on day 0 and the average value used as the data of the seedlings.
Flower senescence was classified by eye into wilting, wilting with browning, browning, or petal abscission (Onozaki and Azuma, 2019). Flower vase life was determined as the number of days from harvest until one-third of all of petals showed wilting with or without browning or until petal abscission of more than 10 petals.
Plant growth conditions in two seasons for vase life evaluation
The vase life of six final 3rd-generation lines (823-3, 816-7, 814-13, 828-18, 815-20, and 916-34) and four control cultivars (‘Kamakura’, ‘Port Light Pair Beauty’, ‘Kokucho’, and ‘Micchan’) was evaluated in 2019, 2020, and 2021 (Table 4). That of six final 4th-generation lines (909-1, 909-4, 003-15, 012-19, 003-29, and 012-32), the four above control cultivars, and the three Eternity series cultivars was evaluated in 2020, 2021, and 2022 (Table 5). The mother plants of each cultivar or line for propagating cuttings were grown in 21-cm-diameter pots in a greenhouse with artificial lighting (daylength extension to 14.5 h).

In winter–spring pot cultivation, rooted cuttings were planted in 21-cm-diameter pots with 4 L of standard culture soil (Royal Culture Soil; Tachikawa Heiwa Nouen Co., Ltd., Tochigi, Japan) on 17 October 2019 or 23 October 2020. The shoots from each plant were pinched to 2 or 3 nodes around 3 weeks after planting. Plants were grown until May in a greenhouse at NIVFS heated above 10°C, with artificial lighting (daylength extension to 14.5 h) under red light-emitting diode (LED) lamps (DPDL-R-9W; Nabesei Co., Ltd., Aichi, Japan). Flower vase life was evaluated from January to May 2019 or from December 2020 to May 2021.
In soil cultivation over winter–spring, rooted cuttings were planted 40 cm apart in 60-cm-wide ridges in a greenhouse on 30 July 2020 or 2 August 2021. Slow-release fertilizer (Ecolong 413-180-day type, 170 g·m−2, N:P2O5:K2O = 14:11:13; JCAM Agri. Co., Ltd., Tokyo, Japan) and BM fused phosphate (40 g·m−2; N:P2O5:K2O = 0:20:0; Asahi Industries Co., Ltd., Tokyo, Japan) were applied as base fertilizer. The shoots from each plant were pinched to 2 or 3 nodes on 20 August 2020 or 23 August 2021 and again to 1 or 2 nodes on 16 September 2020 or 22 September 2021. Plants were grown until May in a greenhouse at NIVFS and heated above 10°C, with artificial lighting (daylength extension to 14.5 h), as described above. Flower vase life was evaluated from November 2020 to May 2021 or from December 2021 to May 2022.
Summer experiments were carried out to evaluate the vase life of flowers grown under high temperature. For pot cultivation in the vinyl house under a shade net (cool white, 45%–50% shade; Dio Chemicals, Ltd., Tokyo, Japan) under 50%–55% sunlight (natural daylength, no heating), rooted cuttings of each cultivar or line were planted in 24-cm-diameter pots with 5 L of standard culture soil on 21 April 2020 or 20 April 2021. The shoots from each plant were pinched to 2 or 3 nodes on 17 June in each year. The plants were grown during summer under high temperatures, and cut flowers were placed in 0.5 mL·L−1 CMIT/MIT solution. Flower vase life was evaluated in July and August each year. From 1 July to 31 August in each year, the minimum, mean, and maximum average temperatures in the vinyl house were 22.4–22.5, 27.1–27.3, and 34.1–35.4°C, respectively.
Evaluation of ethylene sensitivity
The three cultivars and four selected lines were grown in 21-cm-diameter pots as above for winter–spring culture except for artificial lighting (K-RD110V75W/D; Panasonic Co., Ltd., Osaka, Japan). Flowers were harvested from January to April 2022 and their ethylene sensitivity was evaluated. Cut flowers with a stem length of 20 cm and no leaves held in 500-mL conical beakers containing 0.5 mL·L−1 CMIT/MIT solution were placed in 70-L transparent acrylic boxes fitted with a septum through which ethylene was introduced at 0 (control) or 10 μL·L−1, as described previously (Azuma et al., 2020). Throughout the experiment, the boxes were kept at 23°C with a 12-h photoperiod provided by cool fluorescent lamps (10 μmol·m−2·s−1 irradiance). After 24 h, each box was opened for 5 min to release the air, then closed again, with ethylene reintroduced at the same concentration. This procedure was repeated daily until petal wilting or abscission was observed. Ethylene sensitivity was evaluated daily and was recorded as the elapsed time from the start of ethylene treatment to the observation of either petal wilting in the florets of three whorls or abscission of > 10 petals, as confirmed by gently touching the petals by hand. Cut flowers of each line in 0.5 mL·L−1 CMIT/MIT solution were held outside the boxes at 23°C and 70% RH under a 12-h photoperiod, with 10 μmol·m−2·s−1 irradiance during the light period, until petal wilting or abscission was observed.
Statistical analyses
The results in Tables 4–6 were analyzed with BellCurve for Excel software (Social Survey Research Information Co., Ltd., Tokyo, Japan) using Tukey’s test (P < 0.05).
Results
Crossing and selection
The frequency distributions of vase life in the 1st to the 5th generations were continuous and normal (Fig. 1). The frequencies of flowers with superior vase life (≥ 7 days) were 3.8% in the 1st generation but rose to 70.4% in the 5th generation. Variation was small in the 1st generation (S.D. = 1.15) but was larger in the 5th (S.D. = 2.03). The mean vase life of the 1st generation derived from crossing 22 cultivars was 4.4 days. This was improved to 8.0 days, a net increase of 3.6 days (Fig. 1). The biggest improvement occurred between the 4th and 5th generations with mean vase life significantly increased by 1.6 days (t-test, P < 0.01).
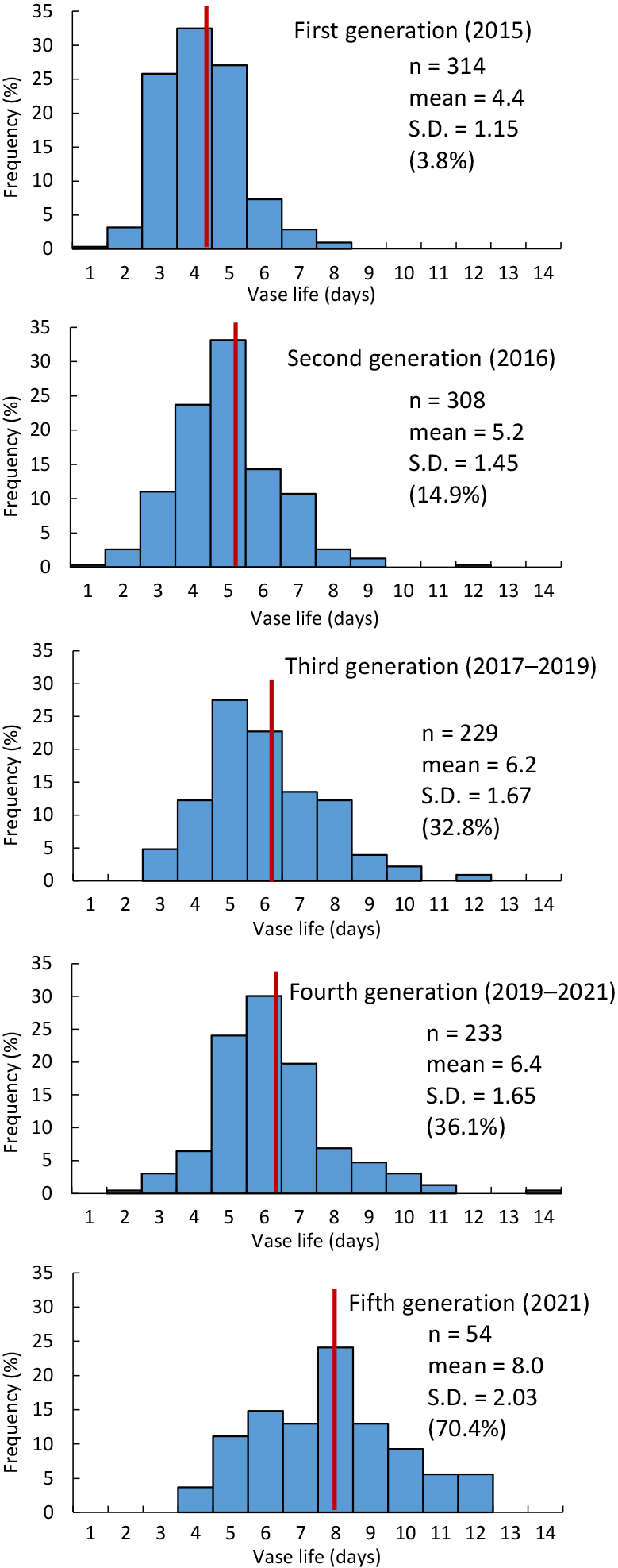
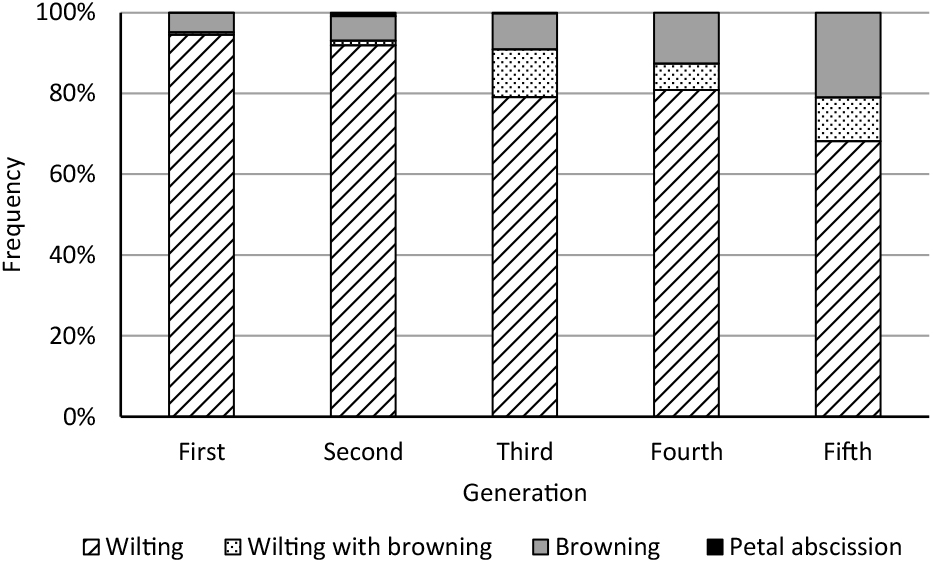
The proportion of flowers with wilting decreased from 94.5% in the 1st generation to 68.2% in the 5th (Fig. 2). Conversely, the proportion of flowers with browning (browning + wilting with browning) increased from 5.4% in the 1st generation to 31.8% in the 5th. Petal abscission decreased from 0.1% in the first generation and 0.9% in the 2nd to nil in the 3rd, 4th, and 5th generations (Fig. 2).
Vase life relationship with earliness and petal thickness
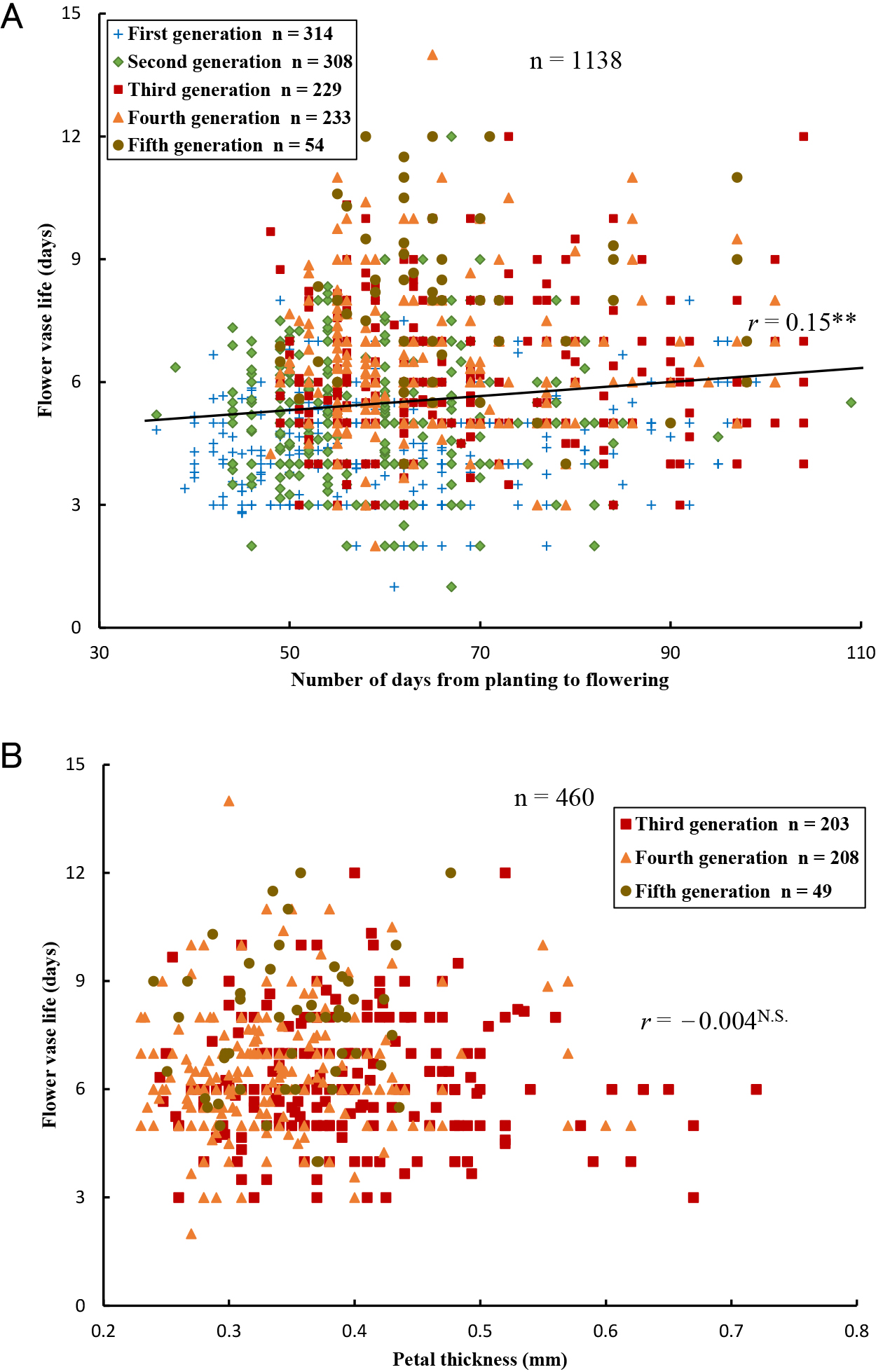
Earliness (the number of days from planting to flowering) varied among 1138 seedlings from 36 to 109 days. It had a weak but significant positive correlation with vase life (Fig. 3A). Petal thickness varied among 460 seedlings from 0.23 to 0.72 mm. It had no significant correlation with vase life (Fig. 3B).
Mean vase life of progeny per cross combination
In the 3rd generation, the difference between the cross combination with the longest mean vase life (cross no. 823: 609-4 × 631-23; 8.1 days) and that with the shortest (no. 821: 628-32 × 615-55; 5.1 days) was 3.0 days (Table 1). In the 4th generation, the difference between those with the longest mean vase life (nos. 130: 828-46 × 823-3; 905: 805-47 × 803-4; 7.1 days) and that with the shortest (no. 013: 828-18 × 815-20; 4.5 days) was 2.6 days (Table 2). In the 5th generation, the difference between those with the longest mean vase life (nos. 115: 909-1 × 003-14; 127: 021-34 × 909-1; 8.9 days) and that with the shortest (no. 109: 012-32 × 021-20; 6.4 days) was 2.5 days (Table 3).
Vase life of cultivars and selected lines
We found a wide range of variation in cut flower vase life of the four control cultivars and six selected 3rd-generation lines (Fig. 4A) grown in two seasons and three cultivation styles (Table 4). In winter and spring, the mean vase life of ‘Kamakura’ was 5.2 to 5.9 days in DW, 6.3 to 8.0 days in GLA, and 8.3 to 9.2 days in GLA+BA; that of the six selected lines was 6.0 to 11.7 days in DW (1.2×–2.0× that of ‘Kamakura’), 7.8 to 12.2 days in GLA (1.2×–1.9×), and 9.7 to 14.7 days in GLA+BA (1.2×–1.7×) (Table 4). In summer in the vinyl house with 0.5 mL·L−1 CMIT/MIT, the mean vase life of three control cultivars (‘Kamakura’, ‘Port Light Pair Beauty’, and ‘Kokucho’) was 3.5 to 4.0 days and that of ‘Micchan’, a long-vase-life cultivar (Onozaki and Azuma, 2019), was 6.8 days; that of the four selected lines was 7.4 to 10.9 days (1.9×–2.7× that of ‘Kamakura’). Under high temperature (28°C) with GLA solution, the mean vase life of ‘Kamakura’ was 5.6 days while that of the six selected lines was 6.5 to 9.8 days (1.2×–1.8×) (Table 4).

We also found a wide range of variation in the vase life of the cut flowers of the four control cultivars, three Eternity series cultivars, and six selected 4th-generation lines (Fig. 4B) in two seasons and three cultivation styles (Table 5). In winter and spring, the mean vase life of ‘Kamakura’ was 5.6 to 6.3 days in DW, 6.7 to 6.8 days in GLA, and 7.4 to 9.2 days in GLA+BA; that in the six final lines was 8.3 to 15.9 days in DW (1.5×–2.5× that of ‘Kamakura’), 9.4 to 14.6 days in GLA (1.4×–1.9×), and 12.0 to 15.3 days in GLA+BA (1.4×–1.9×) (Table 5). In summer in the vinyl house with 0.5 mL·L−1 CMIT/MIT, the mean vase life of the control cultivars ‘Kamakura’, ‘Kokucho’ and ‘Micchan’ was 4.0, 5.5, and 5.5 days, respectively; that of the five selected lines was 7.6 to 12.0 days (1.9×–3.0× that of ‘Kamakura’). Under high temperature (28°C) with GLA solution, the mean vase life of ‘Kamakura’ was 6.1 days and that in the five selected lines was 7.8 to 11.2 days (1.3×–1.8×) (Table 5).
In winter and spring, the mean vase life of 003-15 was 13.8 to 15.9 days (2.5× times that of ‘Kamakura’) in DW, 12.0 to 14.6 days (1.8×–2.2×) in GLA, and 13.9 to 15.3 days (1.7×–1.9×) in GLA+BA, and was significantly longer than that of control cultivars ‘Kamakura’ and ‘Port Light Pair Beauty’ (Table 5). In DW during senescence, the outer petals of the control cultivar ‘Port Light Pair Beauty’, with short vase life (Onozaki and Azuma, 2019), began to wilt rapidly after 4 days (Fig. 5B) and completely lost ornamental value after 8 days (Fig. 5C); ‘Micchan’ showed wilting after 8 days (Fig. 5C); whereas line 003-15, with ultra-long vase life, did not show visible signs of senescence until 12 days (Fig. 5D).
Ethelene sensitivity of cultivars and final selected lines
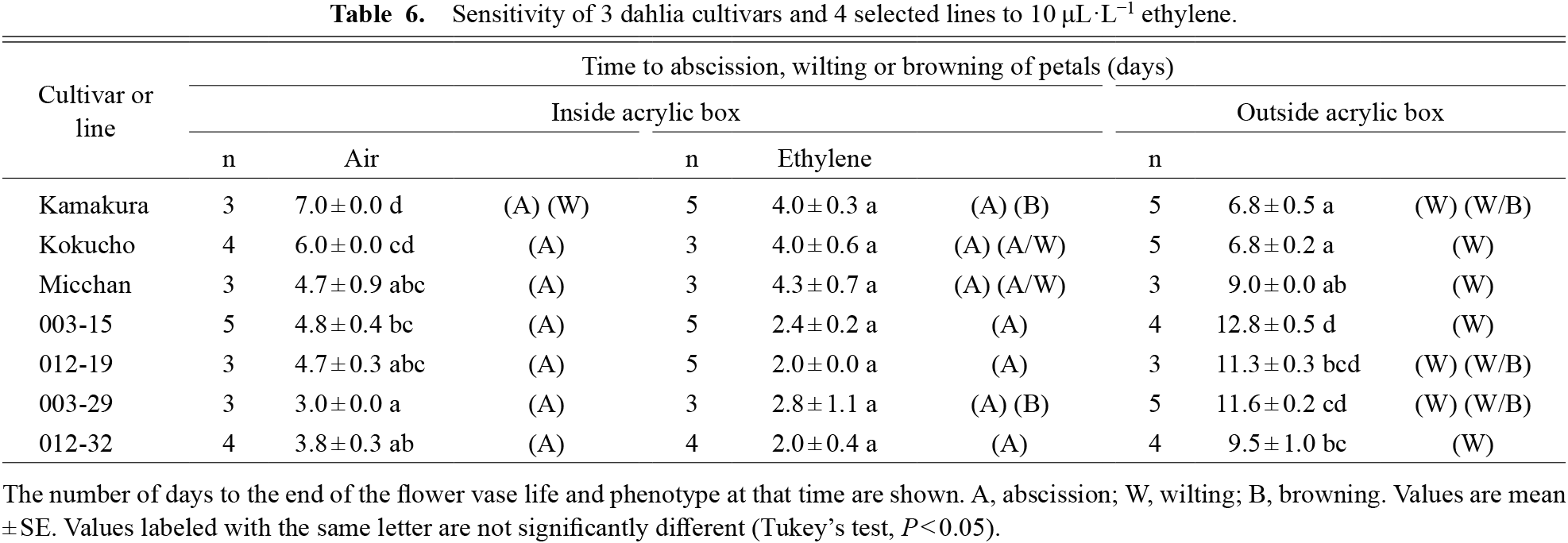
Treatment with 10 μL·L−1 ethylene shortened the time to petal wilting or abscission in all cultivars or lines examined relative to that in air (Table 6). Outside the acrylic box, all cultivars and lines showed petal wilting or browning but no abscission. Particularly, lines 003-15 and 003-29 had significantly longer times to petal wilting or browning than the three control cultivars. However, petal abscission was observed in the ethylene treatment, as well as inside the box without ethylene. Moreover, four final selected lines with long vase life had higher ethylene sensitivity than the three cultivars, although the difference in reaction was not significant.
Discussion
Cut dahlia flowers are now popular in Japan owing to their wide variation in color, shape, and size, but their marketability is limited by their short vase life (Yang et al., 2021). To expand dahlia consumption, we developed lines with a genetically determined long vase life. The mean vase life of the 1st generation, derived from crossing 22 cultivars in 2014, is 4.4 days (Fig. 1). In contrast, that of the 5th generation is 8.0 days (Fig. 1).
We previously genetically improved carnation vase life over nearly two decades (Onozaki, 2018a), crossing and selecting promising offspring for seven generations from 1992 to 2008 (Onozaki et al., 2001, 2006a, 2011). This increasing mean vase life from 7.4 days in the 1st generation to 15.9 days in the 7th generation using conventional crossing techniques. We developed three cultivars—‘Miracle Rouge’, ‘Miracle Symphony’ (Onozaki et al., 2006b), and ‘Kane Ainou 1-go’ (Hotta et al., 2016)—with a genetically determined long vase life, and two lines—532-6 (Onozaki et al., 2011) and 806-46b (Onozaki et al., 2015)—with an ultra-long vase life (> 27 days) under standard conditions.
Four cycles of selection and crossing led to a 3.6-day increase in the mean vase life of dahlia seedling populations from the 1st to the 5th generations (Fig. 1). A selection effect was observed in all generations. This clearly demonstrates the efficacy of crossbreeding for genetic improvements in vase life, not only in carnations but also in dahlias.
In addition to research on carnations and dahlias by our group, the inheritance, response to selection, or QTL analysis of vase life are reported for gerbera (Wernett et al., 1996), Asiatic hybrid lily (Van der Meulen-Muisers et al., 1999), rose (Carvalho et al., 2015; Fanourakis et al., 2012), and chrysanthemum (Van Geest et al., 2017).
We evaluated the relationships between flower vase life with earliness (number of days from planting to flowering) and petal thickness (Fig. 3). Previously, in carnations, we found a significant positive correlation (r = 0.49*) between vase life and petal thickness (Onozaki et al., 2011). A famous Japanese dahlia breeder (Yusaku Konishi, September 2014) and a main retailer in the flower market (Jun Shishido, October 2017) both suggested that long vase life in dahlia flowers may be related to petal thickness and tightness. Generally, thick and hard petals appear advantageous for cut flower vase life by reducing damage during transport, but we observed no correlation. Ichimura et al. (2002) also reported no significant correlation between vase life and petal thickness for cut rose flowers (r = 0.26). On the other hand, we did find relationships with senescence pattern, when the flowers lost ornamental value (Fig. 2): the wilting senescence pattern decreased gradually with generation, and the rate of browning and wilting with browning increased from 5.4% to 31.8%.
In carnation, the senescence pattern for normal cultivars like ‘White Sim’ and ‘Nora’ are petal in-rolling with rapid whole-petal wilting (Onozaki et al., 2006b), well-known characteristics of ethylene-dependent senescence in carnation flowers. On the other hand, ‘Miracle Rouge’ and ‘Miracle Symphony’ with long vase life did not show petal in-rolling and rapid wilting at senescence, but faded and turned brown from the petal edges (Onozaki, 2018a; Onozaki et al., 2006b). It is possible that the short vase life of normal dahlia flowers is caused by programed cell death (PCD) (Azuma et al., 2019). Further studies are required to clarify the mechanisms among long vase life, senescence pattern, and PCD in dahlias.
Wild dahlia species are native to the highlands of Mexico and Central America (Behr and Debener, 2004; Okumura and Fujino, 1989). Although these regions are at low latitudes, the climate is relatively cool owing to the high altitudes, where dahlias grow at elevations between 1,500 and 3,700 m (Saar et al., 2003). Under high temperatures, such as summer in Japan, dahlia does not grow well (Miyoshi, 2018; Yamagata, 2022). As the vase life of ‘Kamakura’, ‘Kokucho’, and ‘Shukuhai’ in DW decreased from July to October and increased from December to February (Naka et al., 2015), we evaluated the vase life of dahlia seedlings during summer (from July to early September) to develop dahlias that grow and bloom under high temperatures and have excellent vase life under Japanese summer conditions. The vase life of our breeding lines 823-3, 916-34, 003-15, and 003-29 grown in a vinyl house in summer with 0.5 mL·L−1 CMIT/MIT solution was ≥ 10 days (2.5×–3.0× that of ‘Kamakura’) (Tables 4 and 5). The results clearly show that our breeding procedure is effective for dahlias in breeding a genetically determined long vase life, not only for winter–spring, but also in summer, when vase life generally decreases.
The 4th-generation selected line 003-15 achieved the longest vase life of all, with means of 13.8 and 15.9 days in DW (Table 5), which we define as “ultra-long vase life”. Line 003-15 showed no rapid wilting, which is the typical senescence pattern of dahlia, and instead slowly desiccated and wrinkled.
Although line 003-15 had ultra-long vase life in DW, the vase lives were shorter in GLA and GLA+BA in 2021–2022 winter and spring (Table 5). In GLA solution, stem bases of 003-15 gradually discolored and the solution became turbid at > 10 days, despite antibacterial properties. Mold often grew on the stem surface and on the GLA solution surface. This may explain the shorter vase life of 003-15 in GLA solution than in DW.
BA spray extends the vase life of 27 dahlia cultivars by 0.6 to 2.8 days relative to controls (Tsujimoto et al., 2016). Combined treatment of BA spraying and sugar solutions is very effective at extending dahlia vase life (Shimizu-Yumoto, 2018). We found that the combination of BA spray + GLA solution extended the vase life of 12 lines by 0.5 to 4.0 days relative to GLA (Tables 4 and 5). These results clearly indicate that BA treatment is effective not only for normal cultivars, but also for our breeding lines with long vase life.
The four selected lines were sensitive to exogenous application of 10 μL·L−1 ethylene, which caused petal abscission in 2.0 to 2.8 days (Table 6). We observed petal abscission in all seven cultivars and lines, and under high RH (inside the acrylic box) without ethylene. Azuma et al. (2020) reported that the RH in the same acrylic boxes was 100%, much higher than the 70% RH outside. Particularly, in the four selected lines the reaction time under high RH (inside the box) without ethylene was 3.0 to 4.8 days (Table 6), less than half that under 70% RH (outside the box). Thus, ethylene treatment and high RH both lead to petal abscission, shortening vase life in our four selected lines. In addition, it is possible that endogenous ethylene from cut flowers accumulated inside the closed box and induced petal abscission. It is necessary to clarify this in future work.
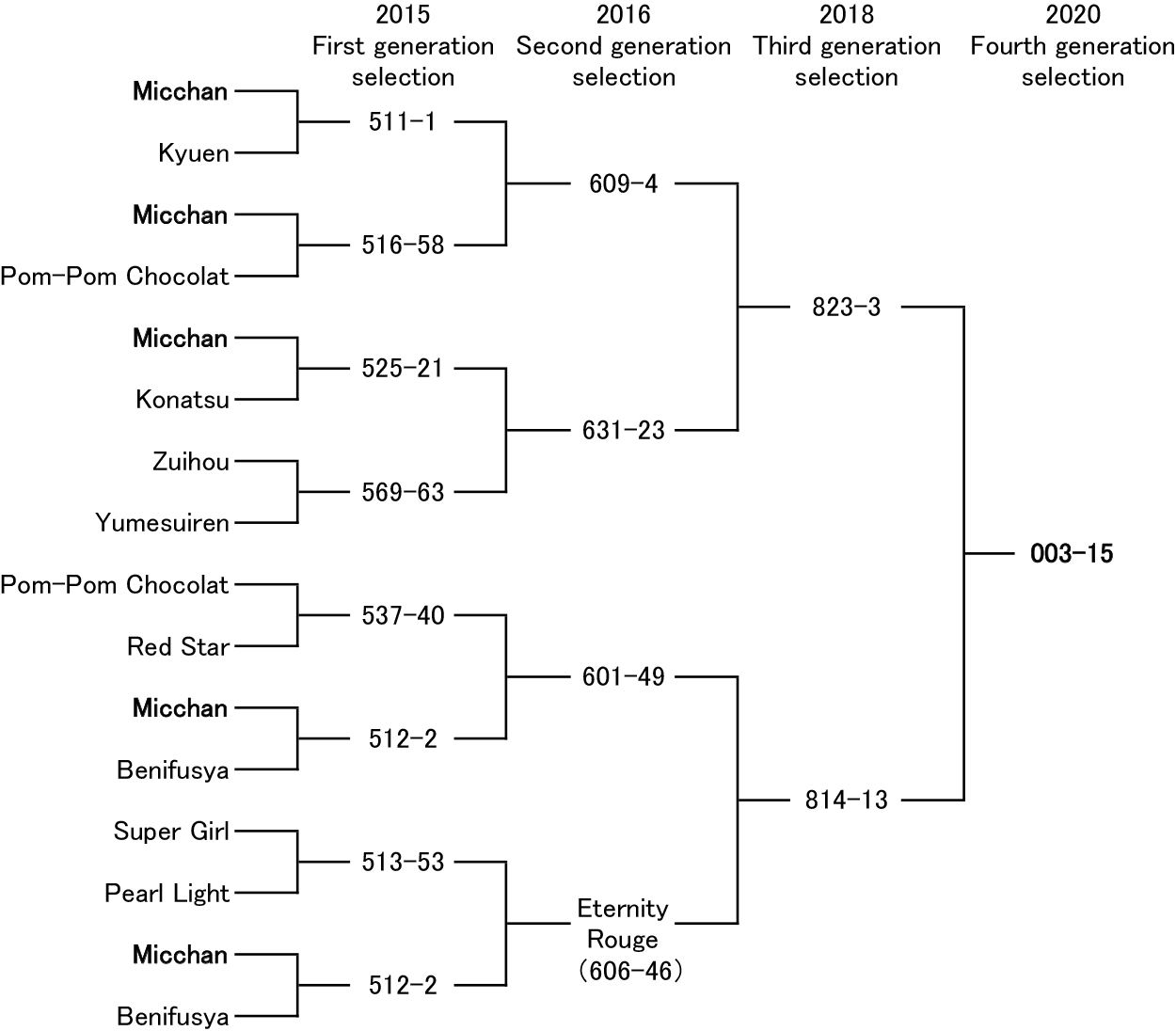
Our previous results strongly suggest that ‘Micchan’ has genes related to long vase life, and that the trait is heritable (Onozaki and Azuma, 2019). We traced the breeding parents of line 003-15 and found that ‘Micchan’ was frequently used as the breeding seed parent (Fig. 6). In the crossings for the 1st generation, 5 of the 16 parents were ‘Micchan’ (Fig. 6), so 31.25% of the total genome in line 003-15 is derived from ‘Micchan’. Dahlias show huge variation in flower traits such as color, shape, and size, owing to their high polyploidy (Okumura and Fujino, 1989; Wegner and Debener, 2008), likely auto-allo-octoploidy (2n = 8x = 64) (Gatt et al., 1998). This complex genome can produce various allele patterns. Our results suggest that the genes related to long vase life derived from ‘Micchan’ may be accumulated or duplicated in line 003-15.
Dahlias have a wide variety of flower types and are classified by the Royal Horticultural Society into 14 groups accordingly (Roy, 2019). Our initial breeding materials of 22 commercial dahlia cultivars consisted of five different flower types (Onozaki and Azuma, 2019). However, most selected lines with long vase life had well arranged flower types, such as formal decorative. ‘Eternity Torch’ and line 816-7 are valuable as semi-cactus blooming types. However, flower types with unusual petal shapes such as inward curved or twisted did not appear (Fig. 4; Tables 4 and 5). Therefore, selection and crossing for long vase life seems to reduce flower type diversity. This may be due to links between the genes related to long vase life derived from ‘Micchan’ and genes related to well arranged flower types, such as formal decorative and ball.
In Japanese morning glory (Ipomoea nil (L.) Roth), a NAC (NAM/ATAF1,2/CUC2) transcription factor, EPHEMERAL1, is a key regulator of ethylene-independent petal senescence (Shibuya, 2018; Shibuya et al., 2014). Recently, we identified three NAC transcription factors likely to be involved in senescence in the Eternity series (Nishio et al., 2022). We plan to identify the senescence-related genes in our final selected lines.
In conclusion, we further improved dahlia flower vase life by means of conventional cross-breeding techniques, accomplishing a significant genetic improvement by the 5th generation. We identified line 003-15 with ultra-long vase life in the 4th generation as the optimal line. Flower trade is increasingly a global business, with high-quality Japanese cut flowers a valuable export item. Therefore, cut flower transportability is an increasingly important trait. Breeding dahlia cultivars with both longer vase life and ethylene resistance enables handling in ethylene-contaminated environments. To achieve this, we initiated another breeding program to develop ethylene-resistant dahlias, and selected three lines (Fujimoto and Onozaki, 2022). We also plan further crossings between selected 4th- or 5th-generation lines and developed ethylene-resistant lines.
In the autumn of 2021, we crossed 20 primary-selected 5th-generation lines in 13 combinations, but obtained almost no 6th-generation progeny (data not shown). Dahlias are generally known to have self-incompatibility (Miyoshi, 2018; Roy, 2019). Dahlia coccinea, one cultivated dahlia progenitor (Okumura and Fujino, 1989), is self-incompatible and depends completely on diurnal visitors for pollination (Figueroa-Castro and Cano-Santana, 2011). On the other hand, Behr and Debener (2004) reported two self-fertile dahlia genotypes. It is possible that our 5th generation breeding lines became genetically inbred. As the influence of weather and temperature at the time of crossing may also have influenced the crosses in autumn 2021, we plan to verify the crossbreeding feasibility among the 5th-generation selected lines.
Acknowledgements
We sincerely thank Ms. A. Yamagata, Dr. M. Mato (Akita Agricultural Experiment Station), Mr. N. Tsujimoto, Mr. K. Inda (Nara Agricultural Research Center), Mr. S. Yamashita, Ms. M. Hiraishi (Kochi Agricultural Research Center), Mr. K. Okishio, Mr. H. Nakamura, Ms. M. Kurogi, and Dr. K. Nakamura (Miyazaki Agricultural Research Institute) for their contributions to the trial production of lines selected for release. We also thank the members of the Technical Support Center of Central Region, Tsukuba Operation Unit 6, Fujimoto-Owashi Technical Team, NARO, for their technical support in field and greenhouse management.
Literature Cited
- Azuma, M., T. Onozaki and K. Ichimura. 2019. Effects of bacterial proliferation and soluble carbohydrate levels on the vase life of cut dahlia (Dahlia variabilis) flowers. Hort. J. 88: 106–115.
- Azuma, M., T. Onozaki and K. Ichimura. 2020. Difference of ethylene production and response to ethylene in cut flowers of dahlia (Dahlia variabilis) cultivars. Sci. Hortic. 273: 109635. DOI: 10.1016/j.scienta.2020.109635.
- Behr, H. and T. Debener. 2004. Novel breeding strategies for ornamental dahlias I: Analysis of the Dahlia variabilis breeding system with molecular markers. Eur. J. Hort. Sci. 69: 177–183.
- Boxriker, M., R. Boehm, J. Möhring and H. P. Piepho. 2017. Efficient statistical design in two-phase experiments on vase life in carnations (Dianthus caryophyllus L.). Postharvest Biol. Technol. 128: 161–168.
- Boxriker, M., J. Möhring and H. P. Piepho. 2018. Genetic and phenotypic correlation for breeding relevant traits in Dianthus caryophyllus L. Postharvest Biol. Technol. 143: 129–136.
- Carvalho, D. R. A., C. F. S. Koning-Boucoiran, D. Fanourakis, M. W. Vasconcelos, S. M. P. Carvalho, E. Heuvelink, F. A. Krens and C. Maliepaard. 2015. QTL analysis for stomatal functioning in tetraploid Rosa × hybrida grown at high relative air humidity and its implications on postharvest longevity. Mol. Breed. 35: 172. DOI: 10.1007/s11032-015-0365-7.
- Fanourakis, D., D. R. A. Carvalho, V. W. Gitonga, A. W. van Heusden, D. P. F. Almeida, E. Heuvelink and S. M. P. Carvalho. 2012. Breeding cut roses for better keeping quality: first steps. Acta Hortic. 937: 875–882.
- Figueroa-Castro, D. M. and Z. Cano-Santana. 2011. Effectiveness of vectors of pollen and longevity of capitula for four species of Asteraceae in central Mexico. Southwestern Naturalist 56: 162–171.
- Fujimoto, T. and T. Onozaki. 2022. Breeding for improvement of flower life in dahlia. 8. Ethylene sensitivity of crossed seedlings between different ethylene sensitivity parents and selection of lines with exogenous ethylene resistance. Hort. Res. (Japan) 21 (Suppl. 2): 215 (In Japanese).
- Gatt, M., H. Ding, K. Hammett and B. Murray. 1998. Polyploidy and evolution in wild and cultivated Dahlia species. Ann. Bot. 81: 647–656.
- Giovannini, A., M. Laura, B. Nesi, M. Savona and T. Cardi. 2021. Genes and genome editing tools for breeding desirable phenotypes in ornamentals. Plant Cell Rep. 40: 461–478.
- Hotta, M., H. Hattori, T. Hirano, T. Kume, Y. Okumura, K. Inubushi, Y. Inayoshi, M. Ninura, J. Matsuno, T. Onozaki, M. Yagi, H. Yamaguchi and N. Yamaguchi. 2016. Breeding and characteristics of spray-type carnation ‘Kaneainou 1 go’ with a long vase life. Res. Bull. Aichi Agric. Res. Ctr. 48: 63–71 (In Japanese with English abstract).
- Ichimura, K., Y. Kawabata, M. Kishimoto, R. Goto and K. Yamada. 2002. Variation with the cultivar in the vase life of cut rose flowers. Bull. Natl. Inst. Flor. Sci. 2: 9–20.
- Ichimura, K., H. Shimizu-Yumoto, K. Shibuya and H. Mochizuki-Kawai. 2011. Investigation of the vase life of cut flowers in different seasons. Bull. Natl. Inst. Flor. Sci. 11: 49–65 (In Japanese with English abstract).
- Ichimura, K., M. Taguchi and R. Norikoshi. 2006. Extension of the vase life in cut roses by treatment with glucose, isothiazolinonic germicide, citric acid and aluminum sulphate solution. JARQ 40: 263–269.
- Konishi, Y. 2009. Flower breeding of dahlia—Looking for new flowers. p. 167–171. In: Japan Dahlia Society (ed.). Encyclopedia of Dahlia. Seibundo-Shinkosha, Tokyo (In Japanese).
- Miyoshi, K. 2018. Current status and future prospects in commercial cultivation of dahlia cut flowers (4). Agric. Hortic. (Noko-to-Engei) 73: 40–45 (In Japanese).
- Naka, T., H. Yumoto, N. Tsujimoto and Y. Torata. 2015. Seasonal variation on the vase life and sugar concentration of cut Dahlia flower. Hort. Res. (Japan). 14 (Suppl. 2): 266 (In Japanese).
- Nishio, Y., M. Azuma, H. Momose, T. Onozaki and A. Tateishi. 2022. Investigation of NAC transcription factors regulating flower senescence in Dahlia. Hort. Res. (Japan) 21 (Suppl. 2): 210 (In Japanese).
- Okumura, S. and M. Fujino. 1989. Dahlia Cav. p. 174–180. In: Y. Tsukamoto (ed.). The grand dictionary of horticulture Vol. 3. Shogakukan, Tokyo (In Japanese).
- Onozaki, T. 2018a. Breeding of carnations (Dianthus caryophyllus L.) for long vase life. Breed. Sci. 68: 3–13.
- Onozaki, T. 2018b. Breeding and production status and future development direction in dahlia. SHOKUCHO 52(8): 6–12 (In Japanese).
- Onozaki, T. and M. Azuma. 2019. Breeding for long vase life in dahlia (Dahlia variabilis) cut flowers. Hort. J. 88: 521–534.
- Onozaki, T. and M. Azuma. 2022. Development of dahlia cultivars ‘Eternity Torch’, ‘Eternity Romance’ and ‘Eternity Rouge’ with long vase life. J. NARO Res. Dev. 10: 1–19 (In Japanese with English abstract).
- Onozaki, T., H. Ikeda, M. Shibata, N. Tanikawa, M. Yagi, T. Yamaguchi and M. Amano. 2006b. Breeding and characteristics of carnation Norin No. 1 ‘Miracle Rouge’ and No. 2 ‘Miracle Symphony’ with long vase life. Bull. Natl. Inst. Flor. Sci. 5: 1–16 (In Japanese with English abstract).
- Onozaki, T., H. Ikeda and T. Yamaguchi. 2001. Genetic improvement of vase life of carnation flowers by crossing and selection. Sci. Hortic. 87: 107–120.
- Onozaki, T., N. Tanikawa, M. Yagi, H. Ikeda, K. Sumitomo and M. Shibata. 2006a. Breeding of carnations (Dianthus caryophyllus L.) for long vase life and rapid decrease in ethylene sensitivity of flowers after anthesis. J. Japan. Soc. Hort. Sci. 75: 256–263.
- Onozaki, T., M. Yagi and K. Tanase. 2015. Selection of carnation line 806-46b with both ultra-long vase life and ethylene resistance. Hort. J. 84: 58–68.
- Onozaki, T., M. Yagi, K. Tanase and M. Shibata. 2011. Crossings and selections for six generations based on flower vase life create lines with ethylene resistance or ultra-long vase life in carnations (Dianthus caryophyllus L.). J. Japan. Soc. Hort. Sci. 80: 486–498.
- Roy, D. 2019. Dahlia. p. 27–30. In: D. Roy (ed.). Breeding of ornamental crops. Alpha Science International Ltd. Oxford, U.K.
- Saar, D. E., N. O. Polans and P. D. Sørensen. 2003. A phylogenetic analysis of the genus Dahlia (Asteraceae) based on internal and external transcribed spacer regions of nuclear ribosomal DNA. Syst. Bot. 28: 627–639.
- Shibuya, K. 2018. Molecular aspects of flower senescence and strategies to improve flower longevity. Breed. Sci. 68: 99–108.
- Shibuya, K., K. Shimizu, T. Niki and K. Ichimura. 2014. Identification of a NAC transcription factor, EPHEMERAL1, that controls petal senescence in Japanese morning glory. Plant J. 79: 1044–1051.
- Shimizu-Yumoto, H. 2018. Postharvest characteristics of cut flowers and techniques for extending vase life, with a focus on Eustoma, Gentiana and Dahlia. AGri-Bioscience Monographs 8: 1–22.
- Shimizu-Yumoto, H. and K. Ichimura. 2013. Postharvest characteristics of cut dahlia flowers with a focus on ethylene and effectiveness of 6-benzylaminopurine treatments in extending vase life. Postharvest Biol. Technol. 86: 479–486.
- Shimizu-Yumoto, H., N. Tsujimoto and T. Naka. 2020. Acid invertase activities of dahlia ‘Kokucho’ petals during flower opening and following cutting and treatment with 6-benzylaminopurine. Sci. Hortic. 272: 109525. DOI: 10.1016/j.scienta.2020.109525.
- Tsujimoto, N., T. Naka, Y. Torata, H. Yumoto and A. Higashi. 2016. Cultivar variation in the extending effect of 6-benzylaminopurine (BA) preparation on dahlia vase life. Bull. Nara Agric. Res. Cen. 47: 11–17 (In Japanese with English abstract).
- Van der Meulen-Muisers, J. J. M., J. C. van Oeveren, J. Jansen and J. M. van Tuyl. 1999. Genetic analysis of postharvest flower longevity in Asiatic hybrid lilies. Euphytica 107: 149–157.
- Van Geest, G., A. Post, P. Arens, R. G. F. Visser and U. van Meeteren. 2017. Breeding for postharvest performance in chrysanthemum by selection against storage-induced degreening of disk florets. Postharvest Biol. Technol. 124: 45–53.
- Wegner, H. and T. Debener. 2008. Novel breeding strategies for ornamental dahlias II: Molecular analyses of genetic distances between Dahlia cultivars and wild species. Europ. J. Hort. Sci. 73: 97–103.
- Wernett, H. C., G. J. Wilfret, T. J. Sheehan, F. J. Marousky, P. M. Lyrene and D. A. Knauft. 1996. Postharvest longevity of cutflower Gerbera. I. response to selection for vase life components. J. Amer. Soc. Hort. Sci. 121: 216–221.
- Yamagata, A. 2022. Dahlia technology system in cool regions. p. 232–246. In: Saishin Nougyou Gijyutsu kaki Vol. 14. Noubunkyo, Tokyo (In Japanese).
- Yang, Y., S. Takenaga, S. Ohno and M. Doi. 2021. Ethylene-sensitive abscission layer formation and petal senescence in cut dahlia inflorescences. Hort. J. 90: 460–468.