Abstract
Chili pepper is cultivated in the absence of fruit-promoting treatments or insect pollinators. The fertilization ability of both males and females, as well as natural self-pollination ability under high temperatures, are necessary for autonomous set fruit under high temperatures. The reproductive traits related to autonomous self-pollination ability and fertilization ability of both males and females after pollination under control (CK) and high temperature (HT) treatments were investigated in this study. ‘Takanotsume’ (TK) showed a significantly higher percentage of autonomous fruit set than ‘Peruvian Purple’ (PP) in both CK and HT treatments, suggesting that TK has a strong autonomous fruit set regardless of temperature. On the other hand, the percentage of autonomous fruit set of PP was 0% in the HT treatment, while autonomous fruit set was observed in the CK treatment. Therefore, TK had autonomous fruit set ability even at high temperatures when PP did not have any. TK had more pollen on the stigma than PP, suggesting that TK has a higher ability for autonomous self-pollination. Flower morphology was not considered a factor in the autonomous self-pollination of TK. In contrast, TK tended to release more pollen than PP. One factor that can support autonomous self-pollination is pollen dispersion. A significant difference was observed in the fruit set rate during artificial pollination between the treatments (CK and HT) of the pollen parent and the cultivars (TK and PP) of the seed parent. In fact, female fertility in TK was higher than that in PP, regardless of the temperature condition. Almost no fruit set of PP was observed via autonomous self-pollination in the HT treatment, but fruits were set by artificial pollination using both the pollen and pistil of PP in the HT treatment. This result is consistent with the finding that PP exhibits decreased autonomous self-pollination at high temperatures. The higher autonomous fruit-set ability of TK than PP under high temperatures may be attributed not only to superior female fertility, but also to its autonomous self-pollination ability under high temperatures. Therefore, pollen dispersal ability under high temperatures was considered a key factor for autonomous fruit-set.
Introduction
The genus Capsicum includes important crops that are used worldwide as spices, vegetables, or ornamental plants. The genus belongs to the Solanaceae family and consists of several wild and five domesticated species (Bosland and Votava, 2000). C. annuum is a particularly important species as it is consumed by 25% of the world’s population as a vegetable, spice, or food coloring (Smith, 2015). C. annuum is mainly cultivated in temperate areas, including Japan.
Owing to global warming, the air temperature is currently increasing rapidly (IPCC, 2022). Plant fertility decreases with increasing air temperature in many areas (Driedonks et al., 2016; Hedhly et al., 2008; Mesihovic et al., 2016). Many agricultural crops, including vegetables, are expected to produce lower yields. In fact, high temperatures above 33°C for C. annuum cause poor fruit setting (Erickson and Markhart, 2002). Therefore, it is increasingly important to implement countermeasures against yield loss at high temperatures. There is a strong demand for breeding programs that produce stable yield, even at high temperatures.
Heat stress can affect various reproductive abilities (Hedhly et al., 2008), such as pollen release (Sato et al., 2000), pollen fertility (Dane et al., 1991; Iwahori, 1965; Peet et al., 1998; Sato et al., 2006), pollen germination rate (Sato et al., 2000; Yamazaki and Hosokawa, 2019), pollen number (Xu et al., 2017), pollen tube elongation, and female fertility (Xu et al., 2017). In addition, high-temperature stress causes morphological abnormalities in flowers and affects self-pollination. Under high temperatures, the total length of stamens becomes shorter, and the distance between the anther and stigma increases in Arabidopsis thaliana (Katano et al., 2020). In tomato (Solanum lecopersicum), the pistil becomes longer than the stamen-column, and the stigma is exposed from the stamen-column, making self-pollination difficult at high temperatures (Levy et al., 1978; Smith, 1932). These factors are thought to limit fruit set under high temperatures.
Autonomous self-pollination that occurs without pollen vectors is called “automatic self-pollination”, “autonomous autogamy”, (Richards, 1986) or “autofertility” (Yashiro et al., 1999). “Autonomous fruit set” refers to autonomous self-pollination and fruit-set without wind or insect pollinators. Solanaceae vegetables, such as tomatoes and chili peppers, have autonomous fruit set ability. These plants can set fruits even in greenhouses without any treatment to promote fruit set. Chili peppers appear to have a strong autonomous fruit set ability because they are cultivated by farmers without insect pollinators or vibration pollination. Therefore, it is important to leverage autonomous fruit-set for the fruit production of chili peppers in greenhouse horticulture.
Even chili peppers cannot maintain their ability to autonomously set fruits in greenhouses under high temperatures. We discovered an F1 hybrid of C. chinense with higher autonomous fruit set ability, even at high temperatures (Yamazaki and Hosokawa, 2019). The F1 hybrid maintained pollen germination even at high temperatures, which was thought to lead to autonomous fruit set. However, there may be many traits related to autonomous fruit set under high temperatures. To understand the mechanism that maintains autonomous fruit set even at high temperatures, it is necessary to investigate the traits of genetic resources that maintain autonomous fruit set even under such temperatures.
In a previous study, we discovered that C. annuum ‘Takanotsume (TK)’ has a highly autonomous fruit set under high temperatures (Yamazaki et al., 2022b). TK is a cultivar that has been cultivated in Japan for a long time, and is native to Hamamatsu, Shizuoka Prefecture (Kumazawa et al., 1954). TK has a long harvest period and many flowers and is currently the most popular high-yielding cultivar in Japan. By clarifying the fruiting mechanism under high temperatures, it may be possible to obtain information to improve the yield of chili peppers under such temperatures. The purpose of this study was to investigate the factors influencing autonomous fruit setting under high temperatures in TK. We investigated reproductive traits related to high-temperature autonomous fruit set, such as the pollen germination rate, female fertility, and autonomous self-pollination. In addition, flower morphology, pollen count, and pollen dispersal ability, which are thought to be related to autonomous self-pollination, were also investigated.
Materials and Methods
Plant materials and growth conditions
C. annuum ‘Takanotsume (TK)’ and ‘Peruvian Purple (PP)’ were sown on seeding rockwool culture mats (Nippon Rockwool, Tokyo, Japan) and placed in a phytotron (5:00–19:00, 28°C; 19:00–5:00, 20°C, natural day length). The control cultivar, PP, does not have autonomous fruit set ability during the high temperature season, and has a compact plant form, making it suitable for cultivation in an incubator. The seedlings were transplanted into 8 cm diameter black plastic pots filled with culture soil (Kumiai Nippi; Nihon Hiryo, Tokyo, Japan) two months after sowing and raised for another month. The plants were placed in incubators (LH-240SP; Nippon Medical and Chemical Instruments, Osaka, Japan). The day length was set to 12 h in incubators that were fully equipped with daylight color fluorescent lamps (FLR20SD/M; NEC, Tokyo, Japan) at a photosynthetic photon flux density (PPFD) of 250 μmol·m−2·s−1. The air temperature during the dark period was set at 20°C, and the temperature during the light period was set at 30°C in the control plot (CK). In the high temperature treatment plot (HT), the temperature was set at 35°C for 6 h in the middle of the light period (Table 1), based on the procedure of Yamazaki and Hosokawa (2020). Two individuals of each cultivar and each treatment were used for the following investigations, 21 days after cultivation in incubators.

Branches with unflowered buds were marked with masking tape on the first day of the survey. The branches were harvested at marked positions after four weeks. The numbers of fruits and fallen flowers were counted. The percentage of autonomous fruit set was calculated based on the number of flowers and fruits. Two individuals of each cultivar and treatment were used for the investigations, and three replicated surveys were conducted. Five branches from each individual plant were investigated.
Evaluation of the pollen numbers on the stigma with autonomous self-pollination
The flowers were collected one day after flower opening. Pistils were picked from the collected flowers using tweezers and placed on an agar medium. Pistils on the agar medium were observed under a stereoscopic microscope (SZX7-TR30-2-SET; Olympus, Tokyo, Japan), and the number of pollen grains on the stigma was visually counted. The stigmas were stained with 0.05% (w/v) toluidine blue staining solution as necessary. Twenty flowers were investigated per replicate for both the cultivars and treatments.
Evaluation of total pollen numbers in the anthers
Anthers were collected from each flower on the day the flower opened. Pollens were suspended in 100 μL of pollen culture solution prepared by dissolving 100 ppm boric acid and 5% sucrose in distilled water. Pollen suspensions in a Thoma hemocytometer (03-101-1; Erma, Saitama, Japan) were observed under a biological microscope (BX51N-34-FLD-1; Olympus), and digital images were obtained using a CCD camera (DP71; Olympus). The total pollen number was counted using a hemocytometer, and the pollen density was calculated. The pollen density in the suspension was multiplied by the volume (100 μL) of the solution used to suspend one flower to determine the total number of pollen grains per flower. Ten flowers from each cultivar and treatment were investigated, and the pollen count on the hemocytometer was replicated three times per flower.
Evaluation of pollen dispersal
The day before flower opening, flower buds were put in 1.5 mL microtubes. The anther dehiscence stage was recorded on the day the flower opened as described by Yamazaki et al. (2022b). The microtubes were collected one day after flower opening inside the microtubes. The pollen culture medium (50 μL) was added to the microtubes and suspended. A 10 μL sample was dropped onto the hemocytometer, and the numbers of released pollen per flower were calculated as described above. Ten flowers from each cultivar and treatment plot were tested, and the hemocytometer count per flower was repeated three times.
Evaluation of flower morphology
The lengths of anthers, filaments, stamens, and styles were measured using electronic calipers (AD-5765A150; A&D, Tokyo, Japan) on the day the flower opened. We calculated the difference between the total length of the pistil and the total length of the stamen. The degree of anther dehiscence was classified into stages 1 to 4, according to the procedure described by Yamazaki et al. (2022b) as follows: Degree of anther dehiscence 1, does not dehisce at all; degree of anther dehiscence 2, 3, and 4, dehisces only at the tip, half, or whole, respectively. Each cultivar and treatment group was investigated, with 20 flowers per replicate.
Fruit set after artificial self- or cross-pollination
For artificial pollination, pollen was collected from the flowers on the day they opened. Flowers with pistils were emasculated one day before flower opening. Sixteen combinations of two cultivars (TK and PP) and two temperature treatments (CK and HT) of pollen and pistils were investigated. The percentage of fruit set was determined two weeks after artificial pollination. Fifteen flowers were investigated for each treatment and cultivar.
Observation of pollen tube elongation in the pistils
Stamens were removed from the buds one day before flower opening, and artificial self- or cross-pollination was performed using pollen collected from the flowers on the day they opened. Pistils were collected 24 h after artificial pollination, followed by fixation in acetic acid: ethanol (1:3 (v/v)) for 1 h. The pistils were immersed in 8N NaOH at 20°C, overnight. Pollen tube elongation in the pistil was observed under a biological microscope after staining with 0.1% aniline blue in 0.1 M H3PO4. BP330-385 and BA420 were used as excitation and absorption filters, respectively. Sixteen combinations of pollination using pollen parent and seed parent of two cultivars (TK or PP) and two temperature treatments (CK or HT) were observed, with nine flowers each.
Seed numbers
The numbers of seeds and ovules that did not become seeds were counted from each cultivar and treated in fruits set by autonomous self-pollination. A stereoscopic microscope (SZX7-TR30-2-SET; Olympus) was used to count dead ovules. Fifteen fruits from each cultivar and treatment were investigated. However, the number of fruit sets was extremely small in the HT of PP and only four fruits were investigated.
Pollen germination
The liquid medium was prepared by dissolving 100 ppm of boric acid and 5% sucrose in distilled water. Anthers were added to the liquid medium, and the pollen was suspended. The agar medium was prepared by adding 1% agar to the liquid medium in a plastic petri dish. The pollen suspension was spread on the agar medium and left overnight in an incubator at 25 or 35°C in the dark. Pollen germination on the pollen medium was observed using a biological microscope at 40x. Images of pollen germination were captured using a CCD camera (DP71; Olympus) attached to a microscope. The numbers of germinated and non-germinated pollen grains were counted using the image analysis method constructed in a previous study (Yamazaki et al., 2022a), and the pollen germination rate was calculated. Eighteen flowers were investigated for each cultivar and treatment group.
Histological analysis of anthers
Flowers (buds) were classified according to the stages of flower bud development by Erickson and Markhart (2002) as follows: Stage 1, calyx lobes closed, petal is not visible with green buds; Stage 2, calyx lobes begin to separate; Stage 3, calyx continues to separate and petals become visible; Stage 4, petals protrude; Stage 5, petals protrude almost twice as long as the sepals. Anthers at each flower bud development stage were fixed in 3% (v/v) glutaraldehyde in 50 mM Na-Pi (pH 6.8) at 4°C overnight. The fixed buds were embedded in SCEM (Section Lab, Yokohama, Japan) and frozen in dry ice-chilled hexane. The embedded blocks were sliced to a thickness of 6 μm using a cryostat (CM3050S; Leica, Nussloch, Germany), according to the procedure described by Kawamoto (2003). Cross-sections were stained with toluidine blue O. Images of anther tissues were obtained using a biological microscope (BX51N-34-FLD-1; Olympus) and a CCD camera (DP71; Olympus) attached to the microscope.
Pollen fertility
Anthers were collected from one flower on the day the flower opened, added to 100 μL of acetocarmine, and suspended. Pollen grains were observed under a biological microscope (BX51N-34-FLD-1; Olympus). Three pollen images were captured for each sample using a CCD camera (DP71; Olympus) attached to the microscope. Fertile and sterile pollen from the images were measured to determine pollen fertility. Twenty flowers from each cultivar and treatment were investigated.
Statistical analysis
Statistical analyses including Student’s t-test, Tukey-Kramer test, analysis of variance (ANOVA), Kruskal-Wallis rank sum test and correlation analysis were performed using R ver.4.0.3 (R core team, 2020).
Results
Autonomous fruit set
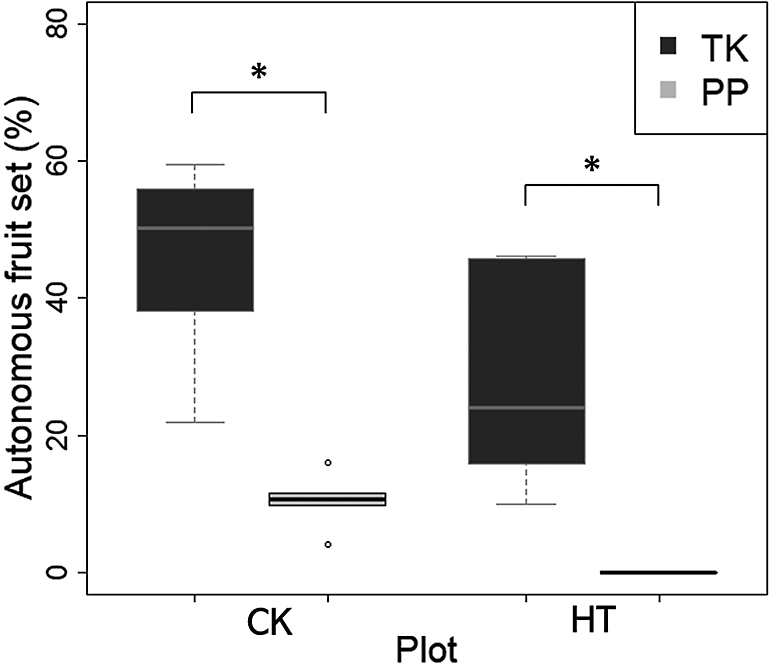
TK had a significantly higher percentage of autonomous fruit set than PP in both CK and HT (Fig. 1). Autonomous fruit set of PP was observed in CK; however, the percentage of autonomous fruit set was 0% in HT (Fig. 1). TK produced a lot of fruit set, regardless of temperature. In addition, TK had autonomous fruit-set ability even at high temperatures when PP did not have any fruit set.
Autonomous self-pollination
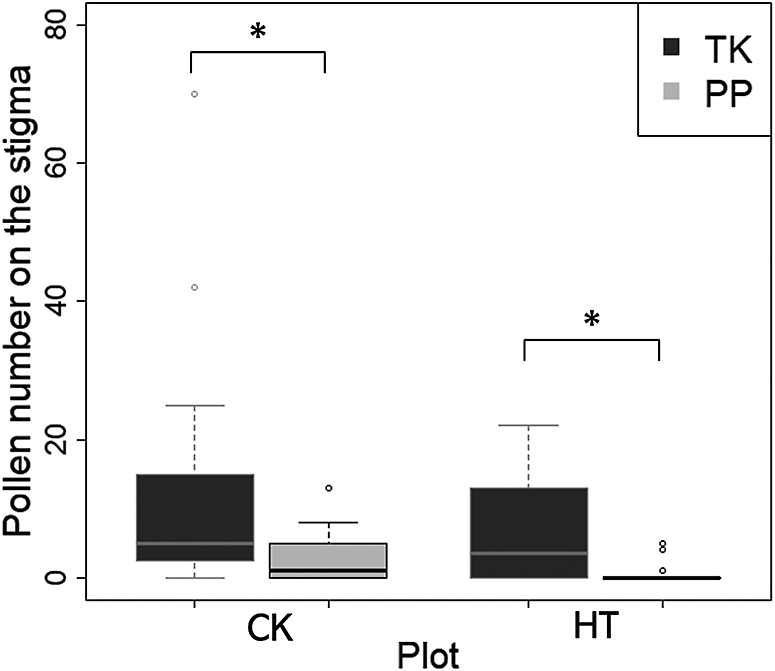
The pollen numbers on the stigma with autonomous self-pollination in TK were significantly higher than those in PP in both CK and HT (Fig. 2). In particular, the pollen number on the stigma of PP was 0.55 in HT, suggesting that almost no pollen was pollinated on the stigma under high temperatures. In contrast, although TK slightly reduced the number of pollens pollinated by autonomous self-pollination in HT compared to that in CK, these numbers were about the same. In particular, under high temperatures, TK was more likely than PP to maintain the ability to self-pollinate autonomously.
Flower morphology
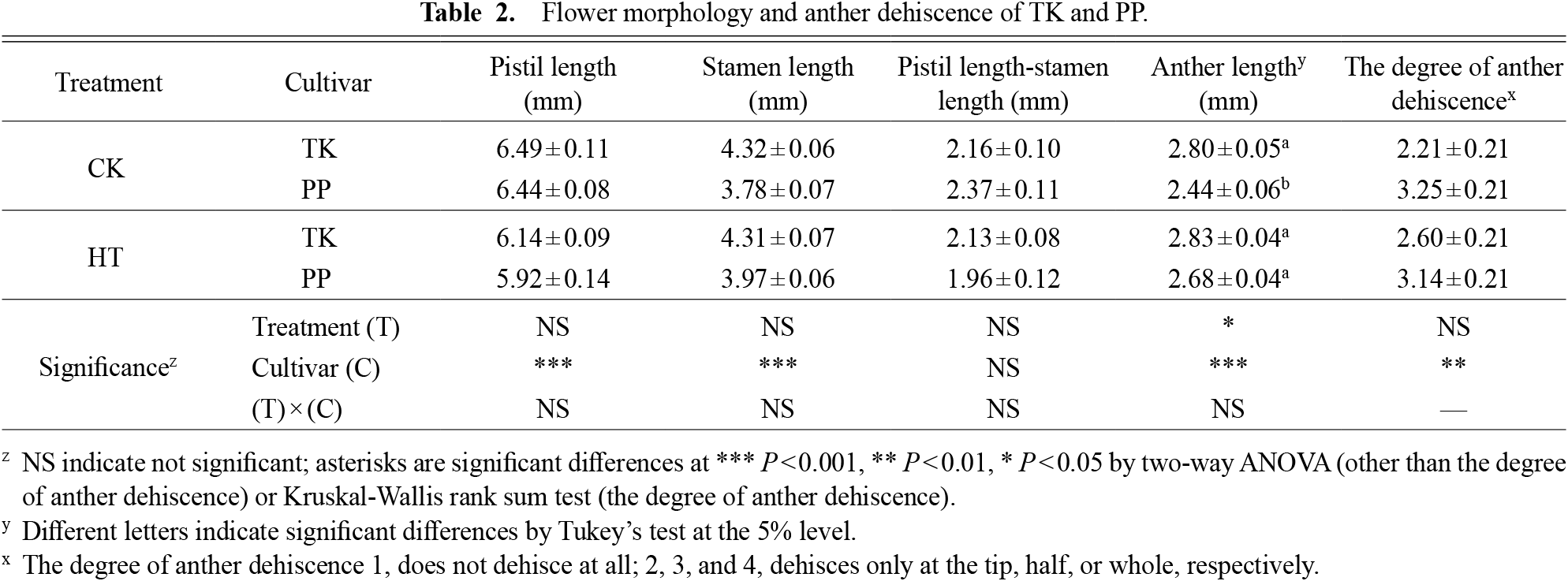
There were no significant differences in the pistil length or the total stamen length between the treatments (HT or CK), but there were significant differences between the cultivars (Table 2). The pistil length and the total stamen length of TK were longer than those of PP. In contrast, no significant difference was observed in the distance between the anther and stigma between cultivars and treatments (Table 2). The distance between the anther and stigma was not considered a factor in autonomous self-pollination of TK. There were significant differences in the anther size between the two treatments and cultivars. The anther size of PP was significantly larger than that of TK in CK (Table 2). There was a significant difference in the anther dehiscence only between the cultivars. The degree of TK anther dehiscence was slightly lower than that of PP in both CK and HT (Table 2). Therefore, the degree of anther dehiscence and anther size were not considered as factors in the autonomous self-pollination ability of TK.
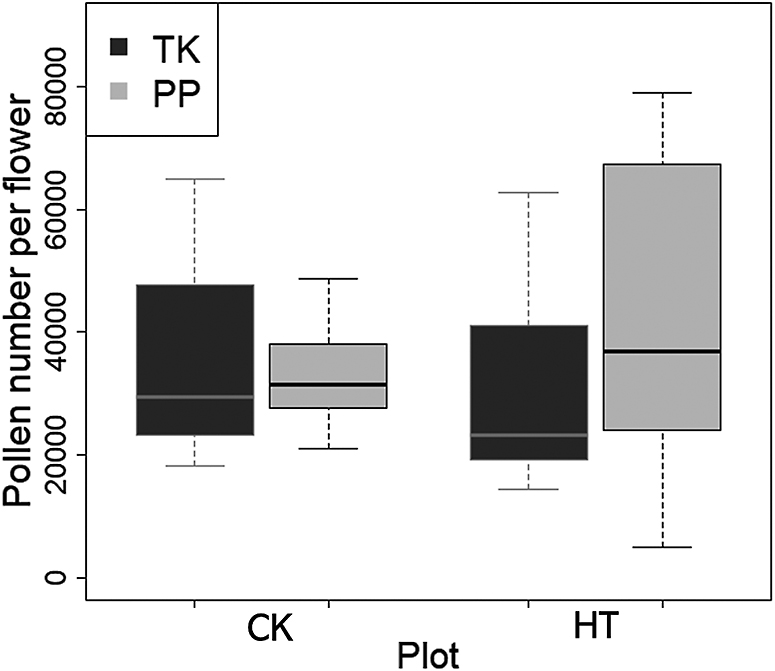
No significant difference was observed in the number of pollen produced in flowers between the cultivars in the HT and CK treatments (Fig. 3). Although there was no statistically significant difference, the pollen number in HT was slightly higher in PP than that in TK.
Pollen release
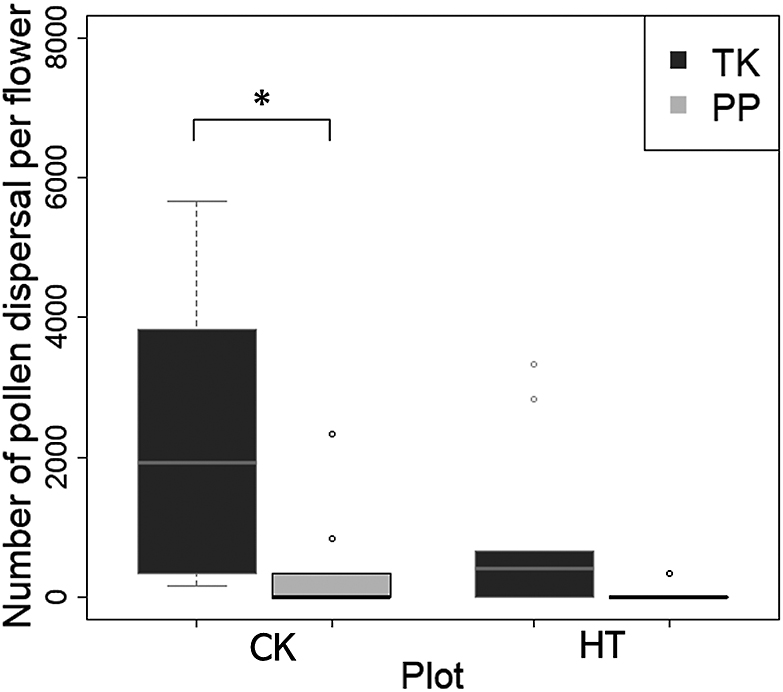
TK had a significantly higher number of pollens released per flower than PP in CK (Fig. 4). In HT, the mean value of PP pollen release was 33.3, whereas that of TK was 833; however, this was not statistically significant. Pollen release hardly occurred in PP because of high-temperature stress, whereas it did occur in TK. No correlation was found between the number of pollens released and the degree of anther dehiscence (Fig. S2). Therefore, the high autonomous self-pollination ability of TK is related to pollen dispersal, which does not depend on the degree of anther dehiscence or the pollen number in the anthers.
Fruit set after artificial self- or cross-pollination
Figure 5 shows the percentages of fruit set with artificial self- and cross-pollination. A four-way ANOVA of the percentage of fruit set with artificial pollination showed significant differences in the factors of pistil cultivars and pollen treatments (Table S1). This result suggests that the female fertility of TK was higher than that of PP regardless of the temperature treatment. This may explain why TK had a higher autonomous fruit set than PP in CK. The pollen fertilization ability of both cultivars decreased at high temperatures. Almost no autonomously set PP fruit was observed under HT (Fig. 1); nonetheless, fruit set was observed with artificial self-pollination under HT (Fig. 5). Autonomous self-pollination is a factor limiting fruit set when there are no pollen vectors, such as wind or pollinator insects.
Pollen tube elongation in the style
The results for pollen tube elongation within the pistil are shown in Figure S1 and Table S2. Pollen tube elongation was observed in almost all the pollination combinations. In contrast, fewer pistils with elongated pollen tubes were pollinated by PP pollen in HT. No difference was observed in the number of pollen tubes between male and female cultivars. Thus, the degree of pollen tube elongation within the pistil was not considered a factor in the autonomous fruit set in TK.
Pollen fertility, pollen germination, and histological analysis
No significant differences in pollen fertility were observed between the cultivars in either CK or HT (Fig. S3). In HT, the TK pollen germination rate was significantly higher than that of PP at a culture temperature of 25°C (Fig. 6). In CK, no significant difference was noted in pollen germination rates between cultivars at a culture temperature of 25°C. No significant difference was observed in pollen germination rates between cultivars and treatments at a culture temperature of 35°C (Fig. 6). Both cultivars had slightly lower pollen germination rates at a culture temperature of 35°C than at 25°C. This result suggests that the pollen germinated mainly at night when the temperature drops during the high-temperature season. Anther tissue observations showed no difference in stages I to III between treatments, and pollen tetrads were formed normally (Fig. 7). In contrast, in stages IV and V, abnormal pollen development was observed in the two cultivars under HT conditions (Fig. 7). Erickson and Markhart (2002) reported that high temperatures affect pollen stages after the formation of pollen tetrads. Our results are consistent with this report. No difference in pollen development between the cultivars was observed, suggesting that both cultivars were affected by high temperatures.

The ovule number of PP was significantly larger in CK than in TK, and slightly larger in HT (Fig. S4A). No significant difference was noted in the number of ovules between the treatments, suggesting that ovule number did not decrease under high temperatures. Nonetheless, the seed number of TK was significantly larger than that of PP in HT and tended to be larger in CK (Fig. S4B). This was because the pollen numbers of TK pollinated by autonomous self-pollination were larger than those of PP (Fig. 2). The minimum number of seeds per fruit was two for TK and four for PP, suggesting that there was no difference in the number of seeds required for fruit set.
Discussion
Causative traits of autonomous fruit set under high temperatures
TK showed a significantly higher percentage of autonomous fruit set than PP in both treatments (Fig. 1). On the other hand, under high temperatures, PP did not set fruit autonomously at all. We investigated the reproductive traits that lead TK to maintain autonomous fruit-set ability, even at high temperatures (Fig. 8). TK maintained its autonomous self-pollination ability, even at high temperatures, compared to PP (Fig. 2). Furthermore, TK had higher female fertility than PP, regardless of the temperature conditions (Fig. 5; Table S1). These two factors may be involved in TK autonomous fruit set under high temperatures. The higher female fertility of TK could be indirectly related to autonomous fruit set under high temperatures, as female fertility was higher in both CK and HT. The decrease in autonomous self-pollination restricted autonomous fruit set under high temperatures (Figs. 1 and 5). Thus, autonomous self-pollination is important for the superior ability of TK to achieve autonomous fruit set under high temperatures. We did not find any factors in the flower morphology of TK that could cause more autonomous self-pollination (Table 2). High pollen dispersion may be one of the key factors that increase autonomous self-pollination under high temperatures in TK (Fig. 4). Previous studies have shown that the pollen germination rate of TK is higher than that of other cultivars, suggesting that this may be the cause of autonomous fruit set (Yamazaki et al., 2022b). In the present study, the pollen germination rate of TK was significantly higher than that of PP (Fig. 6). However, no significant difference between TK and PP pollen was observed in the rate of fruit set after artificial pollination, which suggests that the effect of the pollen germination rate on fruit set was limited.

TK showed higher pollen release than PP in the CK and HT treatments (Fig. 4). This may be related to the strong autonomous self-pollination ability of TK. However, in TK, pollen release was significantly reduced in HT compared to that in CK, whereas the autonomous self-pollinated pollen on the stigma did not change significantly between the treatments. This suggests that other factors are involved in TK’s ability to maintain autonomous self-pollination, even at high temperatures. In the histological analysis of anthers, no differences between cultivars were observed in the morphology of the pollen or anthers (Fig. 7). The spatial distance between the stigma and anther is important for autonomous self-pollination (Luo and Widmer, 2013; Opedal, 2018; Torang et al., 2017), but this distance changes under high temperatures, making autonomous self-pollination difficult. However, no significant difference was observed in the distance between the anthers and stigmas between the cultivars (Table 2). Thus, distance was not considered a factor in autonomous self-pollination. Pollens with lightweight and smooth surfaces are easily dispersed by wind and vibration (Ackerman, 2000). It is possible that there was a difference in the ultrastructure of TK and PP under HT, resulting in a difference in the autonomous self-pollination ability of TK and PP. We observed that the ultrastructures of the TK and PP stigmas were different (data not shown). The TK stigmas had a smooth surface and a hemispherical shape with a raised center, whereas the PP stigmas had an uneven surface and a depression in the center. Therefore, it is also possible that the difference in the ultrastructure of the stigma is also involved in the autonomous self-pollination ability of TK. Further observations and investigation of the stigma ultrastructure stigma and pollen surface are needed.
The effect of female reproductive ability for autonomous fruit-set
The higher female fertility of TK compared to PP in both the HT and CK treatments may be responsible for the higher autonomous fruit set ability regardless of temperature treatments (Fig. 5; Table S1). The rehydration of pollen on the stigma, pollen germination, pollen tube elongation in the style, and fertilization are required for fruit set after pollination. Artificial pollination and observation of pollen tube elongation in the pistil showed that there was no difference between TK and PP in terms of pollen receptivity of the stigma and pollen tube elongation in the pistil (Fig. S1; Table S2). During the process from pollination to pollen tube elongation, there was no causative trait for the high female fertility of TK. Therefore, there may be other factors involved in the process from fertilization to fruiting. In addition, the number of ovules was larger in PP, and the minimum number of seeds per fruit did not differ significantly among cultivars. These traits are not considered to be a factor in high female fertility. Therefore, the high female fertility in TK may be related to fertilization. During double fertilization, helper cells secrete pollen tube-attracting peptides that attract pollen tubes into the ovule (Higashiyama and Takeuchi, 2015). TK and PP may possess differences in ovule function, such as differences in the degree of pollen tube attraction; however, further research on female fertility is necessary.
Conclusion
By comparing each reproductive trait (Fig. 8), we clarified that the autonomous fruit set of TK under high temperatures could mainly be caused by maintenance of pollen dispersion. TK had a significantly higher pollen germination rate than PP in HT, although there was no difference between the cultivars in CK (Fig. 6). However, there was no difference in the percentage of fruit set between TK pollen and PP pollen after artificial pollination (Fig. 5), suggesting that pollen germination had a limited effect on autonomous fruit setting. The control cultivar, PP did not set autonomous fruit in HT (Fig. 1), nor did it self-pollinate autonomously (Fig. 2). The maintenance of autonomous self-pollination by pollen dispersal mainly induces the autonomous fruit setting of TK under high temperatures.
Literature Cited
- Ackerman, J. D. 2000. Abiotic pollen and pollination: ecological, functional, and evolutionary perspectives. Plant Syst. Evol. 222: 167–185.
- Bosland, P. W. and E. J. Votava. 2000. Peppers: vegetable and spice capsicums. CABI Publishing, New York.
- Dane, F., A. G. Hunter and O. L. Chambliss. 1991. Fruit set, pollen fertility, and combining ability of selected tomato genotypes under high-temperature field conditions. J. Amer. Soc. Hort. Sci. 116: 906–910.
- Driedonks, N., I. Rieu and W. H. Vriezen. 2016. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant Reprod. 29: 67–79.
- Erickson, A. N. and A. H. Markhart. 2002. Flower development stage and organ sensitivity of bell pepper (Capsicum annuum L.) to elevated temperature. Plant Cell Environ. 25: 123–130.
- Hedhly, A., J. I. Hormaza and M. Herrero. 2008. Global warming and sexual plant reproduction. Trends Plant Sci. 14: 30–36.
- Higashiyama, T. and H. Takeuchi. 2015. The mechanism and key molecules involved in pollen tube guidance. Annu. Rev. Plant Biol. 66: 393–413.
- Intergovernmental Panel on Climate Change (IPCC). 2022. Climate change 2022: Impacts, Adaptation, and Vulnerability. In: H.-O. Pörtner, D. C. Roberts, M. Tignor, E. S. Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V. Möller, A. Okem and B. Rama (eds.). Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge and New York.
- Iwahori, S. 1965. High temperature injuries in tomato. IV. Development of normal flower buds and morphological abnormalities of flower buds treated with high temperature. J. Japan. Soc. Hort. Sci. 34: 33–41.
- Katano, K., T. Oi and N. Suzuki. 2020. Failure of pollen attachment to the stigma triggers elongation of stigmatic papillae in Arabidopsis thaliana. Front. Plant Sci. 11: 989. DOI: 10.3389/fpls.2020.00989.
- Kawamoto, T. 2003. Use of a new adhesive film for the preparation of multi-porpose fresh-frozen sections from hard tissues, whole-animals, insects and plants. Archives Histol. Cytol. 66: 123–143.
- Kumazawa, S., T. Ohara and K. Niiuchi. 1954. The differentiation of varieties of peppers in Japan. J. Japan. Soc. Hort. Sci. 23: 152–158 (In Japanese with English abstract).
- Levy, A., H. D. Rabinowitch and N. Kedar. 1978. Morphological and physiological characters affecting flower drop and fruit set of tomatoes at high temperatures. Euphytica 27: 211–218.
- Luo, Y. and A. Widmer. 2013. Herkogamy and its effects on mating patterns in Arabidopsis thaliana. PLoS ONE 8: e57902. DOI: 10.1371/journal.pone.0057902.
- Mesihovic, A., R. Iannacone, N. Firon and S. Fragkostefanakis. 2016. Heat stress regimes for the investigation of pollen thermotolerance in crop plants. Plant Reprod. 29: 93–105.
- Opedal, O. H. 2018. Herkogamy, a principal functional trait of plant reproductive biology. Int. J. Plant Sci. 179: 677–687.
- Peet, M. M., S. Sato and R. G. Gardner. 1998. Comparing heat stress effects on male-fertile and male-sterile tomatoes. Plant Cell Environ. 21: 225–231.
- Richards, A. J. 1986. Plant Breeding Systems. 529 pp. George Allen & Unwin, London, U.K.
- Sato, S., M. Kamiyama, T. Iwata, N. Makita, H. Furukawa and H. Ikeda. 2006. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 97: 731–738.
- Sato, S., M. M. Peet and J. F. Thomas. 2000. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 23: 719–726.
- Smith, O. 1932. Relation of temperature to anthesis and blossom drop of the tomato together with a histological study of the pistils. J. Agr. Res. 44: 183–190.
- Smith, S. H. 2015. In the shadow of a pepper-centric historiography: Understanding the global diffusion of capsicums in the sixteenth and seventeenth centuries. J. Ethnopharmacol. 167: 64–77.
- Torang, P., L. Vikstrom, J. Wunder, S. Wotzel, G. Coupland and J. Agren. 2017. Evolution of the selfing syndrome: anther orientation and herkogamy together determine reproductive assurance in a self-compatible plant. Evolution. 71: 2206–2218.
- Xu, J., M. Wolters-Arts, C. Mariani, H. Huber and I. Rieu. 2017. Heat stress affects vegetative and reproductive performance and trait correlations in tomato (Solanum lycopersicum). Euphytica 213: 156. DOI: 10.1093/jxb/erq053.
- Yamazaki, A. and M. Hosokawa. 2019. Increased percentage of fruit set of F1 hybrid of Capsicum chinense during high-temperature period. Sci. Hortic. 243: 421–427.
- Yamazaki, A. and M. Hosokawa. 2020. Relationship between fruit-set ability of an F1 hybrid of Capsicum chinense and the pollen germination rate under high temperature condition. Hort. Res. (Japan) 19 (Suppl. 1): 159 (In Japanese).
- Yamazaki, A., A. Takezawa, K. Nagasaka, K. Motoki, K. Nishimura, R. Nakano and T. Nakazaki. 2022a. Development of simple method for pollen germination rate measurement by machine learning. Hort. Res. (Japan) 21 (Suppl. 2): 330 (In Japanese).
- Yamazaki, A., A. Takezawa, R. Nakano, K. Nishimura, K. Motoki, M. Hosokawa and T. Nakazaki. 2022b. Indicator candidate traits for autonomous fruit set ability under high temperatures in Capsicum. J. Hortic. Res. 30: 105–116.
- Yashiro, K., Y. Sakai and H. Namai. 1999. Relationships between pollen–ovule ratio and autofertility, self-compatibility, automatic self-pollination ability in heterogeneous incomplete autogamous plants, Thai mustard. Breed. Sci. 49: 39–42.