Introduction
Ocular ischemic neovascular diseases currently constitute the most common causes of
moderate and severe visual loss.1 These
diseases can be broadly divided into two categories: retinal neovascular diseases and
subretinal neovascular diseases.1 Retinal
neovascular diseases involve neovascularization from retinal vessels, whereas subretinal
neovascular diseases are associated with subretinal neovascularization that occurs in the
avascular outer retina and subretinal space. Retinal neovascular diseases generally include
diabetic retinopathy2 and retinopathy of
prematurity,3 and subretinal
neovascular diseases include age-related macular degeneration.4
To date, treatments for these diseases consist of laser photocoagulation, topical injection
of corticosteroids, intravitreal injection of anti-vascular endothelial growth factor
(anti-VEGF) agents and vitreoretinal surgery.5 Although VEGF inhibition strategies have proven to be effective in some
clinical studies, it is evident that other pro-angiogenic factors play an important role in
these diseases, as reported in relation to cases in which anti-VEGF therapies have
failed.6,7 Briefly, one report demonstrated that
SDF-1/CXCR4 may contribute to the involvement of bone marrow-derived cells that work with
VEGF to promote the development of neovascularization in the eye in cases of both retinal
and subretinal neovascularization.8 Another
report indicated that paracrine collaboration of IGF-1 and VEGF may contribute to the
development of choroidal neovascularization.9 Angiopoietins and their interactions with VEGF or other angiogenic factors
also have been postulated to regulate pathological vascular development, maintenance and
permeability in the eye.10,11,12 In
addition to angiogenic factors, several reports have suggested that increased monocyte
chemoattractant-1 (MCP-1/CCL2) levels in the eye may enhance diabetic retinopathy13; indeed, upregulation of CCL2 has been
consistently reported in the vitreous fluid of patients with diabetic retinopathy and
ischemic retinopathies.13,14,15,16,17,18 Finally,
the long-term use of anti-VEGF therapies has the possibility of significant adverse effects
regarding blockade of crucial physiological roles of VEGF in the eye.19,20 Therefore, new types of effective treatments are
desirable.
Hypoxia-inducible factor (HIF) is a master regulator of hypoxia-responsive genes that act
against hypoxia.21 Hypoxia-responsive
genes are involved in angiogenesis, erythropoiesis, energy metabolism and
inflammation.22,23 Therefore, HIF can induce pathological
pro-angiogenic gene expressions (including that of VEGF-A) under retinal hypoxia, ultimately
leading to the development of ocular ischemic neovascular diseases.24,25 Herein, we describe the inhibition of HIF as a promising
therapeutic strategy for ocular diseases and summarize promising recently investigated HIF
inhibitors as treatments for ocular ischemic neovascular diseases.
Overall Roles of HIF
Many organisms use oxygen to produce ATP as an energy source to sustain themselves. The
majority of all living things, including humans, cannot maintain their metabolism without
oxygen. Organisms are sometimes exposed to transient or chronic hypoxia resulting from
environmental factors or diseases. As a response to such factors and diseases, under hypoxic
conditions, HIF exerts an important role in adaptive regulation of energy metabolism by
triggering the switch from mitochondrial oxidative phosphorylation to anaerobic
glycolysis.26
HIF is a basic helix–loop–helix DNA binding protein of the Per-ARNT-Sim family
(bHLH-PAS).27 HIF was discovered by
Semenza et al. in 1992 as a hypoxia-dependent inducer of erythropoietin in a liver cancer
cell line (Hep3B).28 In 1995, HIF-1 was
reported to be a heterodimer of HIF-1α and HIF-1β/ARNT (an aryl hydrocarbon receptor nuclear
translocator).23,29,30,31 Subsequently, HIF-2α/EPAS2 and HIF-3α were identified.28,32 Among the HIF-α subunits, HIF-1α and HIF-2α have an N-terminal
trans-activation domain (N-TAD) and a C-terminal transactivation domain (C-TAD) on the
C-terminal side, which is involved in transcriptional activation (Fig. 1). In contrast, HIF-3α does not possess C-TAD (Fig. 1). Multiple variants of HIF-3α have been
characterized.33,34 In mouse variants, neonatal embryonic PAS and
inhibitory PAS domain (IPAS) proteins have been identified.35 Moreover, there is a large number of human variants of
HIF-3α.35 HIF-3α has relatively weak
transcriptional activity because it does not include C-TAD. IPAS behaves as a dominant
negative form of HIF.34,36,37 Each
HIF-α subunit contains an oxygen-dependent degradation domain that regulates the stability
of HIF-α.33,34 In contrast to HIF-α family members, HIF-1β is
stably expressed in cells and allows HIF-α family members to bind to DNA and stimulate
transcription of their target genes.27

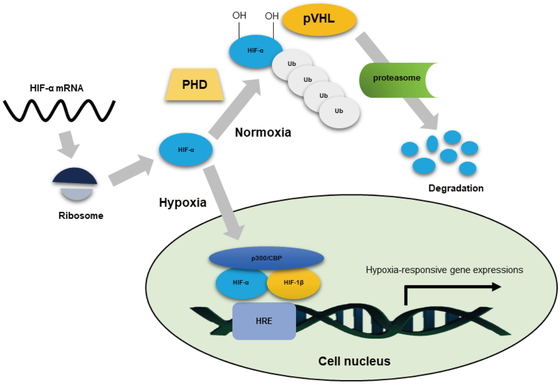
Under normoxia, HIF-α is hydroxylated by prolyl hydroxylases (PHD) (Fig. 2). Prolylhydroxylated HIF-α is ubiquitinated by E3 ubiquitin
ligase complex (composed of pVHL, Elongin B/C, Cul2 and Rbx1), resulting in the rapid
proteasome-dependent degradation of HIF-α (Fig.
2). In contrast, under hypoxic conditions, the activity of PHD is reduced and HIF-α
is stabilized (Fig. 2). HIF-α then translocates
into the nucleus to form heterodimers with HIF-1β that then bind to the hypoxia-response
element, which is located upstream of HIF target genes (Fig. 2). Furthermore, the C-TAD region is important for optimal transcriptional
activity of HIF. However, HIF can be induced to a lesser extent without binding to the
CREB-binding protein (CBP)/p300. Therefore, recruitment of CBP/p300 enhances HIF-mediated
transcriptional activity (Fig. 1 and Fig. 2). Despite the constant biosynthesis of HIF-α,
under normal oxygen conditions, the transcriptional activity of HIF is kept very low by the
mechanism, i.e., the oxygen-dependent hydroxylation of asparagine by the asparaginyl
hydroxylase factor inhibiting HIF-1 (FIH-1) inhibits the interaction of HIF with
CBP/p300.38
HIF-mediated cellular responses to low levels of oxygen fall into two categories: short-
and long-term adaptation. For short-term adaptation, the oxygen supply can be rapidly
increased by HIF activation through upregulation of the vasodilatory enzyme iNOS (inducible
nitric oxide synthase).39 The enzymatic
product of iNOS, nitric oxide, induces a fast increase in blood flow via the relaxation of
vascular smooth muscle cells.40 Oxygen
homeostasis can also be modulated by an increase in the glycolysis machinery through
induction of glycolytic enzymes such as glucose transporter-1 (GLUT1)/SLC2A141,42 and suppression of mitochondrial respiration through induction
of pyruvate dehydrogenase kinase-1 (PDK1).43,44 GLUT1 and
PDK1 are well known HIF target genes. When it comes to long-term adaptation, a series of
hypoxia-stimulated angiogenesis processes are initiated through induction of pro-angiogenic
genes such as VEGF-A, angiopoietin-1, angiopoietin-2, platelet-derived growth factor, basic
fibroblast growth factor and monocyte chemoattractant protein-1 (MCP-1/CCL2), which finally
induces endothelial cell proliferation, sprouting, migration, adhesion and tube formation as
well as vascular permeabilization.45,46 Because
these factors are regulated by HIF, HIF can be regarded as the master regulator of
angiogenesis. This process would appear to resolve the matter of chronic low oxygen levels
in tissues. However, numerous pathological diseases related to hypoxia have a general basis
in this process.47,48,49,50,51
Pathological ocular diseases related to hypoxia include diabetic retinopathy (DR),
age-related macular degeneration (AMD), glaucoma and retinopathy of prematurity (ROP); these
conditions are discussed separately below.
HIF Inhibition in Ocular Diseases
Age-related macular degeneration
AMD occurs when abnormal new blood vessels develop under the retina, a process called
choroidal neovascularization (CNV). AMD leads to severe visual loss in the elderly
population. There remains neither a strongly effective preventive treatment nor a cure for
AMD.52,53 Although the development of AMD is
associated with complex pathological mechanisms, increasing evidence suggests that
dysfunction of retinal pigment epithelium (RPE) may be mainly involved in neovascular and
atrophic forms of AMD.54 RPE is a
monolayer of polarized cells that resides below the retina and interacts with
photoreceptor cells to achieve retinal homeostasis.55 The outer blood–retina barrier can be modulated by RPE via regulation
of the delivery of nutrients and oxygen to the outer retina and the clearance of metabolic
waste from the outer retina. RPE produces growth factors such as VEGF to support the
physiologies of the retina and choriocapillaris.54,56,57
However, aging and increased oxidative stress can result in dysfunction of RPE, ultimately
leading to RPE producing pathological amounts of growth factors, especially
VEGF.58,59 This process may be a pathological cause of
the development of CNV, which finally leads to AMD.60 To investigate this process, we attempted to control RPE via
inhibition of the HIF/VEGF-A axis in RPE. We found a significant reduction of the CNV
volume in a RPE-specific Hif-1α conditional knockout mouse model of CNV
compared with that of the CNV volume in the age-matched control mouse model of
CNV.61 Furthermore, by carrying out
extensive screenings of drugs and food ingredients, we discovered several HIF inhibitors
(lactoferrin, rice bran, vitamin B6, Garcinia cambogia and hydroxycitric
acid) and demonstrated that diet supplemented with these ingredients can reduce the CNV
volume in a mouse model of CNV and reduce VEGFA mRNA expression under
CoCl2-induced hypoxia.61,62,63 Lee et
al. investigated the effects of resveratrol on HIF-1α and VEGF expressions in human
ARPE-19 cells and a mouse model of CNV.64 Resveratrol, a natural compound found in grapes, has been suggested to
suppress VEGF in various cancer cells, possibly through HIF-1α inhibition.65,66,67 In ARPE-19 cells, resveratrol significantly reduced HIF-1α and VEGF-A
expressions by blocking the PI3K/Akt/mTOR signaling pathway and promoting proteasomal
HIF-1α degradation; moreover, oral administration of resveratrol reduced the CNV volume in
a mouse model of CNV.64 Song et al.
found that chrysin (5,7-dihydroxyflavone), another flavonoid naturally present in many
fruits and vegetables, inhibited HIF-1α and VEGF expressions with an accompanying
reduction of the CNV volume in a rat model of CNV.68 Using a non-pharmacological approach, Koo et al. attempted to use
therapeutic gene editing in a mouse model of CNV.69 The intravitreal delivery of CRISPR RNA-guided endonuclease LbCpf1
(derived from Lachnospiraceae bacterium ND2006) targeted to
Hif-1α or Vegfa reduced the volume of CNV as
efficiently as aflibercept (an anti-VEGF drugs currently in clinical use).69 Furthermore, LbCpf1 targeted to
Hif-1α or Vegfa exerted long-term therapeutic effects,
unlike aflibercept.69 In summary, even
though the mode of action in vivo requires further investigation, control of the
HIF/VEGF-A axis in the eye could have therapeutic value in the management of CNV and
ultimately suppress the development of AMD.
Retinopathy of prematurity
ROP is an ocular disease of prematurely born babies, usually seen in those having
received neonatal intensive care in which oxygen therapy is administered because of
insufficient lung development.70 In
contrast to the damaging vessels found in AMD, retinal blood vessels grow disorganized in
ROP and result in scarring and retinal detachment, which may lead to blindness in serious
cases.70 The development of retinal
neovascularization also involves complex pathological mechanisms similar to that of CNV.
The involvement of multiple interlinked structural and functional changes caused by
abnormal crosstalk between retinal neurons, glial cells and vasculatures has been
suggested as the mechanism of ROP development.71 Neurons (with glial cells) act as sensors of oxygen and regulators of
vasculatures by interacting with various types of vessels in the central nervous
system,72 and impairment of their
crosstalk results in uncontrolled neurovascular homeostasis.72,73 In the eye (a component of the central nervous system), the activation
of HIFs in neurons (especially HIF-1α) and glia cells (especially HIF-2α) are suggested to
have critical pathological roles in retinal vascular diseases.74 We demonstrated that no change in Hif-2α
mRNA expression and only a slight increase in HIF-2α protein expression was seen in the
ischemic retina, whereas expressions of Hif-1α mRNA and HIF-1α protein
showed significant increases along with upregulation of Vegf mRNA
expression in a mouse model of oxygen-induced retinopathy (OIR), a condition that mimics
ROP.75,76 Furthermore, we demonstrated that
Hif-1α and Vegfa in retinal neuronal cells were
responsive to CoCl2-induced hypoxic stress, whereas Hif-2α was
not induced.77 This finding suggests
that the HIF-1α/VEGF-A axis in retinal neurons may be a major regulator of retinal
neovascularization. In contrast, Weidemann et al. demonstrated that astrocyte-derived VEGF
may be essential for retinal neovascularization in a model of OIR,78 and Korovina et al. suggested that HIF-2α in
hematopoietic cells may contribute to the development of retinal neovascularization in a
model of OIR.79 Against this background,
for a better understanding of the pathological mechanisms of retinal neovascularization,
comprehensive studies need to be continued regarding the effects of the main subtypes of
HIF on the development of retinal neovascularization as well as investigations of
VEGF-producing cell types and systemic vascular cell conditions.
As a pharmacological treatment for ROP, we suggested topotecan, a topoisomerase I
inhibitor that decreased HIF-1α stabilization in a mouse model of OIR with a reduction in
upregulated Vegfa and Angpt2 mRNA expressions in the
ischemic retina, which ultimately prevented retinal neovascularization.75,76 Doxorubicin (a widely used HIF-1α inhibitor) also prevented
retinal neovascularization in a mouse model of OIR, even though the mode of action of
doxorubicin was not directly stated in the study.75,76 We
also attempted to use marine products to prevent retinal neovascularization in a mouse
model of OIR.80 Among various materials
from marine fish, Decapterus tabl ingredients were shown to possess HIF
inhibitory effects under dimethyloxalylglycine-induced hypoxic conditions.80 Furthermore, oral administration of
Decapterus tabl ingredients suppressed retinal neovascularization in a
mouse model of OIR even though the mode of action in vivo and the effector compounds in
the ingredients have not been clarified.80
In addition to marine products, we focused on investigating novel HIF inhibitors from
mushroom products and found that 2-azahypoxanthine (AHX), a fairy chemical from the fungus
Lepista sordida,81,82 had an
inhibitory effect on HIF activation in retinal cells and suppressed Vegfa
mRNA upregulation under CoCl2-induced hypoxic conditions.77 Oral administration of AHX significantly
suppressed retinal neovascularization in a mouse model of OIR.77 Yoshida et al. applied digoxin (a cardiac glycoside) to
suppress retinal neovascularization by inhibiting HIF activation83 based on the suggestion that digoxin blocks tumor growth via
HIF-1α synthesis inhibition.84 In the
ischemic retinas in a model of OIR, digoxin selectively inhibited expressions of HIF
target genes such as Bnip3, Glut1 and
Epo and also inhibited expressions of hypoxia-induced angiogenic growth
factors and their cognate receptors such as Vegfa, Sdf-1
(stromal cell-derived factor 1), Scf (stem-cell factor),
Pdgf-b (platelet-derived growth factor subunit B),
Pgf (placental growth factor), Vegfr2 (vascular
endothelial growth factor receptor 2), Cxcr4 (C-X-C chemokine receptor
type 4), Cd117 and Pdgfr-b (platelet-derived growth
factor receptor subunit B).83 Usui-Ouchi
et al. designed an allosteric peptide inhibitor of HIFs to examine whether HIF inhibition
peptide therapy is applicable to counteract retinal neovascularization.85 The peptide design concept was the
disruption of HIF binding to coactivators CBP/p300 to reduce pathological HIF-mediated
hypoxia-responsive gene expressions as well as HIF-related pro-inflammatory gene
expressions. The peptide inhibitor of HIFs reduced the upregulated expressions of
Vegfa, Epo, Tnf-α,
Ccl2, Ccl3 and Il-1β in ischemic
retinas and effectively regulated retinal neovascularization in a mouse model of
OIR.85 Furthermore, a combination of
this peptide inhibitor and a reduced concentration of the anti-VEGF agent aflibercept
showed promising therapeutic effects on the regulation of retinal
neovascularization.85
By taking a different approach from the direct application of HIF inhibitors, we
demonstrated that pemafibrate could prevent retinal neovascularization via upregulation of
serum levels of fibroblast growth factor 21 (FGF21), a possible HIF inhibitor, in a mouse
model of OIR.75 Pemafibrate is a
selective peroxisome proliferator-activated receptor α modulator that was recently
approved in Japan as a powerful therapeutic agent against hyperlipidemia. Pemafibrate
activated PPARα in the liver, resulting in an increase in serum levels of FGF21.86,87 Previously, Fu et al. showed that FGF21 administration could
suppress pathological retinal neovascularization in a mouse model of OIR.88 Following this, we found a significant
increase in serum levels of FGF21 by pemafibrate administration in a model of
OIR.75 Moreover, we suggested that a
decrease in retinal HIF-1α immunoreactivities along with a reduction in
Vegf mRNA expression in a mouse model of OIR may have a strong
correlation with FGF21 induction by pemafibrate.75 Chen et al. examined whether retinal neovascularization could be
modulated by treatment with fenofibrate, another PPARα agonist.89 Fenofibrate also ameliorated retinal neovascularization in a
rat model of OIR and inhibited tube formation and migration in endothelial
cells.89 The effects of fenofibrate
can be partially explained by the blockade of activation of HIF-1α and upregulation of
VEGF.89 Taken together, these
reports suggest that the control of HIF-1α, VEGF and other pathological angiogenic factors
could have a great impact on the regulation of retinal neovascularization.
Diabetic retinopathy
DR is one of the most serious and common complications of diabetes in subjects with type
1 or type 2 diabetes mellitus.90 It is a
chronic microvascular complication in the retina caused by chronically uncontrolled
glucose levels in the body during the development of diabetes.91,92 The chronic microvascular damage leads to severe hypoxic conditions and
results in retinal neovascularization.91,92 Chronic
severe hypoxic stress in the retina can induce retinal detachment, which may result in
loss of vision.92 Rodrigues et al.
showed that HIF-1α and VEGF-A were expressed in the retinas of proliferative sickle
retinopathy patients, higher than that in the retinas of controls. This finding implies
that HIF-1α might be important for the development of DR.93 Wang et al. determined that the levels of VEGF-A and HIF-1α
in the vitreous fluid of patients with proliferative DR were higher than those in control
subjects.94 This also implies that
VEGF-A and HIF-1α could be associated with the pathological angiogenic processes of
DR.94 Yamada et al. focused on
genetic variation in HIF-1α by examining all regions of the gene for
single-nucleotide polymorphisms in type 2 diabetes patients and performed association
studies in Japanese type 2 diabetes patients with or without retinopathy.95 Mutated HIF-1α (P582S) was found to be
associated with type 2 diabetes and had a higher transcriptional activity than the
wild-type, which implies that HIF-1α may have a strong association with the occurrence of
type 2 diabetes.95 Even though no
association with retinopathy was detected with the P582S HIF-1α mutation, Yamada et al.
suggested that P582S HIF-1α should be assessed in larger studies to confirm this
finding.95 Ekberg et al. suggested
that the P582S HIF-1α mutation may have a protective effect against the development of
severe DR with a strong risk reduction, even when adjusting for known risk factors for DR
such as diabetes duration, hyperglycemia and hypertension.96 This finding implies that HIF-1α may be important for the
development of severe DR. In summary, the potential roles of HIF-1α in the development of
DR have been much discussed in clinical studies.
Preclinical animal studies of DR are problematic because the currently available
experimental models do not clearly and exactly recapitulate the entire pathophysiology of
changes in neurons and vasculatures that occur at each stage of human DR.97,98 For example, the streptozotocin-induced diabetic model (one
of the most widely used experimental models for diabetes) has a limited ability to mimic
proliferative DR, which means that neovascularization is not clearly seen in the
model.99,100,101,102,103
The galactose feeding approach to diabetic modeling requires a long period to develop DR
in mammals,104,105,106 and proliferative DR is still rarely formed in the model
using this approach. Pancreatectomy, which directly removes the pancreas, is only
applicable to large animals such as cats or monkeys, but these have large variations among
individuals.107,108
The direct injection of VEGF to the eyes to induce neovascularization is not mediated by
hyperglycemia and, after VEGF is metabolized physiologically, the effects of VEGF could
easily disappear.109 A mouse model of
OIR (also used for studying ROP) has been classically used because neovascularization is
highly inducible and applicable to rodents without technical difficulties.110 However, this method is also not DR
specific, and only newborn rodents are available for induction of retinal
neovascularization. The development of clinically relevant murine models, e.g., combining
ischemic experimental models with streptozotocin injection111 or high-fat diet supplementation112 to increase glucose levels, must first be established to
better understand how HIF modulation has protective (or deleterious) effects in the
development of DR.
Glaucoma
Glaucoma is a group of eye conditions that results in damage to the optic nerve, which
leads to vision loss. This disease is characterized by high intraocular
pressure.113 Elevated eye pressure
can be induced by overproduction of the aqueous humor that flows throughout the eye or by
dysfunction of the drainage system of the aqueous humor through the trabecular meshwork at
the angle where the iris and cornea meet.113 Even though the occurrence of neovascular glaucoma is lower than that
of other types of glaucoma, neovascular glaucoma is known as a potentially blinding
secondary glaucoma114 characterized by
the development of new vessels over the iris that obstruct outflow of the aqueous humor
and thereby increase intraocular pressure.115 Interestingly, elevated intraocular pressure was suggested to increase
HIF-1α expression in the retina in an experimental model of glaucoma116; furthermore, there was an increase in
immunoreactivities of HIF-1α in the retina and optic nerve head of glaucomatous human eyes
compared with those of control eyes.117
Consequently, we investigated whether HIF-1α inhibition has protective effects on the
retina in a mouse model of glaucoma.118
Using the general HIF-1α inhibitor topotecan, we demonstrated that impairment of visual
function in a mouse model of glaucoma could be suppressed by topotecan administration via
reduction in stabilized HIF-1α protein levels in the retina.118 Furthermore, we found that another HIF inhibitor,
halofuginone (a synthetic derivative of febrifugine), could prevent neurodegeneration in a
mouse model of glaucoma via inhibition of HIF-1α activation in the retina.119 Compared with the involvement of HIF in
AMD and/or ROP, the effects of HIF modulation in glaucoma have been less studied.
Therefore, more studies are needed to better understand how HIF inhibition could have
protective effects in glaucomatous retinas.
Corneal diseases
In addition to the retina and choroid, corneal neovascularization is also a
sight-threatening condition caused by new vessel formation from the limbus and its
invasion into the cornea, resulting in ocular inflammation, infection and
hypoxia.120,121 HIF-1α expression has been detected in
corneal neovascularization, and VEGF-A, also known to be a main pro-angiogenic factor in
corneal neovascularization, is under regulation of the HIF-1α signaling pathway in the
cornea.122,123 Therefore, HIF inhibition has been
suggested as a promising target for treating corneal neovascularization. Chen et al. used
shRNA targeting HIF-1α to suppress VEGF-A expression and corneal neovascularization in a
mouse model of closed-eye contact lens wear.122 Fu and Xin demonstrated that silencing HIF-1α or silencing VEGF-A
inhibited corneal neovascularization, while double-target interference of VEGF and HIF-1α
improved pathological changes regarding corneal neovascularization to a greater
extent.124 Kwon et al. examined
whether rapamycin (an effective antifungal agent125) could inhibit corneal neovascularization in an alkaline-burned
corneal mouse model.126 As a result,
mRNA expressions of substance P and Hif-1α were
inhibited by rapamycin treatment (1–3 days) with a reduction in VEGF-A expression on day 7
in corneal tissues.126 This implies
that rapamycin could be useful as an angiogenic regulator in the treatment of corneal
neovascularization. Zhang et al. focused on identifying the profile of miRNA expressions
in neovascularized corneas after alkali burn injuries to the eye and found that miR-21 in
alkali-burned corneas showed the greatest increase.127 Using subconjunctival injection of antagomir-21, an inactivator of
miR-21, progression of neovascularization was attenuated by inhibiting expressions of
VEGF-A and HIF-1α.127 Taken together,
this evidence suggests that targeting HIF-1α could also be useful for the management of
corneal neovascularization.

