Abstract
Our understanding of the biology of the intestinal epithelium has advanced since the
establishment of an organoid culture system. Although organoids have enabled investigation
of the mechanism of self-renewal of human intestinal stem cells in vitro,
it remains difficult to clarify the behavior of human normal and diseased intestinal
epithelium in vivo. Recently, we developed a xenotransplantation system
in which human intestinal organoids are engrafted onto epithelium-depleted mouse colons.
This xenograft recapitulated the original tissue structures. Upon xenotransplantation,
normal colon organoids developed normal colon crypt structures without tumorigenesis,
whereas tumor-derived organoids formed colonic tumors resembling the original tumors. The
non-tumorigenicity of human intestinal organoids highlights the safety of organoid-based
regenerative medicine. As an example of regenerative medicine for short bowel syndrome, we
devised a unique organ-repurposing approach to convert colons into small intestines by
organoid transplantation. In this approach, the transplanted rat small intestinal
organoids not only engrafted onto the rat colons but also remodeled the colon
subepithelial structures into a small intestine-like conformation. Luminal flow
accelerated the maturation of villi in the small intestine, which promoted the formation
of a lymphovascular network mimicking lacteals. In this review, we provide an overview of
recent advances in gastrointestinal organoid transplantation and share our understanding
of human disease biology and regenerative medicine derived from these studies.
Background
The intestinal epithelium is one of the most rapidly renewing tissues, and this homeostasis
is strictly controlled by the intestinal stem cells located at the bottom of the
crypts.1 Intestinal stem cells are
defined by their capacity for multipotent differentiation and long-term self-renewal.
Previously, long-term culture from primary adult tissue stem cells was not possible, which
limited intestinal stem cell research to experiments in mice and cell lines. In 2007,
in vivo genetic lineage tracing experiments using a genetically
engineered mouse model demonstrated that the crypt base columnar cells, marked by the
expression of LGR5, serve as intestinal stem cells.1 When Cre recombinase was activated by the
administration of tamoxifen, LGR5-LacZ-positive cell-derived LacZ+ blue clones
emanated from the crypt bottoms and later extended up the sides of the crypts. Owing to the
discovery of LGR5, researchers have obtained a better understanding of the
regulatory niche signals that cooperatively direct intestinal stem cells to perform their
functions properly. By recapitulating the in vivo extracellular niche
microenvironments of intestinal stem cells, Sato et al. established an in
vitro three-dimensional (3D) culture system known as organoids, which allowed the
long-term expansion of adult intestinal epithelium.2 After being established as a culture method for the mouse small
intestine, this technique paved the way for establishing culture systems for human
gastrointestinal tissues3,4,5,6 and
various other organs.7,8,9 With the
advent of organoids, the study of intestinal stem cells has made tremendous progress over
the past 15 years.10 Organoids are useful
not only as an in vitro research tool, but also as an in
vivo research tool for studying human intestinal epithelium, and have potential
as a cell source for transplantation. In this review, we highlight in vivo
studies on intestinal organoids, including our own work.
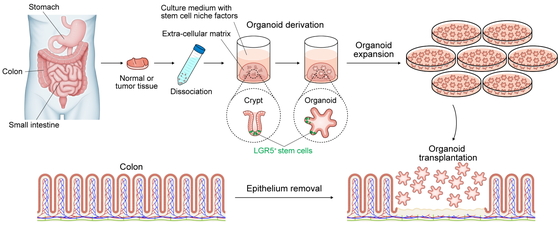
Intestinal Organoids
The organoid culture system reconstructs a niche; that is, a microenvironment essential for
the maintenance of stem cell function, including growth factors and extracellular matrix
that support the proliferation of tissue-derived stem cells ex vivo. For
mouse small intestinal organoid culture, epidermal growth factor (EGF), a bone morphogenetic
protein inhibitor (noggin), and a Wnt activator (R-spondin1) are added to the culture medium
and embedded in Matrigel extracellular matrix.2 Although the composition of the stem cell niche varies by organ and
species, the addition of Wnt-3A and a transforming growth factor-β inhibitor to the above
conditions enabled the culture of mouse colon, and further addition of a p38
mitogen-activated protein kinase inhibitor enabled the culture of human intestinal
organoids.3,11 However, these organoids have had difficulty
maintaining a differentiated cell type because of their strong dependence on growth factors
that inhibit differentiation. Although p38 inhibitor is required for long-term maintenance
of human intestinal organoids, its removal is essential for the reproduction of secretory
cells in vitro. Screening for biologically relevant niche factors produced
by human intestinal subepithelial stromal cells revealed that the combination of
insulin-like growth factor-1 and fibroblast growth factor-2 greatly promotes organoid growth
and represents a niche factor for human intestinal stem cells that could replace EGF and a
p38 inhibitor.6 Single-cell RNA sequencing
revealed that intestinal epithelia cultured under these refined organoid culture conditions
maintain a pattern of gene expression that is highly homologous to that of in
vivo tissues. These conditions also promoted the growth of p38
inhibitor-sensitive colon tumor organoids, including patient-derived ulcerative colitis
(UC)-associated dysplastic organoids, and promoted the efficiency of CRISPR–Cas9-mediated
genome editing.6,12
Normal Colonic Organoid Transplantation
The organoid culture method of normal intestinal stem cells is associated with a low risk
of tumorigenesis because the cultivation of tissue stem cells occurs by simply taking
intestinal tissue as the original source and adding factors. Unlike other organs that are
difficult to access for tissue collection, the intestine has the unique advantage of being
easily accessible using advanced endoscopic technology. Therefore, the application of
patient-derived somatic epithelial stem cells in regenerative medicine, in which they are
grown in vitro and used as cells for transplantation, has been eagerly
anticipated. In 2012, Yui et al. reported the successful transplantation (via a transanal
approach) and engraftment of colonic organoids (from enhanced green fluorescent protein
transgenic mice) into ulcerated areas of the colon of Rag2-knockout mice with dextran
sulfate sodium salt (DSS)-induced colitis.13 This success stimulated research in Japan towards a first-in-human trial
of organoid transplantation for patients with UC. Subsequently, several groups successfully
transplanted various mouse normal organoids into mice and successfully used collagen instead
of Matrigel as extracellular matrix.14,15,16,17,18,19,20,21
Despite these studies in mice, no method had been developed to reconstruct human intestinal
epithelium in vivo by transplanting normal human intestinal epithelium.
Tumor cells grow subcutaneously and under the renal capsule in immunodeficient
mice,4,22 but human normal intestinal cells cannot grow
at such sites where niche factors that support epithelial growth are lacking. Therefore, the
only way to study the human intestinal epithelium was to study it in humans themselves, but
the administration of various drugs and gene editing for analysis in this context naturally
involve ethical difficulties. Most study results were interpreted on the assumption that
human intestinal epithelium would probably be similar to mouse epithelium. However, although
the stem cell marker LGR5 was identified in mice,1 it has not been rigorously demonstrated whether
LGR5 is a stem cell marker in humans. To address this issue, we sought to
establish a method for xenotransplantation of human normal colonic organoids using
immunodeficient mice. Because natural killer (NK) cell activity plays an important role in
the rejection of xenotransplanted cells,
NOD.Cg-PrkdcscidIl2rgtm1Sug/ShiJic (NOG) mice
with multiple immune functional defects, including dendritic cells and the absence of T, B,
and NK cells, were used as human colonic organoid recipients.23 After trying various epithelial abrasion methods, we decided
to use ethylenediaminetetraacetic acid (EDTA), which has been proven to be optimal for
epithelium removal.2,15 Importantly, the crypts of the colonic
epithelium of the recipient mice must be completely removed, including the crypt base where
the stem cells reside, or they will be repaired by the original epithelium, and the
transplanted cells will fail to engraft. Using EDTA-based breaching with minimal damage to
nonepithelial tissue, we were able to establish a technique for removing the crypts and
replacing them with transplanted organoids.24,25 Organoid
xenografts that have been fluorescently labeled by the piggyBac transposon system or
lentivirus-based gene transfer can be observed endoscopically over time. When human colonic
organoids were transplanted into the mouse colon using this technique, the structure
reconstructed in the mouse colon was that of the human colonic crypt, which is clearly
larger than the mouse crypt structure and has different mucus and other properties. This
means that the stem cell intrinsic program of transplanted cells is preserved in different
environments. Transplanted human colonic epithelial cells were found to grow tumor-free in
the mouse colon for more than 10 months, providing the first significant data towards
clinical application (Fig. 1), and showing that
transplanted human intestinal organoids are not tumorigenic in
vivo.24
The establishment of this transplantation system allowed us to conduct experiments with
normal intestinal epithelium, which could not be performed with human cells because of the
lack of an adequate experimental system. First, we aimed to reveal the stem cell nature of
LGR5-positive cells by demonstrating their multipotency and self-renewal
capacities through genetic lineage tracing experiments. Gene editing cannot be performed
in vivo on the human intestine but can be performed in
vitro on human intestinal organoids. The system established in colorectal cancer
organoids using CRISPR–Cas9 gene editing, in which LGR5-positive cell-derived cells are
differentially fluorescently labeled by tamoxifen induction,22 was introduced into human normal colon organoids.24 The organoids were then xenotransplanted into
the colon of NOG mice, resulting in mice with a human cell lineage, thereby enabling
in vivo genetic lineage analysis in the human intestinal epithelium.
Notably, we also found that human and mouse intestinal epithelial stem cells proliferate at
different rates,24,26 suggesting the importance of conducting
studies in human cells. Differences in cell composition and proliferation speed naturally
suggest that drug reactivity and toxicity may also differ between humans and mice. Although
much knowledge can be obtained from experiments with mice, studies using human cells are
extremely important.27,28
Unexpectedly, removal of the epithelium also led to new insights into tissue regeneration.
The injured area of the epithelium is repaired by the surrounding epithelial cells.
Therefore, the injured area can be repaired by the transplanted organoids or adjacent
epithelial cells, which can replace the epithelium even if the cells differ from the
original cells. Colonic mucosal injuries created contiguously to the anus were repaired by
adjacent anally derived squamous epithelium.29,30 This
mechanism may be one explanation for the clonal field expansion of cells with
inflammation-resistant somatic gene mutations and colitis-associated dysplasia in patients
with UC characterized by chronic recurrent epithelial injury.12,31,32
Tumor Organoid Transplantation
Unlike normal intestinal epithelial cell transplantation, much experience has been
accumulated with tumor cell transplantation using cell lines and organoids, including
subcutaneous transplantation into nude mice and subrenal capsular transplantation in NOG
mice.4,22,33 Although orthotopic transplantation of mouse or human tumor
organoids by colonoscopy-based, surgery-based, or other local injection into the submucosal
layer or intestinal wall has been actively performed,34,35,36,37,38 these
techniques result in tumor growth from the serosal side rather than from the luminal side,
which differs from the original growth and invasion of the tumor. Moreover , using this
approach, low-grade tumors would be difficult to engraft. To resolve these problems,
diseased cells need to be transplanted from the mucosal side into injured regions caused by
the DSS-induced colitis39 or EDTA-based
breaching models.40,41,42 Because normal human cells can be transplanted, the addition
of genetic alterations in multiple steps can prospectively provide new insights into
tumorigenesis. Genotype–phenotype studies can be performed to observe the phenotypic changes
that occur in vivo as a result of acquired genetic changes. In other words,
diseased cells can be created from normal cells and reproduced in vivo. For
instance, we have successfully transplanted human colonic organoids that expressed
BRAFV600E and R-spondin rearrangements (EIF3E-RSPO2 fusion) into the mouse
colon. These xenografts showed serrated changes, but further introduction of GREM1
overexpression promoted polypoid tumor formation, reproducing the unique endoscopic and
histologic features of traditional serrated adenoma (TSA), including slit-like serrations
and penicillate nuclei. These findings indicate that the ectopic expression of GREM1 is
important in pathologic changes, consistent with GREM1 upregulation in clinical TSA compared
with the level in adjacent superficially serrated adenoma.40 Prospective genetic analysis has advantages over marker-based
speculation for understanding the effects of specific genetic alterations in diseased human
tissues. In vivo studies with organoid transplantation provide pathological
insights to bridge the gap between genetic analysis and the histological phenotypes of
tumors (Fig. 2A).
This approach allows the transplantation of not only colorectal cancer but also gastric
cancer (GC) organoids.41 Diffuse GC
exhibits a unique histology and is subdivided into signet-ring cell carcinoma (SRCC) and
non-SRCC; the latter is also referred to as poorly cohesive carcinoma not otherwise
specified (PCC-NOS). In the stomach, transplantation from the mucosal surface is also useful
for research aimed at elucidating the invasion mechanism. However, there are limitations in
transplantation experiments of human GC in the mouse model, in which it is difficult to
perform gastroscopy. Taking advantage of the preserved properties of transplanted cells, we
attempted to transplant diffuse GC organoids into the rectum of mice, where colonoscopy can
easily be performed. Organoid engraftment of diffuse GC was successfully performed, and
endoscopic observation confirmed that the xenografts increased in size diffusely over time.
Interestingly, the xenotransplanted GC tissue showed heterogeneity between the luminal side
and the area near the fibroblasts where the crypt base was originally located. Because
fibroblasts provide growth factors such as Wnt and R-spondin, the histological appearance
suggested that the transplanted GC cells were differentiating in a growth factor-dependent
hierarchical manner as they moved away from the fibroblasts. In other words, PCC-NOS and
SRCC are not separate, but rather PCC-NOS differentiates into SRCC. Similar changes in
morphology related to fibroblast positioning were observed in a model in which these
organoids were transplanted into the stomach wall. SRCC is sometimes referred to as
undifferentiated-type GC, but it actually has a “differentiated” GC histology. In
vitro experiments also confirmed that PCC-NOS differentiated into SRCC when Wnt
and R-spondin were removed from the medium.41 Evidently, cells from different organs are able to remodel the colon
while retaining their unique identity.
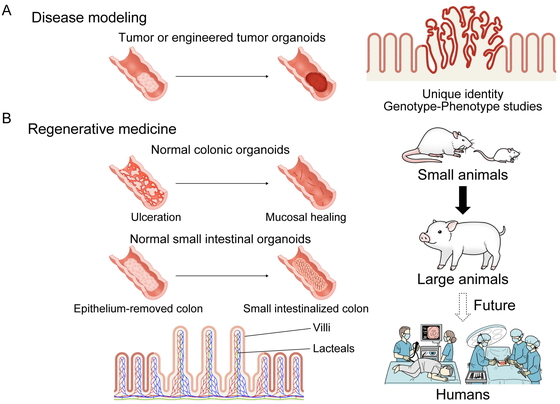
Normal Small Intestinal Organoid Transplantation
The small intestine plays an important role in sustaining life, including the digestion of
food and absorption of nutrients. For patients with UC, which is one of the most common
diseases of the large intestine, many biologics, small molecule inhibitors, and other
treatments are being actively developed, and new therapeutic agents are being launched one
after another. In contrast, there are diseases of the small intestine, once referred to as
the “dark continent” because of the difficulty of examining it, for which sufficient
treatment methods have not yet been established. Most notably, short bowel syndrome (SBS),
which occurs when a large amount of the small intestine has been surgically removed, results
in the inadequate absorption of water and nutrients, commonly leading to diarrhea,
dehydration, nutritional deficiencies, and weight loss. Because nutrient absorption from the
intestine is impaired in SBS, nutritional support is the central focus of treatment, and in
severe cases, lifelong central venous nutrition is required. There are many associated
complications of SBS, such as catheter-related infections, thrombosis, and liver
dysfunction, which can sometimes be fatal.43 Glucagon-like peptide (GLP)-2 analogs, which are promising drugs that
improve villus resorptive function,44,45 have been
approved and used worldwide, but their effectiveness is obviously limited for patients with
only a small residual length of small intestine. Intestinal lengthening procedures such as
serial transverse enteroplasty may also be effective in cases with some residual autologous
intestine,46 but are ineffective if
the remaining small intestine is too short.
The only curative treatment for SBS is intestine transplantation, in which a healthy small
intestine is transplanted into the recipient. However, in these cases, lifelong use of
immunosuppressive drugs is required, and because the small intestine is highly susceptible
to rejection compared with other organs, patient management after transplantation is often
difficult, and the 5-year graft survival rate is below 60%.47 Despite an estimated potential intestinal transplant waiting
list of fewer than 200 patients in Japan, only 35 intestinal transplants have been performed
in 31 patients from 1996 to 2020.48 In
Japan, intestinal transplant has been covered by the national health insurance system since
2018. Although the number of intestinal transplants is expected to increase in the future,
this procedure has not become a definitive treatment option.
Treatment options for SBS are significantly more limited than for diseases of other organs,
but there are high expectations for the use of regenerative medicine in this field. However,
the small intestine has a complex architecture, including crypt-villous structures composed
of diverse epithelial cells, lymphatic and vascular channels, nerves, and muscular layers;
moreover, it must also perform a diverse range of functions such as digestion, absorption,
peristalsis, and secretion. The concept of using regenerative medicine for the small
intestine has been unrealistic until now because it is extremely difficult to create the
small intestine itself outside the body and on a human scale. The same difficulties apply to
organoids because, as cells, organoids alone cannot regenerate the small intestine. However,
upon noting the similarities between the colon and small intestine below the submucosa, we
wondered whether the colon could be reconstituted as the small intestine by replacing only
the epithelium of colonic structures with the small intestine through organoid
transplantation (Fig. 2B).
We first simply adopted the method established for human colonic organoid transplantation
directly in human small intestinal organoid transplantation.49 We considered that this approach would be successful given the
previous success in transplantation of mouse small intestinal organoids into the mouse
colon.15 As expected, the transplanted
human small intestinal epithelium formed villous structures not found in the large intestine
and expressed proteins involved in digestion and absorption that are unique to the small
intestine (e.g., sucrase-isomaltase, ileal bile acid transporter). To our surprise, we
noticed that the transplantation of small intestinal organoids led to the formation of
lymphatic vascular structures unique to the small intestine located in the center of the
villi (lacteals), which are essential for lipid absorption. Furthermore, these lacteal-like
structures had the ability to absorb administered fluorescent cholesterol, indicating that
the epithelium reconstructed by organoid transplantation had an absorptive function.
However, the reconstructed villus structures were immature compared with the original mature
human small intestinal villi. Small intestinal organoid transplantation in mice15 or humans49 with only a very small epithelial replacement of the colon near the
anus with insufficient villus formation is not expected to be a treatment option to expand
small intestinal function, and therefore has no clinical significance. We considered the
need to promote the maturation of transplanted small intestinal villi and pondered how to
achieve this goal. We then focused on flow as a factor that promotes villus formation.
Luminal flow is faster in the small intestine, but slower in the large intestine. In
clinical practice, the loss of luminal flow because of dietary restrictions or stoma
construction causes villus atrophy. Together with confirmation of the effect of flow on
villus maturation in vitro as described below, we realized the importance
of reconstructing the small intestinal epithelium at the original position of the small
intestine where flow is present.
We elected to use the rat SBS model, which is larger than the mouse and therefore better
suited for establishing complex surgical techniques. Rat organoids could not be cultured
under the same culture conditions as for mice,2,3 but refined
culture conditions made it possible to establish luciferase-transgenic rat organoids for
syngeneic transplantation and bioluminescence monitoring.6,49 We then
created a “small intestinalized colon” (SIC), in which the colonic epithelium was removed,
and the small intestinal organoids were transplanted to replace the colonic epithelium. The
same rats were used as an SBS model to confirm the efficacy of the treatment. As a surgical
technique, to prevent the passage of stool to allow the transplanted organoids to grow
robustly, the 4-cm-long epithelialized intestine was temporarily attached to the abdominal
wall after organoid transplantation to allow a wide area of organoid engraftment without
bowel obstruction. However, as expected, there was no luminal flow in the transplanted area
and there was insufficient villi formation under those conditions. We then interposed the
SIC to the original site of the small intestine where flow was present, and, as we had
intended, we were able to confirm the formation of villi in the SIC. Total removal of the
jejunoileum caused weight loss and severe diarrhea, and the condition of the rats
deteriorated drastically within 2 weeks. In contrast, weight gain was observed in rats in
which the SIC was constructed, and it was remarkable to consider that the generation of only
a few centimeters of SIC could contribute to survival. However, clinical studies have
confirmed that the presence or absence of as little as 10 cm of small intestine can affect
the prognosis of human SBS patients,50,51 whose
original small intestine length was approximately 5–7 m. In fact, weight gain was observed
in SBS rats with only a few centimeters of healthy terminal ileum remaining, supporting the
finding that the generation of the SIC ensured the rats’ survival.49 Nevertheless, we considered that other parties may cast doubt
over these results, and as long as surgical experiments are performed, it may be possible
that the surgeon’s manipulation affected the results, even if unintentionally. Therefore, to
minimize experimental bias, we conducted the experiments in a blinded fashion using
organoids in which either ileum or colon organoids were transplanted into rats without
informing the surgeon and care staff of their cell types. These experiments demonstrated
prolonged survival in the ileal organoid-transplanted SBS rat group compared with the colon
organoid-transplanted group and extensive engraftment of transplanted cells in rats with
increased body weight. The SIC also formed lacteals and blood vessel networks, confirming
that it would be a functional small intestinalized graft with intestinal peristalsis and
absorption function.49
Villus Formation of the Small Intestinal Epithelium in Vitro
Inspired by the findings obtained from organoid transplantation, we investigated whether
mechanical forces on the luminal surface could be an inductive cue for villus
formation.49 To demonstrate the
involvement of flow, we aimed to evaluate villus maturation in vitro.
However, conventional 3D organoids and induced pluripotent stem-derived organoids are not
suitable as an evaluation system because they do not exhibit villus formation or mature
enterocyte marker expression. Therefore, we decided to add flow to 2D-cultured intestinal
organoids. Normally, when cultured in 2D, organoids show a monolayer structure, but when
flow was artificially added with a shaker, a villus-like structure formed. This change is
not seen in colonic organoids, indicating that the small intestinal epithelium forms
villus-like structures in a flow-dependent manner. RNA-sequencing also confirmed the
expression of mature absorptive epithelial markers, leading to the establishment of a new
in vitro culture evaluation system for small intestine
function.49
Future Perspectives
We believe that this transplantation method, for which the therapeutic concept has been
confirmed in small animals, is technically feasible and has therapeutic potential in large
animals and humans (Fig. 2B). This organoid
transplantation is attracting substantial attention as a potential game changer in the
treatment of SBS.52 Alternative
approaches, such as combining organoids with human decellularized intestinal scaffolds to
create intestinal tracts, are also possible.53,54 If the
first-in-human trial of endoscopic colonic organoid transplantation demonstrates the safety
of organoid transplantation, it will provide significant supportive data for small
intestinal organoid transplantation. However, there remains a need for further verification
of the safety of small intestinal organoid transplantation, which will require further
surgical work. We are currently conducting such research at Keio University School of
Medicine using pigs as a preclinical model. We are currently developing methods based on
surgery, but we believe that the possibilities for organoid transplantation treatment will
continue to expand in the future. These are expected to include minimally invasive
treatments combined with endoscopy and the treatment of diseases through the development of
medical devices and technologies.
Conclusions
We have developed a novel platform for epithelial cell transplantation using various types
of organoids. Organoid transplantation has great potential to promote investigation of the
biology of human diseases and to advance the development of regenerative medicine.
Acknowledgments
This work was supported by the Japan Agency for Medical Research and Development (AMED)
(grant numbers JP21ek0109523 and JP21bm0704069), AMED-CREST (grant number JP18gm1210001),
Japan Society for the Promotion of Science (grant numbers JP21K19540 and JP20H03746), the
Mochida Memorial Foundation for Medical and Pharmaceutical Research, the Takeda Science
Foundation, and Keio Gijuku Fukuzawa Memorial Fund for the Advancement of Education and
Research.
Conflicts of Interest
T.S. is the holder of several patents related to organoid culture. The remaining authors
declare that they have no conflicts of interest related to this article.
References
- 1. Barker N, van Es JH, Kuipers J, Kujala P, van den
Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H:
Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature
2007; 449: 1003–1007. PMID:17934449 https://doi.org/10.1038/nature06196
- 2. Sato T, Vries RG, Snippert HJ, van de Wetering M,
Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters PJ, Clevers H: Single Lgr5 stem
cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009;
459: 262–265. PMID:19329995 https://doi.org/10.1038/nature07935
- 3. Sato T, Stange DE, Ferrante M, Vries RG, van Es
JH, van den Brink S, van Houdt WJ, Pronk A, van Gorp J, Siersema PD, Clevers H: Long-term
expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s
epithelium. Gastroenterology 2011; 141: 1762–1772. PMID:21889923
https://doi.org/10.1053/j.gastro.2011.07.050
- 4. Fujii M, Shimokawa M, Date S, Takano A, Matano M,
Nanki K, Ohta Y, Toshimitsu K, Nakazato Y, Kawasaki K, Uraoka T, Watanabe T, Kanai T, Sato
T: A colorectal tumor organoid library demonstrates progressive loss of niche factor
requirements during tumorigenesis. Cell Stem Cell 2016; 18: 827–838. PMID:27212702
https://doi.org/10.1016/j.stem.2016.04.003
- 5. Nanki K, Toshimitsu K, Takano A, Fujii M,
Shimokawa M, Ohta Y, Matano M, Seino T, Nishikori S, Ishikawa K, Kawasaki K, Togasaki K,
Takahashi S, Sukawa Y, Ishida H, Sugimoto S, Kawakubo H, Kim J, Kitagawa Y, Sekine S, Koo
BK, Kanai T, Sato T: Divergent routes toward Wnt and R-spondin niche independency during
human gastric carcinogenesis. Cell 2018; 174: 856–869.e17. PMID:30096312
https://doi.org/10.1016/j.cell.2018.07.027
- 6. Fujii M, Matano M, Toshimitsu K, Takano A, Mikami
Y, Nishikori S, Sugimoto S, Sato T: Human intestinal organoids maintain self-renewal
capacity and cellular diversity in niche-inspired culture condition. Cell Stem Cell 2018;
23: 787–793.e6. PMID:30526881 https://doi.org/10.1016/j.stem.2018.11.016
- 7. Huch M, Gehart H, van Boxtel R, Hamer K, Blokzijl
F, Verstegen MM, Ellis E, van Wenum M, Fuchs SA, de Ligt J, van de Wetering M, Sasaki N,
Boers SJ, Kemperman H, de Jonge J, Ijzermans JN, Nieuwenhuis EE, Hoekstra R, Strom S,
Vries RR, van der Laan LJ, Cuppen E, Clevers H: Long-term culture of genome-stable
bipotent stem cells from adult human liver. Cell 2015; 160: 299–312. PMID:25533785
https://doi.org/10.1016/j.cell.2014.11.050
- 8. Seino T, Kawasaki S, Shimokawa M, Tamagawa H,
Toshimitsu K, Fujii M, Ohta Y, Matano M, Nanki K, Kawasaki K, Takahashi S, Sugimoto S,
Iwasaki E, Takagi J, Itoi T, Kitago M, Kitagawa Y, Kanai T, Sato T: Human pancreatic tumor
organoids reveal loss of stem cell niche factor dependence during disease progression.
Cell Stem Cell 2018; 22: 454–467.e6. PMID:29337182
https://doi.org/10.1016/j.stem.2017.12.009
- 9. Ebisudani T, Sugimoto S, Haga K, Mitsuishi A,
Takai-Todaka R, Fujii M, Toshimitsu K, Hamamoto J, Sugihara K, Hishida T, Asamura H,
Fukunaga K, Yasuda H, Katayama K, Sato T: Direct derivation of human alveolospheres for
SARS-CoV-2 infection modeling and drug screening. Cell Rep 2021; 35: 109218. PMID:34038715
https://doi.org/10.1016/j.celrep.2021.109218
- 10. Wakisaka Y, Sugimoto S, Sato T: Organoid medicine
for inflammatory bowel disease. Stem Cells 2022; 40: 123–132. PMID:35258629
https://doi.org/10.1093/stmcls/sxab020
- 11. Sugimoto S, Sato T: Establishment of 3D
intestinal organoid cultures from intestinal stem cells. Methods Mol Biol 2017; 1612:
97–105. PMID:28634937 https://doi.org/10.1007/978-1-4939-7021-6_7
- 12. Nanki K, Fujii M, Shimokawa M, Matano M,
Nishikori S, Date S, Takano A, Toshimitsu K, Ohta Y, Takahashi S, Sugimoto S, Ishimaru K,
Kawasaki K, Nagai Y, Ishii R, Yoshida K, Sasaki N, Hibi T, Ishihara S, Kanai T, Sato T:
Somatic inflammatory gene mutations in human ulcerative colitis epithelium. Nature 2020;
577: 254–259. PMID:31853059 https://doi.org/10.1038/s41586-019-1844-5
- 13. Yui S, Nakamura T, Sato T, Nemoto Y, Mizutani T,
Zheng X, Ichinose S, Nagaishi T, Okamoto R, Tsuchiya K, Clevers H, Watanabe M: Functional
engraftment of colon epithelium expanded in vitro from a single adult Lgr5+ stem cell. Nat
Med 2012; 18: 618–623. PMID:22406745 https://doi.org/10.1038/nm.2695
- 14. Fordham RP, Yui S, Hannan NR, Soendergaard C,
Madgwick A, Schweiger PJ, Nielsen OH, Vallier L, Pedersen RA, Nakamura T, Watanabe M,
Jensen KB: Transplantation of expanded fetal intestinal progenitors contributes to colon
regeneration after injury. Cell Stem Cell 2013; 13: 734–744. PMID:24139758
https://doi.org/10.1016/j.stem.2013.09.015
- 15. Fukuda M, Mizutani T, Mochizuki W, Matsumoto T,
Nozaki K, Sakamaki Y, Ichinose S, Okada Y, Tanaka T, Watanabe M, Nakamura T: Small
intestinal stem cell identity is maintained with functional Paneth cells in
heterotopically grafted epithelium onto the colon. Genes Dev 2014; 28: 1752–1757.
PMID:25128495 https://doi.org/10.1101/gad.245233.114
- 16. Yui S, Azzolin L, Maimets M, Pedersen MT, Fordham
RP, Hansen SL, Larsen HL, Guiu J, Alves MR, Rundsten CF, Johansen JV, Li Y, Madsen CD,
Nakamura T, Watanabe M, Nielsen OH, Schweiger PJ, Piccolo S, Jensen KB: YAP/TAZ-dependent
reprogramming of colonic epithelium links ECM remodeling to tissue regeneration. Cell Stem
Cell 2018; 22: 35–49.e7. PMID:29249464
https://doi.org/10.1016/j.stem.2017.11.001
- 17. Guiu J, Hannezo E, Yui S, Demharter S,
Ulyanchenko S, Maimets M, Jørgensen A, Perlman S, Lundvall L, Mamsen LS, Larsen A, Olesen
RH, Andersen CY, Thuesen LL, Hare KJ, Pers TH, Khodosevich K, Simons BD, Jensen KB:
Tracing the origin of adult intestinal stem cells. Nature 2019; 570: 107–111.
PMID:31092921 https://doi.org/10.1038/s41586-019-1212-5
- 18. Bergenheim F, Seidelin JB, Pedersen MT, Mead BE,
Jensen KB, Karp JM, Nielsen OH: Fluorescence-based tracing of transplanted intestinal
epithelial cells using confocal laser endomicroscopy. Stem Cell Res Ther 2019; 10: 148.
PMID:31133056 https://doi.org/10.1186/s13287-019-1246-5
- 19. Jee J, Jeong SY, Kim HK, Choi SY, Jeong S, Lee J,
Ko JS, Kim MS, Kwon MS, Yoo J: In vivo evaluation of scaffolds compatible for colonoid
engraftments onto injured mouse colon epithelium. FASEB J 2019; 33: 10116–10125.
PMID:31211931 https://doi.org/10.1096/fj.201802692RR
- 20. Jee JH, Lee DH, Ko J, Hahn S, Jeong SY, Kim HK,
Park E, Choi SY, Jeong S, Lee JW, Cho HJ, Kwon MS, Yoo J: Development of collagen-based 3D
matrix for gastrointestinal tract-derived organoid culture. Stem Cells Int 2019; 2019:
1–15. PMID:31312220 https://doi.org/10.1155/2019/8472712
- 21. Jee J, Park JH, Im JH, Kim MS, Park E, Lim T,
Choi WH, Kim JH, Kim WR, Ko JS, Jeong SY, Ko SY, Lee JI, Lee KJ, Jeon H, Seo JH, Hwang DY,
Shin HS, Yoo J: Functional recovery by colon organoid transplantation in a mouse model of
radiation proctitis. Biomaterials 2021; 275: 120925. PMID:34171755
https://doi.org/10.1016/j.biomaterials.2021.120925
- 22. Shimokawa M, Ohta Y, Nishikori S, Matano M,
Takano A, Fujii M, Date S, Sugimoto S, Kanai T, Sato T: Visualization and targeting of
LGR5+ human colon cancer stem cells. Nature 2017; 545: 187–192. PMID:28355176
https://doi.org/10.1038/nature22081
- 23. Ito M, Hiramatsu H, Kobayashi K, Suzue K,
Kawahata M, Hioki K, Ueyama Y, Koyanagi Y, Sugamura K, Tsuji K, Heike T, Nakahata T:
NOD/SCID/γcnull mouse: an excellent recipient mouse model for engraftment of human cells.
Blood 2002; 100: 3175–3182. PMID:12384415
https://doi.org/10.1182/blood-2001-12-0207
- 24. Sugimoto S, Ohta Y, Fujii M, Matano M, Shimokawa
M, Nanki K, Date S, Nishikori S, Nakazato Y, Nakamura T, Kanai T, Sato T: Reconstruction
of the human colon epithelium in vivo. Cell Stem Cell 2018; 22: 171–176.e5. PMID:29290616
https://doi.org/10.1016/j.stem.2017.11.012
- 25. Sugimoto S, Fujii M, Sato T: Organoid derivation
and orthotopic xenotransplantation for studying human intestinal stem cell dynamics.
Methods Mol Biol 2020; 2171: 303–320. PMID:32705652
https://doi.org/10.1007/978-1-0716-0747-3_21
- 26. Ishikawa K, Sugimoto S, Oda M, Fujii M, Takahashi
S, Ohta Y, Takano A, Ishimaru K, Matano M, Yoshida K, Hanyu H, Toshimitsu K, Sawada K,
Shimokawa M, Saito M, Kawasaki K, Ishii R, Taniguchi K, Imamura T, Kanai T, Sato T:
Identification of Quiescent LGR5+ Stem Cells in the Human Colon. Gastroenterology 2022;
163: 1391–1406.e24. PMID: 35963362.
https://doi.org/10.1053/j.gastro.2022.07.081
- 27. Sugimoto S, Sato T: Organoid vs in vivo mouse
model: which is better research tool to understand the biologic mechanisms of intestinal
epithelium? Cell Mol Gastroenterol Hepatol 2022; 13: 195–197. PMID:34644539
https://doi.org/10.1016/j.jcmgh.2021.06.027
- 28. Sugimoto S, Sato T: Rebuttal to: in vivo studies
should take priority when defining mechanisms of intestinal crypt morphogenesis. Cell Mol
Gastroenterol Hepatol 2022; 13: 5. PMID:34644540
https://doi.org/10.1016/j.jcmgh.2021.09.010
- 29. Mitoyan L, Chevrier V, Hernandez-Vargas H,
Ollivier A, Homayed Z, Pannequin J, Poizat F, De Biasi-Cador C, Charafe-Jauffret E,
Ginestier C, Guasch G: A stem cell population at the anorectal junction maintains
homeostasis and participates in tissue regeneration. Nat Commun 2021; 12: 2761.
PMID:33980830 https://doi.org/10.1038/s41467-021-23034-x
- 30. Sugimoto S, Iwao Y, Shimoda M, Takabayashi K,
Sato T, Kanai T, Mutaguchi M, Nanki K, Okabayashi K, Kawaida M, Aoki Y, Yoshimatsu Y,
Kiyohara H, Kawaguchi T, Mikami Y, Fukuhara K, Sujino T, Hosoe N, Ogata H, Yahagi N, Keio
IBD Collaborators: Epithelium replacement contributes to field expansion of squamous
epithelium and ulcerative colitis-associated neoplasia. Gastroenterology 2022; 162:
334–337.e5. PMID:34597671 https://doi.org/10.1053/j.gastro.2021.09.051
- 31. Sugimoto S, Naganuma M, Iwao Y, Matsuoka K,
Shimoda M, Mikami S, Mizuno S, Nakazato Y, Nanki K, Inoue N, Ogata H, Kanai T: Endoscopic
morphologic features of ulcerative colitis–associated dysplasia classified according to
the SCENIC consensus statement. Gastrointest Endosc 2017; 85: 639–646.e2. PMID:27884517
https://doi.org/10.1016/j.gie.2016.11.013
- 32. Kakiuchi N, Yoshida K, Uchino M, Kihara T, Akaki
K, Inoue Y, Kawada K, Nagayama S, Yokoyama A, Yamamoto S, Matsuura M, Horimatsu T, Hirano
T, Goto N, Takeuchi Y, Ochi Y, Shiozawa Y, Kogure Y, Watatani Y, Fujii Y, Kim SK, Kon A,
Kataoka K, Yoshizato T, Nakagawa MM, Yoda A, Nanya Y, Makishima H, Shiraishi Y, Chiba K,
Tanaka H, Sanada M, Sugihara E, Sato T, Maruyama T, Miyoshi H, Taketo MM, Oishi J, Inagaki
R, Ueda Y, Okamoto S, Okajima H, Sakai Y, Sakurai T, Haga H, Hirota S, Ikeuchi H, Nakase
H, Marusawa H, Chiba T, Takeuchi O, Miyano S, Seno H, Ogawa S: Frequent mutations that
converge on the NFKBIZ pathway in ulcerative colitis. Nature 2020; 577: 260–265.
PMID:31853061 https://doi.org/10.1038/s41586-019-1856-1
- 33. Matano M, Date S, Shimokawa M, Takano A, Fujii M,
Ohta Y, Watanabe T, Kanai T, Sato T: Modeling colorectal cancer using CRISPR-Cas9–mediated
engineering of human intestinal organoids. Nat Med 2015; 21: 256–262. PMID:25706875
https://doi.org/10.1038/nm.3802
- 34. de Sousa e Melo F, Kurtova AV, Harnoss JM,
Kljavin N, Hoeck JD, Hung J, Anderson JE, Storm EE, Modrusan Z, Koeppen H, Dijkgraaf GJ,
Piskol R, de Sauvage FJ: A distinct role for Lgr5+ stem cells in primary and metastatic
colon cancer. Nature2017; 543: 676–680.
PMID:28358093https://doi.org/10.1038/nature21713
- 35. Roper J, Tammela T, Cetinbas NM, Akkad A,
Roghanian A, Rickelt S, Almeqdadi M, Wu K, Oberli MA, Sánchez-Rivera FJ, Park YK, Liang X,
Eng G, Taylor MS, Azimi R, Kedrin D, Neupane R, Beyaz S, Sicinska ET, Suarez Y, Yoo J,
Chen L, Zukerberg L, Katajisto P, Deshpande V, Bass AJ, Tsichlis PN, Lees J, Langer R,
Hynes RO, Chen J, Bhutkar A, Jacks T, Yilmaz ÖH: In vivo genome editing and organoid
transplantation models of colorectal cancer and metastasis. Nat Biotechnol 2017; 35:
569–576. PMID:28459449 https://doi.org/10.1038/nbt.3836
- 36. Fumagalli A, Drost J, Suijkerbuijk SJ, van Boxtel
R, de Ligt J, Offerhaus GJ, Begthel H, Beerling E, Tan EH, Sansom OJ, Cuppen E, Clevers H,
van Rheenen J: Genetic dissection of colorectal cancer progression by orthotopic
transplantation of engineered cancer organoids. Proc Natl Acad Sci USA 2017; 114:
E2357–E2364.
- 37. O’Rourke KP, Loizou E, Livshits G, Schatoff EM,
Baslan T, Manchado E, Simon J, Romesser PB, Leach B, Han T, Pauli C, Beltran H, Rubin MA,
Dow LE, Lowe SW: Transplantation of engineered organoids enables rapid generation of
metastatic mouse models of colorectal cancer. Nat Biotechnol 2017; 35: 577–582.
PMID:28459450 https://doi.org/10.1038/nbt.3837
- 38. Lannagan TR, Lee YK, Wang T, Roper J, Bettington
ML, Fennell L, Vrbanac L, Jonavicius L, Somashekar R, Gieniec K, Yang M, Ng JQ, Suzuki N,
Ichinose M, Wright JA, Kobayashi H, Putoczki TL, Hayakawa Y, Leedham SJ, Abud HE, Yilmaz
ÖH, Marker J, Klebe S, Wirapati P, Mukherjee S, Tejpar S, Leggett BA, Whitehall VL,
Worthley DL, Woods SL: Genetic editing of colonic organoids provides a molecularly
distinct and orthotopic preclinical model of serrated carcinogenesis. Gut 2019; 68:
684–692. PMID:29666172 https://doi.org/10.1136/gutjnl-2017-315920
- 39. Ganesh K, Wu C, O’Rourke KP, Szeglin BC, Zheng Y,
Sauvé CE, Adileh M, Wasserman I, Marco MR, Kim AS, Shady M, Sanchez-Vega F, Karthaus WR,
Won HH, Choi SH, Pelossof R, Barlas A, Ntiamoah P, Pappou E, Elghouayel A, Strong JS, Chen
CT, Harris JW, Weiser MR, Nash GM, Guillem JG, Wei IH, Kolesnick RN, Veeraraghavan H,
Ortiz EJ, Petkovska I, Cercek A, Manova-Todorova KO, Saltz LB, Lavery JA, DeMatteo RP,
Massagué J, Paty PB, Yaeger R, Chen X, Patil S, Clevers H, Berger MF, Lowe SW, Shia J,
Romesser PB, Dow LE, Garcia-Aguilar J, Sawyers CL, Smith JJ: A rectal cancer organoid
platform to study individual responses to chemoradiation. Nat Med 2019; 25: 1607–1614.
PMID:31591597 https://doi.org/10.1038/s41591-019-0584-2
- 40. Kawasaki K, Fujii M, Sugimoto S, Ishikawa K,
Matano M, Ohta Y, Toshimitsu K, Takahashi S, Hosoe N, Sekine S, Kanai T, Sato T:
Chromosome engineering of human colon-derived organoids to develop a model of traditional
serrated adenoma. Gastroenterology 2020; 158: 638–651.e8. PMID:31622618
https://doi.org/10.1053/j.gastro.2019.10.009
- 41. Togasaki K, Sugimoto S, Ohta Y, Nanki K, Matano
M, Takahashi S, Fujii M, Kanai T, Sato T: Wnt signaling shapes the histologic variation in
diffuse gastric cancer. Gastroenterology 2021; 160: 823–830. PMID:33217450
https://doi.org/10.1053/j.gastro.2020.10.047
- 42. Watanabe S, Nishimura R, Shirasaki T, Katsukura
N, Hibiya S, Kirimura S, Negi M, Okamoto R, Matsumoto Y, Nakamura T, Watanabe M, Tsuchiya
K: Schlafen 11 is a novel target for mucosal regeneration in ulcerative colitis. J Crohn’s
Colitis 2021; 15: 1558–1572. PMID:33596306
https://doi.org/10.1093/ecco-jcc/jjab032
- 43. Massironi S, Cavalcoli F, Rausa E, Invernizzi P,
Braga M, Vecchi M: Understanding short bowel syndrome: Current status and future
perspectives. Dig Liver Dis 2020; 52: 253–261. PMID:31892505
https://doi.org/10.1016/j.dld.2019.11.013
- 44. Jeppesen PB, Pertkiewicz M, Messing B, Iyer K,
Seidner DL, O’keefe SJ, Forbes A, Heinze H, Joelsson B: Teduglutide reduces need for
parenteral support among patients with short bowel syndrome with intestinal failure.
Gastroenterology 2012; 143: 1473–1481.e3. PMID:22982184
https://doi.org/10.1053/j.gastro.2012.09.007
- 45. Drucker DJ, Yusta B: Physiology and pharmacology
of the enteroendocrine hormone glucagon-like peptide-2. Annu Rev Physiol 2014; 76:
561–583. PMID:24161075
https://doi.org/10.1146/annurev-physiol-021113-170317
- 46. Thompson JS, Weseman R, Rochling FA, Mercer DF:
Current management of the short bowel syndrome. Surg Clin North Am 2011; 91: 493–510.
PMID:21621693 https://doi.org/10.1016/j.suc.2011.02.006
- 47. Grant D, Abu-Elmagd K, Mazariegos G, Vianna R,
Langnas A, Mangus R, Farmer DG, Lacaille F, Iyer K, Fishbein T, Intestinal Transplant
Association: Intestinal transplant registry report: global activity and trends. Am J
Transplant 2015; 15: 210–219. PMID:25438622
https://doi.org/10.1111/ajt.12979
- 48. Association TJIRaT: A report from the Japanese
Intestinal Transplantation Registry. Ishoku 2021; 56: 265–271.
- 49. Sugimoto S, Kobayashi E, Fujii M, Ohta Y, Arai K,
Matano M, Ishikawa K, Miyamoto K, Toshimitsu K, Takahashi S, Nanki K, Hakamata Y, Kanai T,
Sato T: An organoid-based organ-repurposing approach to treat short bowel syndrome. Nature
2021; 592: 99–104. PMID:33627870
https://doi.org/10.1038/s41586-021-03247-2
- 50. Dorney SF, Ament ME, Berquist WE, Vargas JH,
Hassall E: Improved survival in very short small bowelof infancy with use of long-term
parenteral nutrition. J Pediatr 1985; 107: 521–525. PMID:3930677
https://doi.org/10.1016/S0022-3476(85)80008-1
- 51. Demehri FR, Stephens L, Herrman E, West B,
Mehringer A, Arnold MA, Brown PI, Teitelbaum DH: Enteral autonomy in pediatric short bowel
syndrome: predictive factors one year after diagnosis. J Pediatr Surg 2015; 50: 131–135.
PMID:25598109 https://doi.org/10.1016/j.jpedsurg.2014.10.011
- 52. Gentilini MV, Rumbo M, Gondolesi GE: Organoid
transplantation: new avenues to treat short bowel syndrome. Transplantation 2021; 105:
2130–2131. PMID:34591815 https://doi.org/10.1097/TP.0000000000003833
- 52. Meran L, Massie I, Campinoti S, Weston AE,
Gaifulina R, Tullie L, Faull P, Orford M, Kucharska A, Baulies A, Novellasdemunt L,
Angelis N, Hirst E, König J, Tedeschi AM, Pellegata AF, Eli S, Snijders AP, Collinson L,
Thapar N, Thomas GM, Eaton S, Bonfanti P, De Coppi P, Li VS: Engineering transplantable
jejunal mucosal grafts using patient-derived organoids from children with intestinal
failure. Nat Med 2020; 26: 1593–1601. PMID:32895569
https://doi.org/10.1038/s41591-020-1024-z
- 53. Fujii M, Sugimoto S, Sato T: Linking human
intestinal scaffolds and organoids to combat intestinal failure. Nat Med 2020; 26:
1517–1518. PMID:32968235 https://doi.org/10.1038/s41591-020-1096-9
