Reviews
Unlocking the therapeutic potential of artificial metalloenzymes
2020 Volume 96 Issue 3 Pages 79-94
Details
2020 Volume 96 Issue 3 Pages 79-94
In order to harness the functionality of metals, nature has evolved over billions of years to utilize metalloproteins as key components in numerous cellular processes. Despite this, transition metals such as ruthenium, palladium, iridium, and gold are largely absent from naturally occurring metalloproteins, likely due to their scarcity as precious metals. To mimic the evolutionary process of nature, the field of artificial metalloenzymes (ArMs) was born as a way to benefit from the unique chemoselectivity and orthogonality of transition metals in a biological setting. In its current state, numerous examples have successfully incorporated transition metals into a variety of protein scaffolds. Using these ArMs, many examples of new-to-nature reactions have been carried out, some of which have shown substantial biocompatibility. Given the rapid rate at which this field is growing, this review aims to highlight some important studies that have begun to take the next step within this field; namely the development of ArM-centered drug therapies or biotechnological tools.
With the abundance of metals present in the earth’s ecosystem, it is not surprising that biological systems have evolved to integrate them as key components that drive cellular functions and metabolism. To do this, nature has employed metalloproteins, which are proteins that have evolved to bind metals with high affinity and specificity. Of particular importance are metalloenzymes, which are found in nature to carry out at least one step in all biological pathways.1) Some notable examples of metalloenzymes (and their metal coenzyme) include ascorbic acid oxidase (copper), alcohol dehydrogenase (zinc), glutathione peroxidase (selenium), and xanthine oxidase (molybdenum).
With the rise of bio-orthogonal chemistry, defined as reactions that can occur inside living systems without interference from native biochemical processes, it has been recognized that the chemoselectivity of abiotic transition metals (e.g., ruthenium, palladium, iridium, gold, etc.) may have major implications in drug therapy. To harness this potential, interest in developing new-to-nature reactions in living systems has ballooned over the past decade.
Transition metals are known to be easily inactivated by biological components; therefore, one way to introduce them safely into living systems is through the use of artificial metalloenzymes (ArMs). Broadly defined as proteins that have been restructured or recomposed to catalyze reactions not previously observed for the native protein,2)–10) researchers have pursued their development through many different approaches, as summarized in Fig. 1.

(Color online) Approaches for ArM development. (a) Repurposing natural metalloenzymes, (b) Protein fold (re)design, (c) Metal-exchange with natural cofactors, and (d) Metal-introduction through ligand anchoring.
One approach to develop ArMs can be taken through repurposing natural metalloenzymes.11)–15) Here, the focus is on identifying enzymes with the capacity to catalyze new-to-nature reactions. To do this, these studies typically revolve around screening or directed evolution-based studies. In one notable example, cytochrome c from Rhodothermus marinus was enhanced via directed evolution to catalyze carbon–silicon bond formation.14)
Although not extensively pursued in the literature, there are significant industrial and biomedical implications of ArMs designed through protein fold (re)design.16)–18) In this approach, computational modeling is used to either design a protein de novo or to redesign protein folds in order to carry out the intended reactions. Despite major challenges, several research groups have already taken steps to break through these barriers, such as the development of a Zn-based hydrolytic metalloenzyme.16)
The most common and pragmatic approach to develop ArMs is through introducing artificial cofactors into protein scaffolds. This can be done either through metal exchange via existing metal cofactors or through metal introduction via ligand anchoring. For metal exchange, a plethora of research has largely focused on exploiting heme, a natural iron–porphyrin complex that benefits from 1) strong metal binding properties, and 2) prevalence in a number of biologically important metalloproteins such as cytochromes, catalases, myoglobin, etc. As a result, numerous studies have developed ArMs where Fe in heme can be replaced with transition metals including Mn, Co, and Ir.19)–23)
In one final approach for ArM development, several research groups have relied on binding catalytic metal complexes into protein binding pockets through ligand anchoring. The clearest benefit of this strategy is the adaptability to different protein scaffolds depending on the desired functionality, biodistribution, and immunological properties. As such, numerous groups have employed this strategy to develop ArMs.
The overall purpose of this review is to highlight recent progress in the field of ArM development, with emphasis on examples that have been shown to be functionally active in biological conditions. Although the field is still in its infancy, this review aims to also feature foundational works that have begun to explore the application of ArM as future drug therapies or as biotechnological tools.
Pioneered by Frances Arnold, the 2018 Nobel laureate, heme-based proteins have been extensively repurposed for new-to-nature reactions. One such focus area is carbene transfer reactions, which include aldehyde olefination,24) alkyne cyclopropenation,25) alkene cyclopropanation,11),12),26)–35) the Doyle–Kirmse reaction,36) N–H bond insertion,37),38) S–H bond insertion,33),39) Si–H bond insertion,14) and B–H bond insertion.40)–42) Crystal structures and computational modeling have revealed that these reactions all likely proceed via a reactive iron porphyrin carbene.43) Another area of ArM exploration involves the study of heme-dependent intramolecular C–H bond aminations. In these studies, various benzenesulfonyl azides have been shown to proceed with high enantioselectivity.22),44),45) As an extension, intermolecular C–H amination was also reported,13) as well as intramolecular C–H amination using iridium-porphyrin complexes.21) Furthermore, the capacity of heme-based ArMs to undergo intermolecular aziridination46) and sulfimidation47),48) has also been reported. Although many of these heme-based ArMs were shown to be catalytically active in biological conditions, as summarized in Table 1, their applicability to medicine or biotechnology still remains largely underexplored.

To a much smaller degree, copper-based ArMs have been investigated as artificial Diels–Alderases.49)–53) Although most are limited to in vitro studies, one notable work showed the construction of a copper-based ArM using the wild-type human A2A adenosine receptor.53) As depicted in Fig. 2, the A2A adenosine receptor is expressed on the surface of human embryonic kidney (HEK) cells. With the introduction of a Cu(II) complex–A2A antagonist, the ArM was constructed directly on the surface of living HEK cells using the wild-type adenosine receptor as a protein scaffold. The resulting cells were then shown to enantioselectively facilitate Diels–Alder cycloaddition between cyclopentadiene and azachalcone.53)
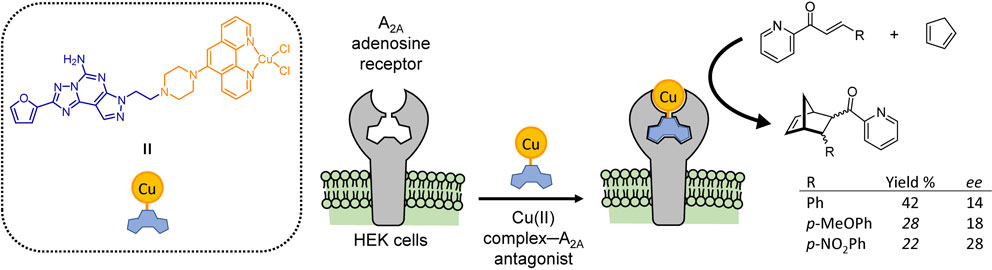
(Color online) Bioactive copper-based ArMs on living cell surfaces. Concept of an A2A adenosine receptor ArM constructed directly on living cell surfaces via anchoring with a preladenant derivative linked to a Cu(II) catalyst complex. Diels–Alder cycloaddition between cyclopentadiene and azachalcone can proceed enantioselectively on the cell surface.
Another approach to develop ArMs is by introducing abiotic transition metal cofactors. The idea is that catalysis mediated by transition metals can be both orthogonal and chemoselective in a biological environment. Additionally, existing biotechnological techniques (directed evolution, etc.) can be applied to optimize or manipulate protein properties to increase activity or substrate selectivity. Currently in the literature, there are numerous reports of ArMs that have been integrated with abiotic transition metals. These include the use of rhodium,54)–66) iridium,19),67)–71) manganese,23),72)–78) ruthenium,69),79)–97) vanadium,98) and chromium.99) For this review, however, importance is mainly placed on examples of ArMs that have been shown to be catalytically active in biological conditions. These noteworthy studies can be viewed as potential foundational pieces for future biomedical research.
Looking at rhodium-based ArMs, several examples have been shown to be catalytically active in biological conditions. For example, one study engineered a streptavidin scaffold to be bound with several different types of biotin-dirhodium complexes. These ArMs were screened and revealed to be catalytically more efficient (reactions include carbene insertion into C–H bonds and intermolecular cyclopropanation) compared with its free-in-solution metal equivalent Rh2(OAc)4.54) In addition, with a cell display platform to secrete streptavidin into the E. coli periplasm by fusion with an outer membrane protein A (OmpA) signal peptide, these dirhodium-based ArMs displayed catalytic activity up to 20 turnovers. For another relevant study, as shown in Fig. 3, a nitrobindin variant was fused with an inactivated esterase autotransporter (EstA) in order to be shuttled to microbial cell surfaces.59) In this case, construction of the rhodium-based ArM was done through covalent linkage between Cys96 of the protein and a Rh(III)-maleimide complex. Subsequent studies then showed that phenylacetylene could be polymerized to PPA (also known as poly(phenylacetylene)) with high favorability to the trans isomer at a high turnover (39 × 106 per cell) on E. coli surfaces.

(Color online) Bioactive rhodium-based ArMs on living cell surfaces. This concept first involves the shuttling of nitrobindin to E. coli membrane surfaces via EstA. Construction of the ArM is done via covalent attachment of a Rh(III)-maleimide complex with Cys96 of the nitrobindin protein. Polymerization of phenylacetylene has been shown to favor the production of trans-PPA with a high TON.
For iridium-based ArMs, research groups have mainly explored their usage as an artificial transfer hydrogenase (ATHase).67)–71),100) Some studies have also focused on their use for carbene insertion into C–H bonds, or internal and aliphatic olefins.19),101) In one notable study, a triggered release methodology was developed using an Ir-based ATHase and a quinolinium substrate.71) Following reduction of the substrate by the ATHase, self-immolation of the substrate can then proceed to release an aryl alcohol moiety (e.g., fluorescent umbelliferone). In addition to highlighting its potential for adaptation to drug therapy, this system was also shown to be biocompatible when expressed in the periplasm of E. coli.
For manganese-based ArMs, a host of in vitro studies have focused on catalyzing reactions such as olefin epoxidation,72),74)–77) and oxidation of a number of different substrate types.23),72),73),76),78) Unfortunately, the current literature does not yet contain reports of Mn-based ArM activity that can proceed in biological conditions.
Although currently underexplored, gold-based ArMs are also another potential avenue for investigation. As one leading example, our group developed an early framework for a gold-based artificial metalloenzyme. In this approach, human serum albumin was chosen as the protein scaffold due to its well-known role as a drug-binding protein with high serum stability. To construct the ArM, an Au(III) catalyst-coumarin complex was used to direct shallow binding into a hydrophobic binding pocket. In this manner, the ArM can be used to facilitate gold-catalyzed propargyl ester labeling, which was shown to require a 2-benzoylpyridine ligand to create an active intermediate via aryl–alkynyl cross coupling.102) As a result, lysine residues on surface proteins can be labelled via propargyl ester-functionalized probes, as depicted in Fig. 4.103)

(Color online) Bioactive gold-based ArMs for protein labelling. Concept of protein labelling using gold-based ArMs to catalyze the nucleophilic addition of amines (lysine residues of proteins) to propargyl ester-based probes. Construction of the ArM is done via anchoring with a coumarin derivative linked to an Au(III) complex.
Finally, ruthenium-based ArMs have been extensively developed to carry out reactions such as ring-closing metathesis (RCM),69),79),84)–90),93),94) ring-opening metathesis polymerization (ROMP),91)–94) and alloc uncaging.95)–97) Despite the abundance of current literature focused on ruthenium-based ArMs, its transition and usage as a drug therapy in living biological systems still remains at an early state. However, with the progress gained by several key pioneering studies, ArM usage in a therapeutic setting may soon become a reality.
In one of the earliest works highlighting the biocompatibility of ruthenium-based ArMs, a cell-surface display system was designed where streptavidin could be secreted into the E. coli periplasm by fusion with an N-terminal signal peptide from OmpA.79) Using this approach, directed evolution studies were then carried out to improve the catalytic activity of streptavidin-based ArMs for metathesis and alloc-cleavage in biological conditions (Fig. 5). Results from these studies showed that with a streptavidin pentamutant (V47A, N49K, T114Q, A119G, K121R), the catalytic efficiency of ring-closing metathesis could be improved by two-fold compared with the wild-type.79) In the case of alloc deprotection, a double mutation of streptavidin (S112M, K121A) showed an almost six-fold improvement in turnover compared with the wild-type.97)

(Color online) Bioactive ruthenium-based ArMs on living cell surfaces via cell surface display of a Lpp-OmpA-fused streptavidin ArM, which is secreted into the periplasmic space and anchored to the E. coli outer membrane.
To develop the concept of cell penetrating ArMs, another report adapted the use of cell-penetrating poly(disulfide) polymers.95) In this approach, the ligand–catalyst complex is directly attached to a cell-penetrating poly(disulfide) polymer, which will then elicit cellular uptake by first undergoing disulfide exchange with cell surface thiols, before eventual release by cytosol glutathione.104),105) Using this technology, ruthenium-containing streptavidin ArMs were shown to be effectively taken up into HEK-293T cells. To further highlight its utility, a system was designed (Fig. 6a), where double deprotection of a molecule can release the hormone triiodothyronine (T3). In the presence of a triiodothyronine-responsive gene switch, this system could then go on to control and upregulate the activity of genetically encoded genes.95)

(Color online) Bioactive ruthenium-based ArMs. (a) Concept of cell-penetrating ArM in conjunction with gene activation via double decaging of the triiodothyronine (T3) hormone. With a triiodothyronine-responsive gene switch, genetically encoded genes (e.g., Sec-nluc) can be controlled and upregulated. (b) Concept of streptavidin ArM anchored directly onto cell surfaces via NHS ester chemistry.
In another study of interest, the eukaryote Chlamydomonas reinhardtii was functionalized with Ru-based ArMs.96) This approach, as shown in Fig. 6b, was accomplished first by anchoring a biotin moiety onto the cell surface via well-known N-hydroxysuccinimide (NHS) ester chemistry. Given that streptavidin is tetrameric in nature, it has the capacity to simultaneously bind both the cell-anchored biotin and a biotin-ruthenium catalyst. To highlight a possible application to drug therapy, profluorophore activation was investigated with alloc-coumarin, where experiments were able to show the possibility of on-cell catalysis.
Currently, one of the challenges preventing in vivo usage of ArMs is the susceptibility of abiotic metals to be quenched by cellular metabolites, such as glutathione (GSH). To address this issue, our group explored the idea of developing ArMs resistant to glutathione quenching.90) The theory behind this approach exploited the deep hydrophobic binding pockets of albumin-based ArMs to ensure that hydrophilic metabolites (e.g., GSH) could be blocked from interacting with the embedded metal catalyst (Fig. 7). As a result, a ruthenium-bound albumin ArM (alb-Ru) that catalyzes ring-closing metathesis was shown to be active even in the presence of up to 1000× equivalents of GSH.
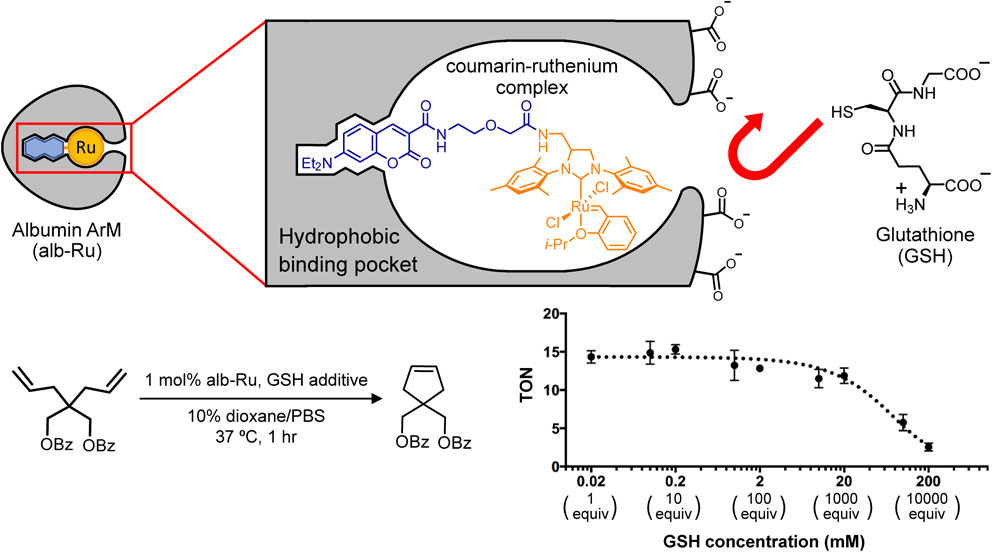
(Color online) Albumin ArMs were found to be capable of protecting metal catalysts anchored inside its hydrophobic binding pocket. For example, an albumin ArM that catalyzes ruthenium-based metathesis was found to be resistant to quenching by up to 1000× equivalents of glutathione (GSH).
Another avenue for ArM development lies in biosensor applications, where they can potentially be used as biotechnological tools to rapidly monitor metabolites in a spatiotemporal manner. In general, enzymes biosensors for diagnostic purposes have found utility not only from a scientific perspective but also from a commercial standpoint. Despite this, one of the major issues impeding enzyme biosensor progress is that they are often derived from pre-existing natural enzymes. As such, current approaches are largely constrained to metabolites within a limited chemical space. With the rising development of ArMs, a unique opportunity exists to design biosensors for difficult-to-detect metabolites.
Shown in Fig. 8, our group has developed one of the first examples of an ArM-based biosensor, which is used to detect the plant hormone, ethylene gas.106) Based on the approach shown in Fig. 8a, the albumin scaffold is used to solubilize and protect a quenched ruthenium catalyst complex. In the presence of ethylene, ruthenium-catalyzed cross metathesis can then occur to remove the quencher and produce a fluorescent signal. Using this ArM-based ethylene probe, studies were conducted on a variety of fruit and plant samples to validate its ability to detect both exogenous- and endogenous-induced changes to ethylene biosynthesis. For example, the probe was used to detect changes in ethylene biosynthesis in the outer pericarp of kiwifruit (Fig. 8b), which is typically upregulated during the ripening process.
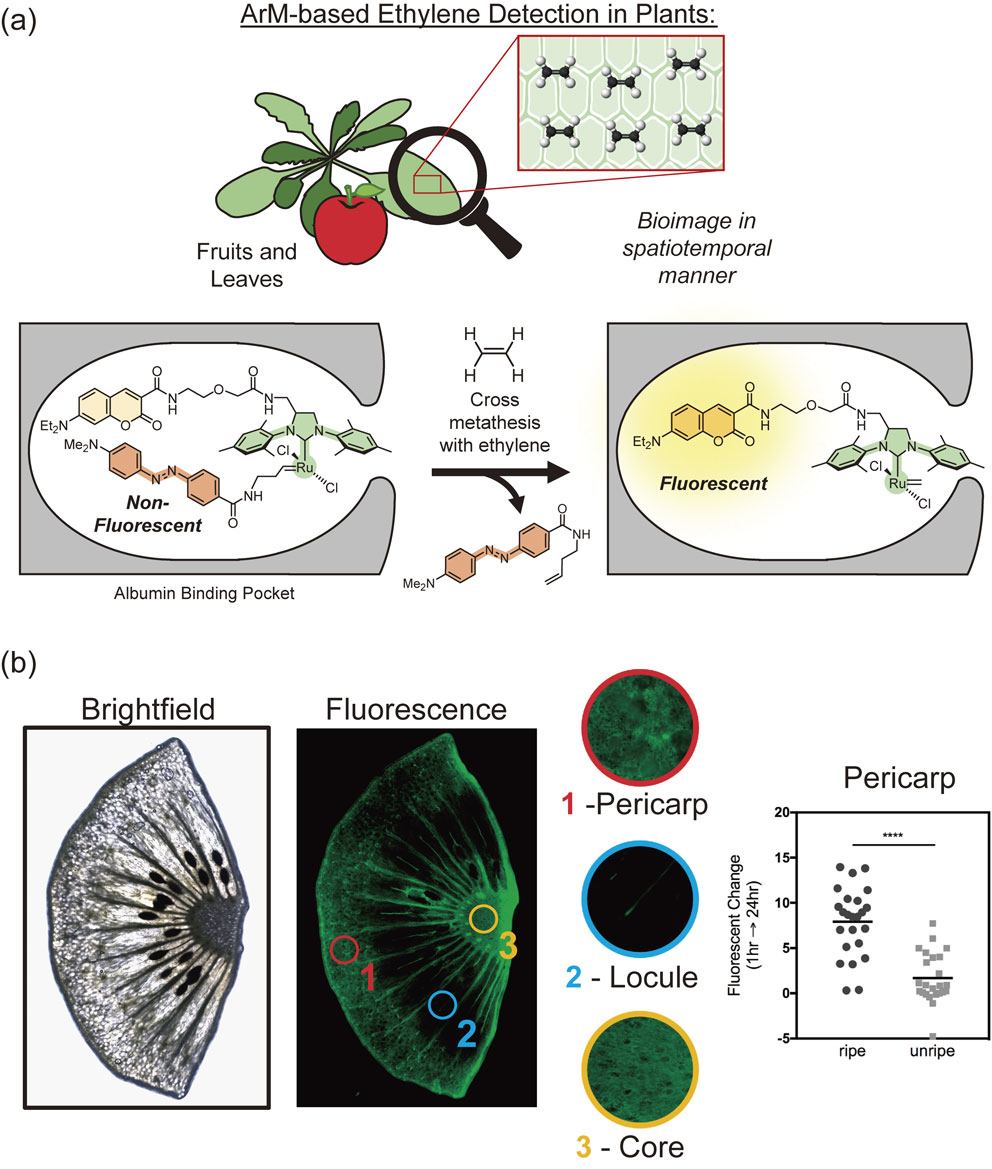
Artificial metalloenzyme-based ethylene probe. (a) The concept behind this approach relies on the presence of ethylene gas to react with a ruthenium catalyst, thereby releasing a quencher to elicit turn-on fluorescence. This probe can be applied directly onto plant tissues to image in a spatiotemporal manner. (b) A cross-section of kiwifruit highlighting the detection of ethylene in different organelles (outer pericarp, locule, core). This technique was subsequently used to monitor the differences in ethylene biosynthesis between unripe and ripening fruits.
Given the bio-orthogonality of abiotic ArMs, there is potential to adapt them towards the development of prodrug therapies. Furthermore, the chemoselectivity of certain metal catalysts carries with them the capacity to access a wider library of potential drug candidates that cannot be accessed through current enzyme technologies. Having said that, similar to the prospects of enzyme therapeutics, clinical viability is largely contingent on their targeted localization to diseased cells/organs. One possible means to achieve this would be through antibody-enzyme conjugates. Coined as antibody-directed enzyme prodrug therapy (ADEPT), the development of this field has been well documented in the literature.107)–109) Despite examples of ArMs constructed from antibody scaffolds,110),111) the use of antibodies to selectively deliver ArMs to targeted cells/organs has unfortunately yet to be investigated.
It should be noted that throughout the development of ADEPT methodologies, one consistent theme has revolved around the issue of protein immunogenicity. In general, enzymes sourced from humans are viewed as undesirable for ADEPT due to the risk of cross-reactivity with naturally expressed enzymes. As a result, bacterial enzymes have become preferred due to their largely different substrate scope. One major drawback of this approach, however, is that proteins of nonhuman origin often suffer from increased immunogenicity. From this perspective, ArMs may solve these problems as human sourced proteins (with no intrinsic catalytic activity) could be converted into an abiotic metalloenzyme. As a consequence, these antibody-ArM conjugates may not only exhibit lower immunogenicity, but also allow for increased bio-orthogonal prodrug activation.
Lectins, also referred to as carbohydrate binding proteins, are naturally expressed proteins that recognize and bind to glycans. Although lectins often bind glycans with relatively weak affinity (i.e., high µM–low mM range), nature circumvents this by relying on the concept of avidity (otherwise known as multivalency), which is grounded upon the combined contributions of several monovalent interactions to elicit strong and selective overall binding. As such, studies have shown that through increasing avidity, lectin-glycan binding can be dramatically increased.112)
As an alternative to antibody-based targeting, a unique opportunity exists to explore lectin-directed, glycan-based targeting (Fig. 9). The primary notion that drives this approach is that certain diseased organs/tumors produce higher levels of specific lectins. For example, cancer cells are known to overexpress lectins that are implicated with processes such as angiogenesis, neoplastic transformation, tumor invasion, and metastasis.113)–116) As such, selective targeting can in theory be obtained by preparing glycoconjugates that match both the identity and spatial arrangement of upregulated lectins of targeted cells.

The concept of lectin-directed, glycan-based targeting is based on the use of a diverse glycan assembly to recognize and bind to lectins (soluble lectins and/or membrane lectins) differentially expressed on cellular surfaces. An advantage of this approach over traditional drug targeting methods is the fact that protein glycosylation is known to increase aspects related to immunogenicity, serum solubility, and several other physical properties.
The biggest advantage of glycan-based methodologies over traditional targeting approaches is the fact that studies have shown glycosylated proteins elicit lower levels of antigenicity and immunogenicity.117) One possible reason for this observation may be due to the effect that protein aggregation has on inducing immunogenicity.118) Because glycosylated proteins often exhibit increased solubility, aggregate formation will likely be reduced. Furthermore, it has also been suggested that the large surface area of attached glycans act to shield the underlying protein sequences from antibody binding. Another beneficial property brought about by protein glycosylation is its increased in vivo stability (i.e., resistance to proteolysis, longer serum half-life). For drug therapy applications, this would thus allow for less frequent dosing.
To pursue glycan-based targeting, our group had previously set out to prepare artificial glycoproteins decorated with a diverse assembly of complex N-glycans.119)–123) Thus, by combining our two areas of research, we went on to establish the concept of glycosylated artificial metalloenzymes (GArMs).
In our first attempt at developing GArM complexes, gold catalyst complexes were anchored into albumin to give GArM-Au. Preferential labeling of organ-specific cells in living mice was thus shown using these GArM-Au complexes (Fig. 10a).103) For example, gold-bound ArMs decorated with α(2,6)sialo-terminated glycans were found to direct preferential labeling to the liver. Confirmed by histological studies, these results also agreed well with previous reports stating that proteins bearing α(2,6)sialo-terminated glycans are generally captured in vivo by the asialoglycoprotein receptor on liver parenchymal cells.124) In the case of ArMs decorated with galactose-terminated glycans, preferential labeling was instead found directed to the intestines. Again, these results matched well with a previous report showing that these glycoprotein complexes accumulate in mouse intestines in vivo (confirmed via dissection studies).119)
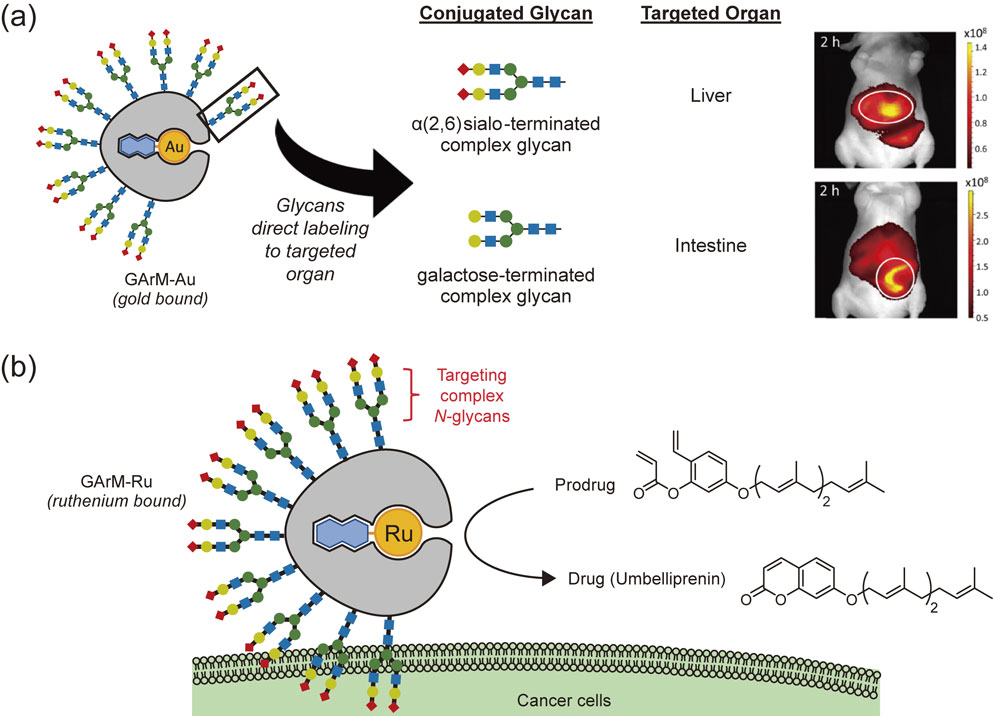
Biofunctionality of glycosylated artificial metalloenzymes (GArMs). (a) By decorating with specific complex N-glycans, GArM-Au can be used to label surface proteins of targeted organs in living mice. (b) Concept of prodrug activation to give the anticancer agent, umbelliprenin, which can occur on the surfaces of targeted cancer cells facilitated by GArM-Ru.
To further add applicability beyond cell labeling, the next attempt by our group aimed at adapting GArM complexes for prodrug activation. Similar to the GArM-Au complexes described above, ruthenium-bound albumin ArMs were decorated with complex N-glycans to give GArM-Ru complexes. In this case, α(2,3)sialo-terminated glycans were chosen as the targeting moiety due to a previous study showing its affinity to certain cancer cells lines.122) In cell-based assays, the observed affinity to SW620, HeLa, and A549 cells is likely due to the upregulated expression of lectins specific for α(2,3)sialo-terminated glycans (i.e., galectin-8,125),126) galectin-1127),128)). Having said that, it should be noted that directly translating the results from cell-based studies into animal-based studies are not always applicable, because many factors from the complex biological environment will have a contributing influence. Nonetheless, we developed our GArM-Ru as a starting point to show its potential for targeted prodrug activation, as depicted in Fig. 10b. Focusing on the anticancer agent umbelliprenin, cell-based studies successfully exhibited targeted cytotoxicity against cultures of HeLa, A549, and SW620 cells.90) For the successful application of our GArM complexes in a therapeutic setting, our group is continuing to engage in not only improving aspects related to glycan-based targeting but also towards developing more robust and active ArMs for the selective conversion of bioactive drugs in vivo.
Looking at the collection of work engaged in developing artificial metalloenzymes, there has been clear progress and evolution in this field over the past decade. In addition to developing biocompatible means to introduce new-to-nature reactions, research groups have also begun to make huge advances in adapting them for applications with important therapeutic and mechanistic implications. In addition, avenues also exist for the development of novel biotechnological tools.
In its current state, however, all of the successful new-to-nature reactions catalyzed by ArMs have been mainly tested in cell-based models with limited therapeutic applications. Despite this, it cannot be denied that the biomimetic approach of ArMs holds great promise for the future once a shift into clinical studies begins.
Looking at biomedical research as a whole, more and more work is likely to be poured into the areas of diagnostics and drug therapy. Given the chemoselectivity of abiotic transition metals (i.e., Ru, Rh, Pd, etc.), researchers will likely look to exploit these properties to not only improve upon current technologies but also possibly towards the development of unexplored methodologies. As such, the field of ArM development should be watched closely as it merges into other traditional and non-traditional research streams.
Edited by Keisuke SUZUKI, M.J.A.
Correspondence should be addressed: K. Tanaka, Biofunctional Synthetic Chemistry Laboratory, RIKEN Cluster for Pioneering Research, 2-1 Hirosawa, Wako, Saitama 351-0198, Japan (e-mail: kotzenori@riken.jp).

Katsunori Tanaka was born in Nara Prefecture in 1973. He received his Ph.D. (2002) from Kwansei Gakuin University, Japan, under the direction of Professor Shigeo Katsumura. After a post-doc with Professors Koji Nakanishi and Nina Berova at Columbia University (2002–2005), he joined Professor Koichi Fukase’s group in Osaka University as an Assistant Professor. He moved to RIKEN as an Associate Chief Scientist in 2012. He was then appointed as a Chief Scientist at RIKEN Cluster for Pioneering Research in 2017 and as a Professor in the Department of Chemical Science and Engineering, Tokyo Institute of Technology in 2019, under the cross-appointment system. He was also appointed as an adjunct Professor in Saitama University (2012), a Professor in Kazan Federal University, Russia (2014), a Group Director of Max Planck- RIKEN Joint Center for Chemical Biology Research (2017), a Deputy Team Leader in GlycoTargeting Research Team, RIKEN (2017), and as an Adjunct Professor in Tokyo Medical and Dental University (2019). His interests include in vivo synthesis, medicinal chemistry, glycochemical biology, and natural products chemistry. He has received a number of domestic and international awards, including a Young Scientists’ Research Award in Natural Product Chemistry (2002; from Japan), The Japanese Society of Carbohydrate Research Award (2010; from Japan), Incentive Award in Synthetic Organic Chemistry (2011; from Japan), ACS Division of Carbohydrate Chemistry, Horace S. Isbell Award (2015; from the USA), Japan Society for the Promotion of Science (JSPS) Prize (2019; from Japan), and The Chemical Society of Japan (CSJ) Award for Creative Work (2020; from Japan).