II. Materials and Methods
Animals
Jic:Sun-Her strain suncus were obtained from CLEA Japan, Inc. (Tokyo, Japan), and BK strain suncus, which are a hybrid of the BAN [14] and KAT [7] strains, were raised in the laboratory animal breeding facility of Okayama Science University.
WKY/NCrlCrlj inbred rats and BALB/cAnNCrlCrlj mice were purchased from Charles River Japan, Inc. (Yokohama, Japan) and rabbits were obtained from Japan SLC, Inc. (Hamamatsu, Japan).
Rats, mice, and suncus were housed in polycarbonate plastic cages containing wood shavings as bedding and were allowed unresticted access to food and water.
The animal experiments in the present study were conducted according to the Guidelines for Laboratory Animal Experiments of the Shigei Medical Research Institute.
Immunization of animals with antigen emulsion
Antigen dissolved in phosphate buffered saline (PBS) and FCA (Difco Laboratories, Detroit, MI) were used for making antigen emulsion. FCA (0.6 ml) and 1.0–1.5 mg/ml antigen solution (0.3 ml) were emulsified in two 1-ml luer lock glass syringes with a double-ended locking needle connector. After emulsification, the glass syringe containing the emulsion was disconnected from the locking connector and a 27G × 3/4” needle was attached.
Animals were anesthetized with sevoflurane (Maruishi Pharmaceutical Co., Osaka, Japan) and injected intramuscularly at both sides of the tail base with emulsion. The amount of emulsion injected was 0.1 ml (100–150 μg of antigen) per rat or suncus, and 0.05 ml (50–75 μg of antigen) per mouse.
Antibodies to suncus IgG
No suncus IgG has been characterized to date and thus it remains unknown whether suncus IgG exists as different isotypes. Furthermore, no commercial antibodies to suncus IgGs are available and thus we produced polyclonal antibodies and mAbs in-house. We have prepared (1) purified suncus IgG as an antigen, (2) polyclonal antibodies and mAbs to suncus IgG, and (3) peroxidase-conjugated and fluorescein isothiocyanate (FITC)-conjugated antibodies for ELISA and immunohistochemistry, respectively.
Purification of suncus IgG from suncus sera
Suncus sera were obtained from Jic:Sun-Her, BK, and KAT strains of suncus with or without antigen immunization. Blood collected at sacrifice was kept for more than 3 hr at room temperature, and the serum was separated from the clot by centrifugation.
Serum from normal suncus was diluted twofold with PBS, ammonium sulfate was added to a concentration of 25 g/100 ml serum with magnetic stirring, and the precipitate formed was obtained by centrifugation. The precipitate was dissolved in PBS and dialyzed against PBS overnight. After the insoluble material was removed by centrifugation, the supernatant was chromatographed on an MEP-gel affinity column (Pall Life Sciences) and the adsorbed fraction was obtained as purified suncus IgG.
Rabbit and rat polyclonal antibodies to suncus IgG
Polyclonal antibodies to suncus IgG were prepared in rabbits and rats. The purified suncus IgG was emulsified with FCA and injected into both sides of the glenoid cavities of the scapula of rabbits and into both sides of the tail base of WKY rats. The same antigen emulsion but with Freund’s incomplete adjuvant was injected as a booster one month after the initial injection.
For rabbit antisera, rabbit blood was collected from the ear vein every week for 2 months. Each rabbit antiserum sample was diluted twofold with PBS and ammonium sulfate was added to a concentration of 25 g/100 ml serum with magnetic stirring. The precipitate formed was collected by centrifugation, then dissolved in PBS and dialyzed against PBS overnight. The insoluble material was removed by centrifugation and the supernatant was chromatographed on a suncus IgG-conjugated gel affinity column. The adsorbed fraction was eluted with 2 M Na2SCN to provide purified antibody to suncus IgG. This antibody was subjected to mouse-serum-conjugated-gel affinity chromatography to remove antibody cross-reactive with mouse IgG. The purified rabbit antibody was conjugated with FITC for immunohistochemistry or with horseradish peroxidase for enzyme-linked immunosorbent assay (ELISA).
To prepare rat antibody to suncus IgG, the immunized WKY rats were injected intraperitoneally with 100 μg of rat nephritogenic mAb b35 to develop proteinuria [11]. The night urine of these rats (from 17:00 to 9:00 the next morning) was collected for 3 weeks using urine-collecting cages 5 days after mAb b35 administration. Each flask used for collecting the urine contained 5 ml of PBS, 0.1% sodium azide, to prevent bacterial growth. Polyclonal antibody to suncus IgG in the urine was precipitated with ammonium sulfate (25 g/100 ml urine) with magnetic stirring and purified as described for rabbit antibody. The purified rat antibody was conjugated with FITC or with peroxidase.
Rat and mouse mAbs to suncus IgG
Mouse and rat mAbs to suncus IgG were prepared using our previously described iliac lymph node method [10, 25]. MEP-gel affinity chromatography-purified suncus IgG with FCA was injected into the tail base of WKY rats or BALB/c mice and the enlarged iliac lymph nodes were collected 17 days later. Cells from the lymph nodes were fused with mouse SP2/0-Ag14 myeloma cells using an electro cell fusion generator (LF201, Nepa Gene Co., Ltd., Chiba, Japan) and plated onto 96-well culture plates (BD Falcon). Positive candidate wells were detected by ELISA with purified suncus IgG as an antigen 9 days after cell fusion. Supernatants from the candidate wells were checked by immunohistochemistry using suncus-IgG-conjugated rat kidney cryostat sections, and by western blot. Fusion cells having desirable characteristics such as clear specificity, strong and reasonable reactivity by ELISA, strong reactivity and a reasonable pattern by western blot, and high cell growth rate, were cloned using the limiting-dilution method. Highly concentrated (1 to 4 mg/ml) antibodies were obtained from a 3-week-period rotation culture (miniPERM® classic bioreactor, MWCO 12.5 kD, 35 ml volume, Sarstedt AG & Co.).
Immunogens used in the present study
1) Preparation of the rat nephritogenic antigen fraction
The procedure used to isolate the rat nephritogenic antigen (NGA) fraction and its related antigenic fractions (i.e., the NC1 fractions of type IV collagen) was similar to that previously reported for the isolation of bovine nephritogenic antigen [21, 22]. Rat renal basement membrane (RBM) was solubilized using collagenase, and then applied to an Ultrogel AcA34 gel filtration column. The main peak fraction (Rat NC1 fraction) was applied to an anti-rat P3 antibody-coupled affinity column. The adsorbed fraction was termed the nephritogenic-antigen (NGA) fraction and the unadsorbed fraction was termed the rat P2 NC1 fraction [22, 24].
2) Rat cryostat kidney sections with suncus IgG on the GBM
Rat kidneys exhibiting suncus IgG binding and renal basement membranes were prepared to detect the specificity and staining strength of the rat antibodies to suncus IgG. The rat nephritogenic antigen fraction was injected into the tail base of 2 male BK suncus and antiserum was obtained from each suncus 3 weeks later. Antiserum (1 ml) was injected into a WKY rat by intraperitoneal injection. Rat kidneys exhibiting suncus IgG binding along the GBM in a linear pattern were collected 1 day after injection and frozen with liquid nitrogen. Cryostat sections of these kidneys were used to detect antibodies to suncus IgG for immunohistochemistry.
Generation of suncus immortalized lymphoid cell lines and fusion partner cell lines
1) Generation of suncus immortalized lymphoid cell lines
Male and female suncus were immunized by injection intramuscularly at the tail base with an emulsion containing FCA and rabbit IgG or the rat NGA fraction. The animals were sacrificed 17 days later and the enlarged paraaortic lymph nodes, sacral lymph nodes, and the spleen were collected. The lymph nodes and spleen of each animal were sieved through a #200 stainless steel mesh and the cells were cultured in a 96-well tissue culture plate (BD Falcon) to obtain immortalized lymphoid cell lines. The culture medium was GIT medium (Wako Pure Chemical, Osaka, Japan) containing 10% fetal bovine serum.
2) Generation of hypoxanthine-aminopterin-thymidine (HAT)-sensitive cell lines
To obtain a HAT-sensitive fusion partner, cell lines were cultured in the presence of 8-azaguanine (Sigma-Aldrich). The concentration of 8-azaguanine in the culture medium was initially 2.0 μg/ml and was increased to 5, 10, and finally 20 μg/ml. Cells increasing in the culture medium containing 20 μg/ml 8-azaguanine were sensitive to HAT medium.
3) Generation of suncus fusion partner cell lines
Immortalized cells and lymph-node lymphocytes were fused using an electro fusion method. Fusion partner cells and lymph-node cells (2–4 × 107) from immunized suncus were fused at a ratio of 1:2 at room temperature using an electro cell fusion generator. The cells were stirred in HAT medium and plated onto 96-well tissue culture plates. Medium was changed at day 7 to hypoxanthine-thymidine (HT) medium and at day 12–14 to GIT medium containing 10% fetal bovine serum. Supernatants were tested for the presence of IgG or for immunogen-specific antibody at day 14–16 using ELISA.
Methods used to characterize the mAbs
1) Determination of protein concentration
The absorbance at 280 nm was used to estimate protein concentration using an extinction coefficient of 1.35 for l.0 mg immunoglobulin and 1.10 for 1.0 mg of rat soluble fraction from renal basement membranes.
2) Rat mAbs used to characterize suncus mAbs
The rat mAbs used were mAb 114 (IgG1, κ) [11], mAb H12 (IgG1, λ) [23], mAb a84 (IgG2a, κ) [11], mAb b35 (IgG2b, κ) [11], and mAb H24 (IgG2c, λ) (established at the same time as mAb H25) [8].
3) ELISA
ELISA was performed in 96-well immuno plates (F96 Maxisorp Nunc Immuno Plates, Thermo Scientific) as previously described [17, 21]. The wells of the ELISA plate were coated with 150 ng/50 μl antigen for 1 hr and blocked with 1% bovine serum albumin for 1 hr. Horseradish peroxidase-conjugated anti-rat immunoglobulins and anti-rat IgG isotypes were purchased from Dako A/S (Glostrup, Denmark) and Binding Site (Birmingham, UK), respectively.
4) Western blots
The IgG fraction was dissolved in sample buffer (pH 6.8) containing 4.4% SDS and 50 mM DTT and boiled for 5 min. SDS-polyacrylamide gel electrophoresis (SDS-PAGE) was performed using 5–20% gradient polyacrylamide gels.
Proteins were transferred from the electrophoresed gels to PVDF membranes (Nihon Millipore, Yonezawa, Japan), washed with 10 mM Tris-HCl buffer (pH 7.4) containing 0.9% NaCl and 0.05% Tween 20 (TBS-T), and then blocked with 5% skim milk in TBS-T. After washing with TBS-T, each membrane was incubated with a given mAb for 60 min and then washed with TBS-T. The mAbs bound to the membranes were detected by incubation for 1 hr at room temperature with peroxidase-conjugated secondary antibody diluted with TBS-T. After washing, peroxidase bound to the sections was detected using diaminobenzidine tablet reagent (Wako Pure Chemical, Osaka, Japan).
5) Indirect immunofluorescence
Frozen kidneys were sectioned into 4 μm-thick slices using a cryostat, and then the sections were stained with primary antibody (typically a culture supernatant containing mAbs) for 1 hr. The sections were washed with PBS and stained for 30 min to 1 hr with the secondary antibody, typically FITC-conjugated polyclonal antibody or mAb at an appropriate concentration. After washing with PBS, the sections were mounted in PermaFluor aqueous mounting medium (Thermo Scientific) or 10%-glycerin PBS and observed with a fluorescence microscope.
6) Direct immunofluorescence
Frozen kidneys were sectioned into 4 μm-thick slices using a cryostat, and then stained with an FITC- and/or Alexa555-conjugated antibody for 30 min to 1 hr. After washing with PBS, the sections were mounted in PermaFluor or 10%-glycerin PBS and observed with a fluorescence microscope.
7) Conjugation of FITC, Alexa 555, or horseradish peroxidase to polyclonal antibodies and mAbs
mAbs obtained from rotation culture were purified by MEP-gel affinity-column chromatography. Each antibody was then conjugated with FITC (Sigma-Aldrich), Alexa 555 (Thermo Scientific), or horseradish peroxidase (Sigma-Aldrich) according to the instructions included with the Alexa 555 reagent.
Rabbit or rat polyclonal antibody to suncus IgG was purified by ammonium sulfate precipitation (25% saturation) followed by suncus-IgG-fraction-conjugated Sepharose CL-4B affinity column chromatography. The purified antibody was conjugated with FITC or horseradish peroxidase according to the instructions supplied by the reagent manufacturer (Thermo Scientific).
Cell fusion with a fusion partner
Eight- to 20-week-old male BK-strain suncus were injected intramuscularly at both sides of the tail base with emulsion (0.1 ml) containing 100–150 μg of antigen fraction. The enlarged paraaortic and sacral lymph nodes were collected 17 days after the injection. Cells from the lymph nodes were fused with SILC cells, SIPH7A cells, or other candidate cells using an electro cell fusion generator and plated onto four 96-well culture plates. No feeder cells were used for cell culture. The HAT culture medium (150 μl/well) was replaced with fresh HAT medium 7–8 days after cell fusion and with HT medium 14 days after cell fusion. Positive candidate wells were detected by ELISA using immunized antigen as an adsorbed antigen 17 days after cell fusion.
III. Results and Discussion
Characterization of mouse mAbs to suncus IgG
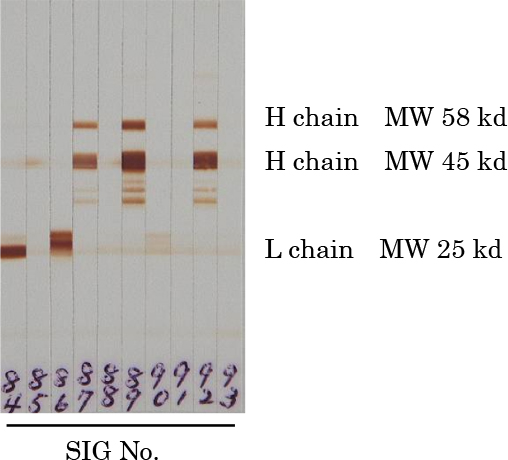
Thirty-three murine clones were established for the detection of suncus IgG. These clones stained rat kidney-bound suncus IgG by indirect immunofluorescence. Fifteen clones recognized suncus IgG by western blot (Table 1): 7 recognized the heavy chain of suncus IgG and 8 recognized the light chains. Two different patterns were detected in the light chains, were tentatively termed Lc1 chain and Lc2 chain (Fig. 1), and corresponded to the kappa or lambda chains of IgG from other mammals.
Table 1.
Characterization of mouse SIG antibodies testing positive in western blot analysis
| SIG No. |
WB pattern |
WB intensity |
Mouse isotype |
Immunohistochemistry |
Rotation culture |
HRP conjugation |
| 62 |
γ chain |
++ |
IgG1 |
++ |
done |
|
| 66 |
γ chain |
++ |
IgG1 |
+ |
done |
|
| 70 |
γ chain |
++ |
IgG1 |
+ |
done |
|
| 76 |
γ chain |
++ |
IgG1 |
++ |
done |
done |
| 87 |
γ chain |
++ |
IgG1 |
++ |
done |
|
| 89 |
γ chain |
++ |
IgG1 |
++ |
|
|
| 92 |
γ chain |
++ |
IgG1 |
++ |
|
|
| 63 |
Lc1 chain |
++ |
IgG2a |
−/+ |
|
|
| 74 |
Lc1 chain |
++ |
IgG1 |
++ |
|
|
| 77 |
Lc1 chain |
++ |
IgG1 |
+ |
|
|
| 80 |
Lc1 chain |
++ |
IgG1 |
++ |
done |
done |
| 81 |
Lc1 chain |
++ |
IgG1 |
++ |
|
|
| 84 |
Lc1 chain |
++ |
IgG2a |
++ |
done |
|
| 64 |
Lc2 chain |
++ |
IgG1 |
++ |
done |
|
| 86 |
Lc2 chain |
++ |
IgG1 |
++ |
done |
done |
These established mouse antibodies were named SIG antibodies and identified by numbers between 61 and 93. Figure 2 shows the specificity of mouse SIG61-SIG90 antibodies as determined by ELISA. Three candidate antibodies were conjugated with peroxidase (SIG76 to the heavy chain of suncus IgG, SIG80 to the Lc1 light chain, and SIG86 to the Lc2 light chain) for ELISA and western blots.
Characterization of rat mAbs to suncus IgG
Forty-three clones for the detection of suncus IgG were established using rats as the mAb-producing animals, of which 21 were positive in western blot and showed conjugation to the heavy chain of suncus IgG. These rat antibodies were named SIG antibodies and were identified with numbers between 21 and 53. SIG23 and SIG47 were purified from the supernatants of rotation cultures and used for sandwich ELISA as capture antibodies.
Development of suncus fusion partner cells
Figure 3 shows the derivation of suncus plasmacytomas, suncus immortalized lymphoid cells, and hybridomas established for producing suncus mAbs.
1) Production and identification of suncus immortalized lymphoid cells by culture
In January and February 2009 we identified proliferating lymphoid cells in culture wells about one week after plating cells from the enlarged lymph nodes of Jic:Sun-Her suncus immunized with emulsion containing FCA and rabbit IgG or rat NGA fraction. The culture was contaminated with mycoplasma-like cells (Fig. 4). The culture medium did not become turbid during culture and the proliferating cells looked normal even as the contaminating microorganism proliferated [19]. We believed the contamination was mycoplasma because mycoplasma strains were previously isolated from house musk shrews [6]. Cloning was repeated in the presence of anti-mycoplasma reagent in the culture medium and cloned cells without contamination were obtained and cell lines were established. In the 7-year period since establishing these clones, we have conducted cell fusions using lymph node cells from BK, KAT and Jic:Sun-Her strain suncus immunized with antigens and have cultured lymph node cells not used for cell fusion. During this time we never again observed the proliferation of lymphoid cells.
Four immortalized lymphoid cell lines (SILC cells) were established from 3 Jic:Sun-Her suncus immunized with emulsion containing FCA and rabbit IgG or rat NGA fraction (Fig. 3).
Two SILC cell lines, SILC1 and SILC2, were obtained independently from the enlarged lymph nodes of two Jic:Sun-Her suncus immunized with rabbit IgG. Lines SILC3 and SILC4 (Fig. 5a) were obtained from the spleen and the lymph nodes of a Jic:Sun-Her suncus immunized with rat NGA fraction. The culture supernatants from the SILC1 to SILC4 cells were tested by sandwich ELISA using mouse or rat mAbs to suncus IgG as a capture antibody and showed that none of these cell lines produced antibodies. The doubling time of these cells was about 20 hr.
Despite the lack of immunoglobulin production, we believed these immortalized cells were plasmacytoma because both the cells and the colonies closely resembled mouse SP2/0-Ag14 myeloma cells [26] used for the production of our mouse and rat mAbs (Fig. 5b). In addition, following cell fusion with B lymphocytes from enlarged lymph node, several fused hybrid cells began secreting suncus IgG.
Karyotype analysis of the SILC2 and SILC4 cells showed that they contained 66 to 70 and 71 to 74 chromosomes (data not shown), respectively, suggesting these were not normal cells. Normal suncus cells contain 40 chromosomes [1] and thus SILC2 and SILC4 are aneuploid tetraploid cells.
2) Induction of IgG-secretion by SILC4-Ag1
Eight HAT-sensitive clones, SILC2-Ag1 to SILC2-Ag4 and SILC4-Ag1 to SILC4-Ag4, were established from the original HAT-resistant clones by selection with 8-azaguanine.
In September 2010 we fused SILC4-Ag1 and lymph node cells from the enlarged paraaortic and sacral lymph nodes of BK suncus immunized with KLH and obtained suncus immunoglobulin-producing hybrid cells (SIPH cells). Sixteen IgG-positive wells were identified from 768 wells by ELISA, of which 6 SIPH cells were cloned using the limited dilution method. Two-week-period rotation cultures of these 6 clones were conducted to determine their ability to stably secret IgG.
The culture medium IgG concentrations were: SIPH1, 1.61 mg/ml; SIPH4, 0.04; SIPH5, 0.08; SIPH7, 0.35; SIPH8, 0.03; and SIPH16, 0.56. These cells secreted monoclonal IgG molecules but none of these IgGs was an antibody to the immunization antigen, KLH. We believe these cells could be used as cell fusion partners if they were HAT-sensitive and with the exception of SIPH7, all the SIPH cell lines were transformed to HAT-sensitive line using 8-azaguanine. The HAT-sensitive cells were named SIPH1-Ag to SIPH16-Ag cells. The original SIPH7 cells were already HAT-sensitive.
The light chains of these IgG molecules were determined. The IgG molecules secreted from SIPH4, SIPH5, and SIPH7 cells had the Lc1 light chain, and those secreted from SIPH1, SIPH8, and SIPH16 cells had the Lc2 light chain.
The SIG antibodies from rats and mice were characterized by performing ELISA with the monoclonal IgG molecules produced by the SIPH1, SIPH5, SIPH7, and SIPH16 cells. The SIG antibodies from rats were reactive with all the monoclonal IgG molecules, as were all the mouse SIG antibodies except for the 8 antibodies to the light chains, which were only reactive with the corresponding monoclonal IgG molecules (Fig. 2). There was no evidence of different isotypes of the suncus heavy chain of IgG.
These results clearly indicate that SILC4-Ag1 cells worked as a fusion partner and could produce suncus mAb following cell fusion, although the mAbs were not immunogen-specific. Furthermore, the results indicate that the SILC cells are plasmacytoma.
3) Production of immunogen-specific mAbs with SIPH cells
Preliminary experiments were conducted to determine whether the SIPH cells could produce immunogen-specific antibodies. Cell fusions were generated using HAT-sensitive SIPH1-Ag, SIPH4-Ag, SIPH5-Ag, SIPH7A, SIPH8-Ag, or SIPH16-Ag cells as the partner cells and lymph node lymphocytes from BK suncus immunized with rat P2 NC1 fraction, human serum albumin (HSA), chicken ovalbumin (OVA), or KLH. The production of immunogen-specific antibodies was confirmed by ELISA (day 7 to day 14 after cell fusion) in all fusions, but the ability to produce these antibodies was generally lost during cloning, suggesting that the fused cells were very unstable. However, one clone to rat basement membrane antigen (STF20), one clone to HSA, and 5 clones to OVA (SOV1-SOV5) were stably maintained and these cloned cells were re-cloned and more stable cell lines were established (Fig. 3).
It seems that the higher the concentration of IgG, the higher the productivity of IgG by the cell line and thus the higher the stability of the cell line. SIPH1, SIPH7, and SIPH16 were chosen as further fusion partner cell candidates. The doubling times of SIPH1-Ag, SIPH7A and SIPH16-Ag were about 24, 20, and 20 hr, respectively.
4) Generation of SIPH7A2 partner cells
SIPH7A cells successively cultured in culture plates for 12 weeks were re-cloned using four 96-well culture plates. Only eight positive wells were identified by ELISA at day 9. Since a total of 324 colonies were observed, the positive colonies represent about 2.5% of the total colonies. These positive cells were considered likely to be more stable and thus were selected as new fusion partners.
Multiple cell colonies were found in the positive wells and were re-cloned again. Five cell lines (SIPH7A2, SIPH7A4, SIPH7A5, SIPH7A6, and SIPH7A8) were established from 5 different wells. We confirmed that these cell lines could be used as partner cells and one line, SIPH7A2, was studied further. We propagated the SIPH7A2 cells (Fig. 5c), and then divided the cells and stored them in 24 freezing tubes as the same lot of cells. For each fusion experiment, frozen cells were thawed and cultured to the cell number required for cell fusion.
5) Production of immunogen-specific mAbs with SIPH7A2 and generation of SIPH75-7Ag3 cells
Cell fusion with SIPH7A2 and lymphocytes from suncus injected with HSA (Sigma-Aldrich) provided 3 clones producing suncus mAbs to HSA and one clone producing an unidentified suncus IgG. Because the clone producing unidentified suncus IgG was rather stable for the production of IgG, re-cloning was performed using a 96-well plastic culture plate. Seven wells each contained one well-grown cell colony, of which 6 produced suncus IgG. These antibody-positive cell lines were named SIPH75-1 to SIPH75-6 and the antibody-negative cell line was named SIPH75-7. SIPH75-7 cells were treated with 8-azaguanine to obtain HAT-sensitive cell lines. One of the HAT-sensitive cell lines which did not secrete suncus IgG was named SIPH75-7Ag3 (Fig. 5d). SIPH75-7Ag3 was propagated, divided into 24 cryostat tubes, and stored for further cell fusions for the generation of immunogen-specific suncus antibodies.
6) Production of immunogen-specific mAbs with SIPH75-7Ag3 cells
6-1-1) Production of Suncus mAbs to rat IgG
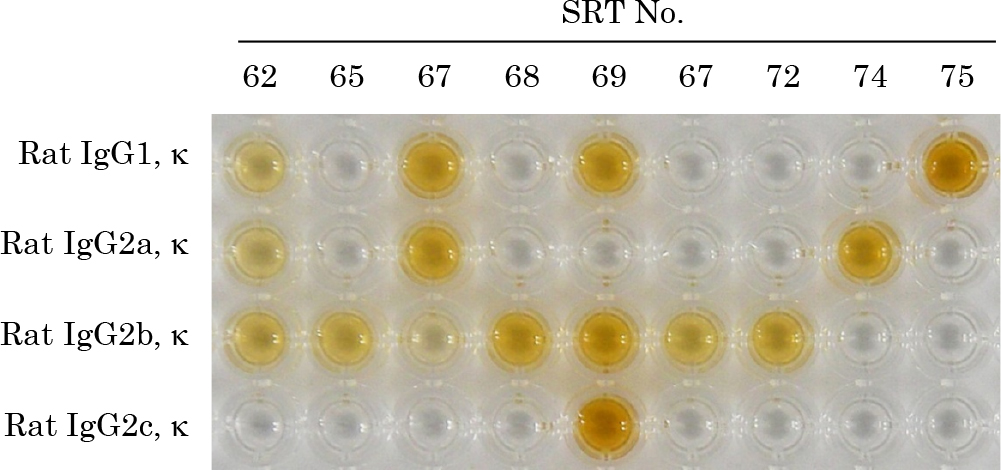
The enlarged paraaortic and sacral lymph nodes from male BK strain suncus immunized with rat IgG were collected 17 days after immunization. Three separate fusions were performed. Cells from the lymph nodes were fused with SIPH75-7Ag3 cells using an electro cell fusion generator and plated in four 96-well culture plates. HAT culture medium was replaced with fresh HAT medium 7 days after cell fusion. In one of the cell fusions, more than 40 positive candidate wells were detected by ELISA using rat IgG as an adsorbed antigen 14 days after cell fusion (Fig. 6).
The supernatant of each positive well identified by ELISA was checked to determine the specificity and strength of the antibody by staining rat cryostat renal sections whose GBMs were previously bound with a mixture of three isotypes of rat mAbs: mAb 114 (IgG1), mAb a84 (IgG2a), and mAb b35 (IgG2b) [11]. When the supernatant of a well stained the rat IgG on the GBM strongly in a linear pattern, the well was selected as a candidate.
Positive clones were frequently lost following the detection of specific antibodies by ELISA and by immunohistochemistry using the supernatant from the well, suggesting that the fused cells were unstable. To increase the efficiency of cloning, the culture medium of wells containing almost confluent cells was changed in the evening, and the next morning the culture medium was assayed for the production of antibody. Cloning was performed when adequate activity was observed. Re-cloning was performed at least twice, and cells were proliferated by rotation culture in a miniPERM® for 3 weeks. Next, an aliquot of each culture was collected and cultured in a 24-well plastic culture plate to confirm the presence of antibody-producing cells.
In this study we established 9 SRT clones of suncus mAbs with different specificities to rat IgG isotypes (Figs. 3 and 7). Table 2 shows the detailed specificity and staining strength of the 9 mAbs: one was specific for light chain κ (SRT62), one for IgG1 isotype (SRT75), one for IgG2a isotype (SRT74), four for IgG2b isotype (SRT65, SRT68, SRT70, and SRT72), and the other two clones recognized multiple rat IgG isotypes (SRT67 and SRT69).
Table 2.
Specificity of 9 immunogen-specific suncus mAbs to rat IgG
| SRT No. |
Reaction with rat IgG isotypes |
| Rat IgG1, κ |
Rat IgG1, λ |
Rat IgG2a, κ |
Rat IgG2b, κ |
Rat IgG2c, λ |
| 62 |
++ |
− |
++ |
++ |
− |
| 65 |
− |
− |
− |
++ |
− |
| 67 |
++ |
++ |
++ |
+ |
− |
| 68 |
− |
− |
− |
++ |
− |
| 69 |
++ |
++ |
− ~ ± |
++ |
++ |
| 70 |
− |
− |
− |
++ |
− |
| 72 |
− |
− |
− |
++ |
− |
| 74 |
− |
− |
++ |
− |
− |
| 75 |
++ |
++ |
− |
− |
− |
Specificity was assayed by ELISA: plates were coated with rat immunoglobulin of the indicated isotypes (all obtained from our previous studies).
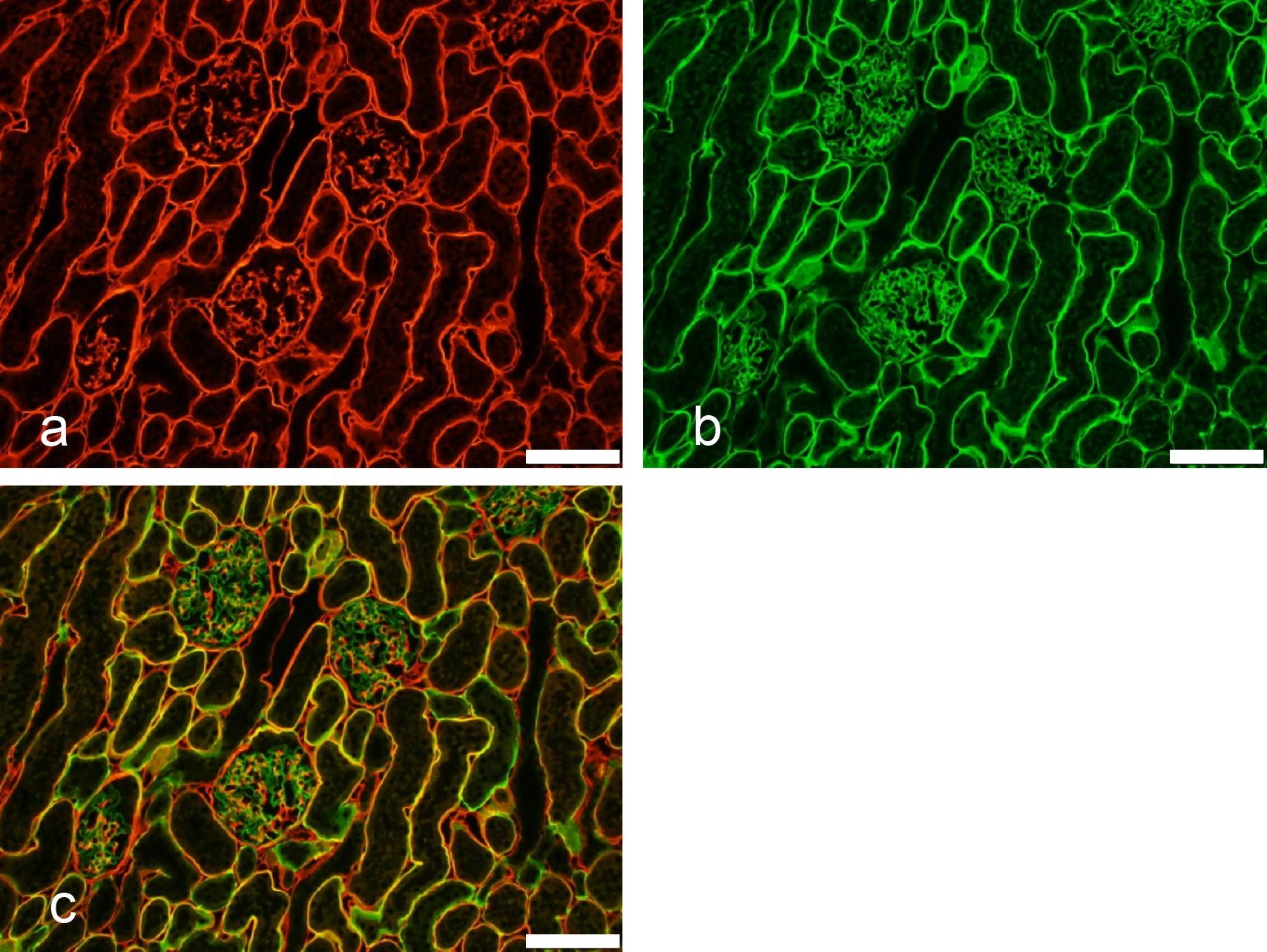
Suncus mAb SRT75 to rat IgG1 was conjugated with Alexa 555 and mAb SRT65 to rat IgG2b was conjugated with FITC. The mixture of the two fluorochrome-conjugated suncus mAbs was used as the secondary antibodies (Fig. 8) for indirect immunofluorescent antibody staining of a rat kidney section. The rat kidney section was first acid-treated, washed with PBS, and then reacted with a mixture of rat mAbs as the primary antibodies: mAb H12 (rat IgG1) to rat α1(IV)NC1 domain of type IV collagen [23] and mAb b35 (rat IgG2b) to rat α4(IV)NC1 domain of type IV collagen [11]. Figure 8 clearly shows the presence of each domain as expected, indicating that the use of suncus mAb in immunofluorescent staining as the secondary antibody is both practical and very useful.
6-2) Production of suncus mAbs to rabbit IgG
Two separate fusions were performed using the SIPH75-7Ag3 cell line and lymph node cells from suncus immunized with rabbit IgG. Seven clones of suncus mAbs to rabbit IgG were established (SRB21, SRB22, SRB23, SRB24, SRB25, SRB27, and SRB28). One clone (SRB27) was cultured in rotation culture and the concentrated antibody obtained was purified using MEP-gel affinity chromatography, and then conjugated with FITC. The conjugated antibody could be used in the same manner as polyclonal goat FITC-conjugated to rabbit IgG.
The cross-reactivity of these 7 SRB mAbs with rat IgG1, IgG2a, IgG2b, and IgG2c was checked by ELISA. mAb SRB23 cross-reacted with rat IgG2c and mAb SRB27 cross-reacted slightly with rat IgG2c. The other 5 SRB antibodies did not cross-react with rat IgG isotypes.
7) Attempt to induce other SIPH cells via cell fusion
By May 2013 we had generated partner cell lines such as SIPH7A2 and SIPH75-7Ag3, but sought to obtain SIPH cells displaying better characteristics. We used SILC1 cells and SILC4-Ag2 cells for this purpose. We first established a HAT-sensitive SILC1-Ag cell line from SILC1 cells using 8-azagunanine. SILC1-Ag cells and SILC4-Ag2 cells were fused with lymph node cells from BK suncus immunized with a rat NC1 fraction as an antigen (Fig. 3). Eighteen IgG-positive wells were found using ELISA, and 4 new SIPH cell lines (SIPH46, SIPH47, SIPH48, and SIPH49) were established using the limited-dilution method (Fig. 3). Only SIPH48-producing mAb was found to be immunogen-specific. Because its staining pattern of rat kidney was the same as that of mAbs to rat α3/α4/α5(IV)NC1 [23], it was named mAb STF31.
These results clearly indicate that SILC1-Ag cells and SILC4-Ag2 cells function as a fusion partner and can produce immunogen-specific suncus mAb, although the production of immunogen-specific mAbs was very inefficient. These results further indicate that the SILC cells are plasmacytoma.
We attempted to generate fusions of SIPH46-Ag and SIPH47-Ag cells and lymph node cells from suncus immunized with rabbit IgG. Three new mAbs to rabbit IgG (SRB51, SRB52, and SRB53) were obtained but these fusions were not pursued because the growth rates of the SIPH46-Ag, SIPH47-Ag, SIPH48, and SIPH49 cells were slower (doubling times of 24 hr or more) than expected.
8) Production of antigen-specific mAbs with SIPH7A2G25 cells
We knew that SIPH cells change during continuous culture and believed that positive cells selected after extended culture would be more stable.
We used (2–5) × 103 cells suspended in 24 ml culture medium and plated onto a 24-well culture plate. After the cells were almost confluent, they were collected and (2–5) × 103 cells were plated on a new culture plate for the next subculture. We continued the process of subculturing and subcloning the cells every 6–8 weeks and selected several clones exhibiting higher antibody production and faster cell growth. We mixed cells from 4–6 wells, generated a new cell line, and continued the cell culture process.
The obtained cell line, SIPH7A2G24, had been cultured for about 120 days and subcloned twice. It was again subcloned using four 96-well culture plates 53 days after the last cloning to obtain antibody-positive cells. The number of wells with one cell colony was counted. Approximately 60% of the one-cell colony wells were positive, indicating that the SIPH7A2G24 cell line was still changing. We mixed cells from 4 antibody-positive wells to generate a new cell line, SIPH7A2G25.
9) Production of immunogen-specific mAbs with SIPH7A2G25 cells
Three separate fusions were performed using the SIPH7A2G25 cell line and lymph node cells from suncus immunized with rabbit IgG. Nine clones (SRB29, SRB30, SRB31, SRB32, SRB33, SRB35, SRB37, SRB38, and SRB39) were obtained (Fig. 3), of which SRB35 and SRB36 were further cultured in rotation culture. The obtained concentrated antibodies were purified using MEP-gel affinity chromatography, conjugated with FITC, and used for staining rabbit IgG (Fig. 9). The conjugated antibody could be used in the same manner as polyclonal goat FITC-conjugated to rabbit IgG.
The cross-reactivity of these 9 SRB antibodies and 3 other SRB antibodies (SRB51, SRB52, and SRB53) with rat IgG1, IgG2a, IgG2b, and IgG2c were tested using ELISA. The rat mAbs used were mAb 114 (IgG1, κ), mAb a84 (IgG2a, κ), mAb b35 (IgG2b, κ), and mAb H24 (IgG2c, λ). None of these 12 SRB antibodies cross-reacted with rat IgG isotypes.
The use of SIPH7A2G25 cells as a fusion partner resulted in many wells containing fusion cells that produced immunogen-specific antibodies. Many of these fusion cells were unstable and rapidly lost their antibody-producing activity, but several fusion cells stabilized and continued producing antibodies. The detection of sufficient activity by ELISA in the culture media at day 18–19 (just before cloning) was indicative of a high probability of successful cloning.
Yield of suncus mAbs in rotation culture
Established cell lines producing a suncus mAb were cultured in rotation culture to obtain a highly concentrated antibody and to confirm the presence of antibody-producing cells even after 3 weeks of continuous culture. The concentration of rat and mouse mAbs following 2–3 weeks of rotation culture is usually 1 to 4 mg/ml culture medium, whereas the concentration of suncus mAbs following 3 weeks of rotation culture was usually 0.3 to 1 mg/ml culture medium. Despite the low yield of suncus antibody in rotation culture, it could be purified in the same manner as rodent antibodies using MEP-gel affinity chromatography. The conjugation of the purified antibody with FITC and the usability of the conjugated antibody for direct immunohistochemistry were the same as for rodent antibodies.
Suncus fusion partner cells and the lymph node method
Non-human fusion partner cells for the production of mAbs have been established only in four species: mice [12, 26], rats [2, 4, 9, 15], rabbits [27, 28], and suncus (the present study). Mouse-mouse homohybridomas and mouse-rat heterohybridomas using SP2/0-Ag14 as a fusion partner are very stable, but mouse-rabbit heterohybridomas [18] were very unstable, suggesting that partner cells of the same species or genetically close species are necessary. Compared to the homohybridomas generated using SP2 partner cells, the other non-mouse homohybridomas constructed to date for producing mAbs are unstable, although their ability to produce mAbs can be maintained by repeated subcloning and by selecting stable cells with robust growth and high antibody production.
All suncus hybridomas established in the present study, such as STF20, STF31, SRB27, SRB35, and SRB36, were cultured in rotation culture for 3 weeks and culture media containing 0.3 to 1 mg/ml antibody were obtained. Despite the current low efficiency of positive candidate cell production by cell fusion, target antibodies were obtained. The lymph node method using enlarged paraaortic and sacral lymph nodes as a source of sensitized B lymphocytes is highly advantageous with suncus.
Suncus mAbs to rat and mouse basement membrane antigens
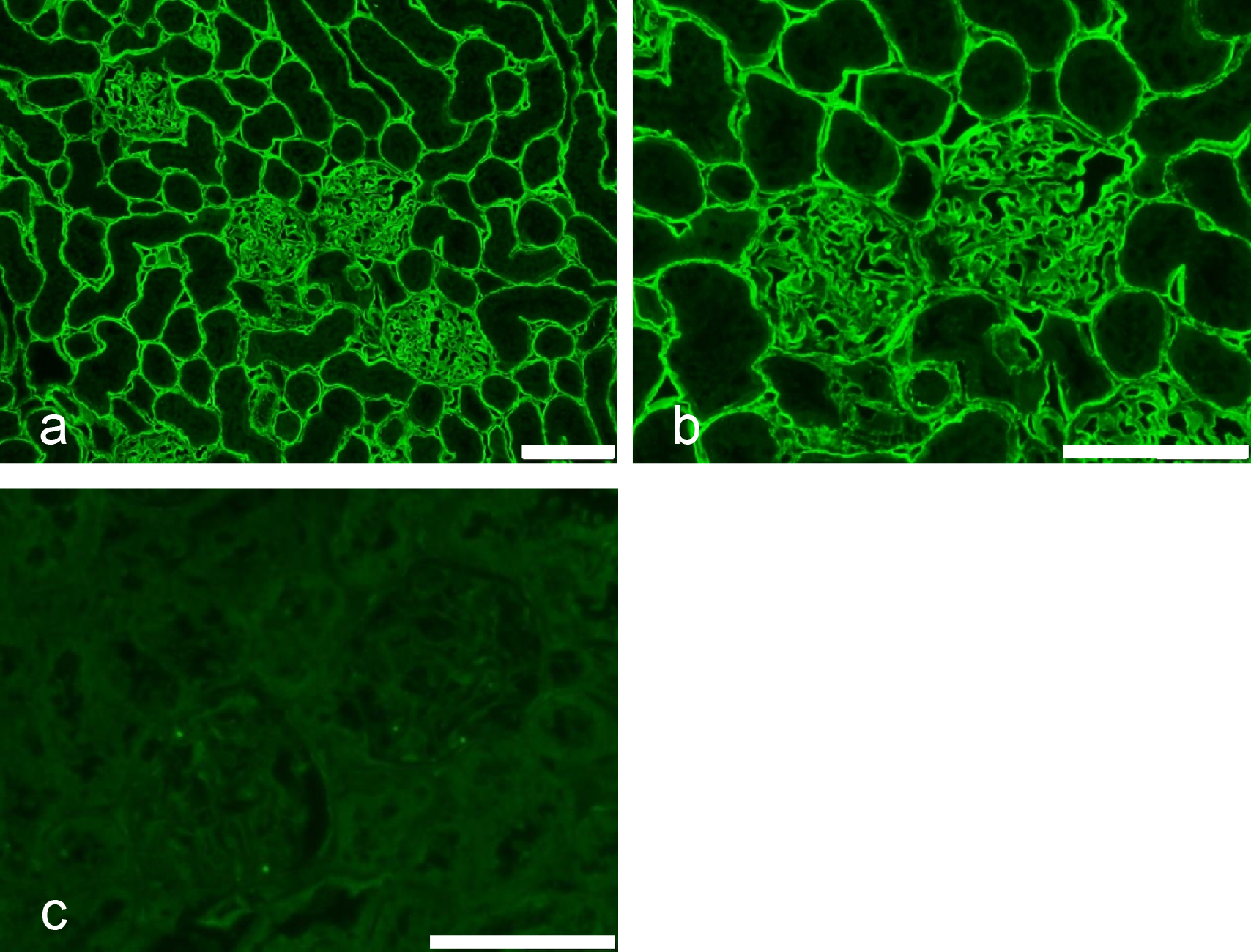
Importantly for mAb production, suncus are insectivores, genetically distant to rodents, and they recognize antigens and epitopes that are not immunogenic in rats or mice. For example, the amino acid sequences of type IV collagen alpha 1 and 2 chains are very similar in rat and mouse: the sequences of α1(IV)NC1 are identical, and the sequences of α2(IV)NC1 differ by only two amino acids. No mAbs reactive with native rat or mouse type IV collagen have been reported, although there are several rat and mouse mAbs that react with denatured proteins [23]. In the present study, mAb STF20 was stably obtained. STF20 cells were cultured in rotation culture, and then the mAb STF20 was purified using MEP-gel affinity chromatography. The purified mAb STF20 was conjugated with Alexa 555. The immunofluorescence pattern using a rat kidney cryostat section is shown in Figure 10 and is the same staining pattern as that of mAb H12 shown in Figure 8a [23].

To confirm the specificity of mAb STF20, sandwich ELISA using affinity-purified mAb b14, which reacts with α5(IV)NC1 [11] as capture antibody, rat NC1 fraction as target antigen, mAb STF20 as detection antibody, and peroxidase-labeled anti-suncus IgG as secondary antibody, was conducted. In this sandwich ELISA the capture antibody, mAb b14, captured the NC1 hetero-hexamers containing the α1/α1/α2(IV)NC1 and α3/α4/α5(IV)NC1 domains of type IV collagen [3]. A positive result was obtained in the sandwich ELISA, indicating that mAb STF20 reacted with rat NC1-hexamer of type IV collagen. The staining pattern of mAb STF20 was an α1/α1/α2(IV)NC1-domain-staining pattern, indicating that mAb STF20 reacts with the α1/α1/α2(IV)NC1 domain of NC1 hetero-hexamers. mAb STF20 is the first mAb shown to react with rat native protein of the α1/α1/α2(IV)NC1 domain of type IV collagen.
A SIPH48-producing mAb, mAb STF31, stained rat renal basement in the same pattern as rat mAbs to the α3/α4/α5(IV)NC1 domain of type IV collagen (Fig. 8b). Many rat mAbs to the native protein of rat α3/α4/α5(IV)NC1 domain have been obtained because the rat protein is an autoantigen and is immunogenic to rats [11].
Production of suncus mAbs in the future
Suncus have been used in medical research for over 40 years [13, 16], for example, to investigate emesis [20] and gastrointestinal hormones [5]. There are several disadvantages to using suncus, such as the lack of inbred stains, specific pathogen-free strains, limited availability from commercial sources, little genetic data, and limited published literature.
Even if fusion partner cells and FITC- and peroxidase-conjugated antibodies to suncus IgG were commercially available, difficulties producing mAbs with suncus will remain, including the availability of the animals and their rearing over a period of several weeks. One solution would be to immunize suncus with antigens in a specialized facility and to collect the lymph node cells 2–3 weeks after immunization. Upon collection, the cells would be sieved through a stainless steel mesh, kept frozen, and then transferred to another research facility for fusion.
The current efficiency of producing suncus-suncus hybridomas which secrete mAbs is much lower than that of rat-mouse or mouse-mouse hybridomas obtained using mouse SP2 partner cells. We anticipate that, just as rabbit partner cells have been improved [28], better suncus partner cells will be generated in future, allowing the production of many suncus mAbs specific for antigens or epitopes that are not immunogenic in rodents.