理論/実験 技術
遊走細胞集団の動的自己組織化についての数理モデルと数値シミュレーション
2023 年 63 巻 4 号 p. 207-211
詳細
2023 年 63 巻 4 号 p. 207-211
多細胞生物が形成され維持されるには,動的な多細胞集合体や秩序だった動きの創発など,細胞集団の動的な自己組織化が重要な場面もあるだろう.本稿では,細胞遊走と接触コミュニケーションによる動的自己組織化に焦点をあて,その数理モデルと数値計算結果例を紹介し,細胞集団の動的自己組織化の原理の一つを提示したい.

生き物の中では構造や集団での動きが様々なスケールで見られる.構造の例は,細胞組織,細胞内小器官,細胞骨格,クロマチン構造など限りがない.それらは生き物を形作ったり機能を生み出したりするのに不可欠である.同時に動きも重要である.例えば動物の形作りには,細胞の変形,分裂,移動など多くの動きが必要である.このような過程を繰り返し,望ましい形の多細胞体を実現する.特に個々の要素がバラバラではなく集団で動くことが重要な場面は多いだろう.
如何にこうした様々な構造や集団的動きが生み出され維持されているかは,細胞生物学や発生生物学に重要な問いであろう.この問いに答えるには,生化学的な側面だけでなく,生物物理学的な側面からの理解も欠かせない.特に生物物理の側面からの重要な概念として自己組織化がある.様々な構造や集団的動きの中には,その形成と維持に自己組織化が関与しているものもあるだろうし,実際そのような例は近年ますます見つかってきている.生き物にどんな自己組織化が起こり得るのか,また実際に見られているのかを知ることは,形態形成や恒常性,機能発現の背後にある機序を理解するための視点の一つを提示するだろう.
本稿の目的は,遊走細胞集団の協調的な振る舞いの結果生じる,動的な自己組織化,つまり動的な多細胞集合体や秩序だった運動の創発を模倣する数理モデル(の一つ)を解説することである.解説は著者らの研究1)-6),特に文献1,5に基づく.
細胞遊走は多くの真核細胞が示す典型的な動きの一つであり,多細胞生物が最終的に複雑な形態を作りあげるのに重要である.さらに,形態形成過程においては,細胞達が協調して遊走することが重要な場合も知られている(例えば神経堤細胞の遊走など7),8)).本稿では,そのような遊走細胞集団の動的自己組織化,特に最も基本的な場合であろう,平面基盤上で遊走する細胞集団の動的自己組織化に焦点をあてる.
遊走細胞達を見渡すと,様々な動的多細胞集合体や協調運動が見られる.例えば,社会性アメーバである細胞性粘菌(Dictyostelium discoideum)に限っても,リング状集合体や回転するマウント構造,或いはバンド状の伝搬密度波などが見られている.如何にして,そうした多彩な動的自己組織化が生じ得るのであろうか.一つのヒントとして,先行研究において,接触運動阻害/誘引(CIL/CAL = Contact Inhibition/Attraction of Locomotion)7),8)や接触追従(CF = Contact Follow)4),9)(文献9ではContact activation of locomotionと呼ばれている)などの接触時の細胞間コミュニケーションが,遊走細胞の協調的行動に重要な役割を果たすと言う知見が蓄積されている.そこで,多彩な動的自己組織化が細胞の遊走能と細胞間接触コミュニケーションによった共通の原理で説明されることが期待される.しかし,そうして多彩なパターンが生み出される様子を実際に示した理論はまだ無かった.
そこで我々は,上述の接触時コミュニケーションを行う遊走細胞集団の数理モデル(図1)を与え,その数値シミュレーションを行った5).この際に,単一細胞の遊走軌跡の解析と数理モデリングに関する著者らの先行研究の知見1),2)と一緒に,アクティブマター物理(用語解説参照)と呼ばれる分野で活発に研究されている自己駆動粒子の集団運動10)の知見を活用した.結果,コミュニケーションの強さを調整するだけで,様々な動的自己組織化パターンが実現されることが明らかになった.以下,この数理モデルの基礎を概説する.また,最後に数値計算結果の一部を紹介する.
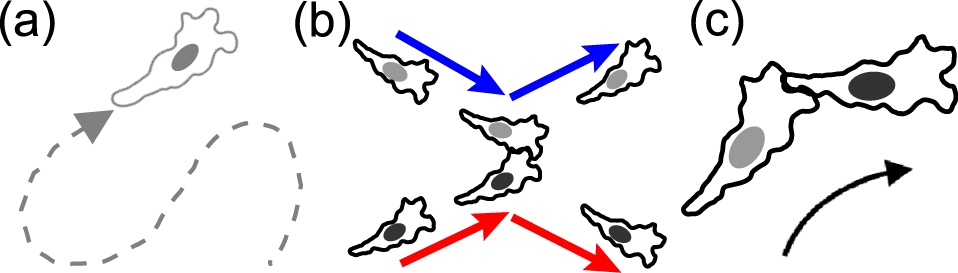
数理モデルの模式図.(a)各細胞は自由に遊走する.但し,ぶつかった際には重なりにくいとする(排除体積効果).細胞間接触コミュニケーションとして,(b)接触運動阻害(CIL),或いはその逆として接触運動誘引(CAL),及び(c)接触追従(CF)を考える.文献5より引用.
遊走細胞の集団の挙動を数理モデル化したいので,まずは1細胞の自由な遊走について理解しモデル化しよう.細胞外からの刺激がなくても,細胞の前後に局在する細胞内シグナル(以下これを細胞内極性と呼ぶ)に従って,自発的に直進性をもって遊走する真核細胞は少なくない.このような遊走の持続性は飢餓状態の細胞性粘菌の遊走の軌跡にも見られる(図2a).遊走方向の自己相関関数をとると,時間差に対して単一の指数関数ではその減数をフィットできず,二つの指数減衰の和で初めてフィットできることがわかる.ここで,一方の時間スケールは数十秒,他方の時間スケールは300秒程度である.つまり,仮足の動きの特徴的時間(数10秒)よりも長い特徴的な運動方向の持続時間があることがわかる.このように,遊走の軌跡を模倣するためには上述の細胞内極性,つまり遊走方向を決める細胞内のコンパスのようなものを考慮した数理モデルが必要になる1).

1細胞遊走の解析とモデリング.(a)遊走方向の自己相関.(b)等方性の自発的な破れ.上が破れない場合.下が破れる場合.著者らの細胞遊走方向モデルでは,下の場合のようなポテンシャルを仮定することで,極性が自発的に形成されるとした.(c, d)数値計算例.方向性の刺激がない場合(c)とある場合(d)の例示.x = +5に周期境界をおいた.文献1より一部改変して引用.
そこで以下の数理モデルを用いた1).各時刻tにおける細胞内極性をベクトルq(t) = (qx(t), qy(t))で表し,それが変分ダイナミクス
| (1) |
に従うと仮定する.ここでポテンシャルはUspp(q) = (Iq/4)(|q|2 – 1)2(Iq > 0)で与えられるとする.つまり,自発的に細胞内極性を確立する能力を,各細胞において等方性が自発的に破れている結果とみなす(図2b).図2b下が上述のUspp(q)の関数形であり,まさに等方性が自発的に破れた状態を導くポテンシャルである.因子Iqは固有極性を維持する強さを制御する係数である.細胞内極性を確立し制御するための細胞内シグナル伝達における揺らぎがあるはずなので,ノイズξq(t)も加えた.簡単のために,
以下では,簡単のため,Iqは極限的に大きいとしよう*1.この場合,qは|q| = 1に強く束縛される.よって極性方向の角度θ(t)をq(t) = (cos θ(t), sin θ(t))で導入すれば,式(1)は
| (2) |
になる.新しいノイズ項ξ(t)も
次に細胞の移動自体についての方程式を与える.遊走方向は細胞内極性の向きで決まるので,その方向
| (3) |
と書ける.v0は遊走速度.なお,仮足の形成や後退など揺らぐ細胞形状に由来して,一般には式(3)右辺に位置ノイズξxが必要だろう.但し,例えば上述の遊走方向自己相関の解析結果では,運動の遅い時間スケールが早い時間スケールに比べて10倍以上遅かった.前者は上述の通りθの変化の時間スケールD–1,後者はおよそ仮足の動きの時間のためξxの時間スケールに対応する.今回はそのような状況を仮定し,遅い時間スケールのダイナミクスに焦点をあて,ξxは無視した.
式(2)と(3)を計算した結果が図2cである.なお,この式はアクティブマターの一種である自己駆動粒子のダイナミクスを記述するためによく使われる式と同じである.そのため,当該分野の先行研究の知見が役に立つわけである.
第6節で走化性(より一般に方向のある刺激への応答)に本モデルを適用した結果も説明するので,その場合の1細胞の数理モデルも説明しておきたい.ここまでと同様,走化性刺激のない状況でも自発的に直進性をもって遊走するとする.真核細胞は,細胞表面の受容体を使って周囲の誘引物質の濃度勾配を空間的に検知し,その検知結果をもとに極性と遊走の方向を制御することで走化性を実現している.そこで,あくまで一定速度で極性方向に遊走するが,勾配検知の結果として生じる極性方向の偏りによって走化性が実現されると仮定する1),2).これにより式(1)の代わりに
| (4) |
に従い極性qが時間発展する1).Iqが無限大の極限とすれば,角度表記でdθ(t)/dt = –fx sin θ + fy cos θ + ξ(t)とも書ける.ベクトルf = (fx, fy)は方向性をもった外部刺激が結果として極性に与えるバイアスを表す.
式(3)と(4)の解析から,濃度が一様な場合(f = 0)でも誘引物質は細胞遊走に影響を与えること,具体的には遊走方向の持続時間を増加させる(直進性を増す)ことが示された1).この予想は細胞性粘菌の実験でも観測された.また,細胞内極性の存在に由来した遊走方向持続性が走化性精度を向上させることが示された1).細胞性粘菌などは即時的な勾配検知では物理的に不可能なほど高い精度で走化性運動することが知られている.その高い走化性精度が極性由来の遊走方向持続性により説明可能であることが示唆される.さらに,数値計算結果の軌跡(図2d)を見てみると,以下のような面白い性質がわかる.一度遊走方向を間違えると,細胞内極性の持続性のためになかなか正しい方向に向き直せない.このような性質は実際,細胞性粘菌の遊走の軌跡にも見られる.
さて本稿の主題は集団挙動であるので,単一細胞の遊走についての説明はここまでにしたい.単一細胞の話にさらに興味のある読者は文献1,2を参照のこと.
前節で単一細胞の遊走の特徴がわかり数理モデルを得た.次にそれらの間の相互作用を考えることで,多細胞挙動のモデル化に移りたい.そのために有用な先行研究の知見を本節でまとめておこう.
(1)細胞間接触コミュニケーションについての知見:以下の接触運動阻害/誘引(CIL/CAL)と接触追従(CF)と呼ばれる細胞間のやりとりが色々な遊走細胞で見られている.(1-1)CIL:ある種の細胞では,2つの細胞が接触した際に,互いに重ならないように各々の細胞が細胞内極性を調整して,あたかもその2細胞が散乱しているかのように振る舞うことが見られる(図1b).これは,例えばアフリカツメガエルの神経堤細胞でよく調べられており,協調的で方向性のある遊走を生み出すことで形態形成に関与していることが知られている7).(1-2)CAL:ある種の細胞は,2つの細胞が接触した際に,CILの逆に互いに引き合うように極性を変調する8).(1-3)CF:ある種の細胞集団では,細胞が前後に連なった時に,後ろの細胞が前の細胞を追いかける動きを示す(図1c).この細胞間コミュニケーションは細胞性粘菌含む幾つかの細胞で観察されており,その分子シグナル機構も調べられている9).CFは回転するマウント構造や伝搬密度波の形成に関与していることが知られている4),9).
(2)集団運動の数理研究からの知見:次にこれらコミュニケーションの各々が集団挙動にどう影響し得るかを,自己駆動粒子系の集団運動の知見に基づき考察する.(2-1)CFは,前後の細胞で非相反的な局所相互作用を引き起こす.この対称性のみ考えれば,これは逃避と追跡による群れ運動モデルで想定される追跡相互作用の仲間である.例えばこれは物体群が鎖状に連なって動く集団挙動を誘発することが知られている.(2-2)次にCIL/CALについて考察する.CILは細胞内極性に作用する短距離反発を実効的に生み出す.これは,動物の群れモデルにおける,局所的な認知的反発の効果に相当する.このような相互作用は,単独では,粒子間の散乱のような効果を与える.さらに,それと体積排除を組み合わせると,近くの粒子の移動方向が揃う効果が実効的に生じる.CALはCILの逆の結果を生むだろう.それが凝集を引き起こすことは自然に想像される.短距離選択的引力-反発による集団運動の数値シミュレーションの結果は示唆的である.反発優位の場合は,極性秩序(本記事では細胞達の極性のPolar orderの意とする)をもつ伝搬密度波を誘発したり,極性秩序をもつ均一な群れ運動を誘発したりするが,誘引優位の場合は凝集を誘発する.CIL/CALでも同様の現象が期待される.(なおこれらに関する参照文献は文献5から辿ることができる.)よって,CFとCIL/CALを合わせれば十分に多彩な動的自己組織化パターンが誘発され得ると期待される.
本節では多細胞の場合の数理モデルについて説明する.2次元空間中で,周期境界をもつ正方形の領域の中に複数(N個)の細胞がいるとしよう.各細胞は第3節と同じく細胞内極性をもった点として近似し,極性は2次元のベクトルqj(t)で記述する(tは時刻,jは細胞の番号j = 1, 2, ..., N).j番目の細胞の位置xj(t)と極性qj(t)は,式(1)と(3)の拡張として,
| (5) |
| (6) |
にそれぞれ従って時間発展する*2.
体積排除は,各細胞を半径r固定のソフトコアの反発ディスクに理想化し,
細胞間接触コミュニケーション
本稿ではこれまで,遊走細胞の動的自己組織化を模倣するための数理モデルを概説してきた.数理モデルは単一細胞の自由な遊走に細胞間接触コミュニケーションの寄与を加えて構築された.最後に,この数理モデルの数値シミュレーション結果とそこから学べたことの例を紹介して解説を終えよう.
様々なαCF及びαCILに対して数値計算を行った結果を図3にまとめた.他のパラメータ値は一定にした.第3節と同じくIqは極めて大きいとした.図3bが結果として得られるパターンの例である.期待通り,リング状パターンや回転する集積体,極性秩序をもつ伝搬密度波など多彩なパターンが見られた5).これらは先述した細胞性粘菌が実際に見せる挙動を含む.また伝搬密度波に関してより詳しい比較を文献4で行った.さらに,図3cのようなSDA = Snake-like Dynamic Assembly とでも言える新しい集団遊走の様式も発見された5).この状況では,細胞は蛇状の集合体を形成している.しかし,これら集合体は非常にダイナミックで,変形し,移動し,融合・消失したり新たに出現したりを繰り返し,一定の形に落ち着くことがない.

遊走細胞集団の動的自己組織化の数値シミュレーション結果.(a)相図.(b)典型的スナップショット.左から,リング状,渦状,極性秩序のある伝搬密度波状のパターン.(c)シミュレーションにより新しく見つかった集団遊走の形式(「Snake-like dynamic assembly」).各点の位置が各細胞の位置を,色がその細胞の極性の向き(図中の色相環を参照)を表す.黒矢印はいくつかの代表細胞の遊走方向を表す.いずれも細胞数密度は1.D = 0.1(おおよそ飢餓状態の細胞性粘菌の値1/300[s–1]を無次元化した値).文献5より一部改変して引用.
このように,典型的な2種類のコミュニケーションの強さを調整するだけで,多彩な動的自己組織化パターンが一つの原理から生み出されるのである.細胞間の接触時のコミュニケーションは,各細胞の自由な遊走と組み合わさることで,多彩な動的自己組織化パターンを誘発する可能性を有していることが示された.
ここで示したものはあくまで動的自己組織化の原理の一つである.例えば,実際の細胞集団では,細胞性粘菌や神経堤細胞に見られるように,各細胞自身から分泌された走化性因子のような化学物質を介した非局所的なコミュニケーションも役割を果たすだろう.これは提示したモデルには組み込まれていない.しかしながら本結果は,何か特定の(実験で観測されたなど)複雑な動的自己組織化の現象を説明したい際に,接触を介したメカニズムの可能性を無視できないと言う教訓を我々に知らしめている.
さて,現実的なパターンが出たのは良いとして,理論モデルを作ったことのより大事な意義は,それをもとにさらなる議論を発展させることにある.このような動的自己組織化能が細胞集団の機能的な振る舞いに果たしうる役割について,本数理モデルを応用することで調べた研究例も紹介しよう.
図4では,僅かな方向性刺激を外から加えた場合(バイアスf = (0, 0.01))の結果3),5)を示した.このバイアスは非常に小さいため,個々の細胞,つまりコミュニケーションのない場合の細胞は,図4aのように殆ど反応できない.その程度の微かなバイアスを与えた.このような極微かなバイアスであっても,細胞がやりとりしてSDAを含むあるクラスの動的自己組織化パターンを形成する場合には,図4bのように個々の細胞が平均的に正確に応答できるようになることがわかった*3.細胞達のオーケストレーションにより個々の細胞の能力を遥かに超えた全体としての能力が誘起されると言える.

また,狭い領域中での遊走に動的自己組織化が如何に関与し得るかも調べた6)が紙面の都合で割愛する.
平岩徹也(ひらいわ てつや)
中央研究院物理研究所副研究員
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/63-4ie2270p01.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)