Abstract
The onset of oral candidiasis is accompanied by inflammatory symptoms such as pain in the tongue, edema or tissue damage and lowers the quality of life (QOL) of the patient. In a murine oral candidiasis model, the effects were studied of terpinen-4-ol (T-4-ol), one of the main constituents of tea tree oil, Melaleuca alternifolia, on inflammatory reactions. When immunosuppressed mice were orally infected with Candida albicans, their tongues showed inflammatory symptoms within 24 h after the infection, which was monitored by an increase of myeloperoxidase activity and macrophage inflammatory protein-2 in their tongue homogenates. Oral treatment with 50 µL of 40 mg/mL terpinen-4-ol 3h after the Candida infection clearly suppressed the increase of these inflammatory parameters. In vitro analysis of the effects of terpinen-4-ol on cytokine secretion of macrophages indicated that 800 µg/mL of this substance significantly inhibited the cytokine production of the macrophages cultured in the presence of heat-killed C. albicans cells. Based on these findings, the role of the anti-inflammatory action of T-4-ol in its therapeutic activity against oral candidiasis was discussed.
Candida albicans is known as an opportunistic infectious microorganism that causes oral candidiasis.1) The major afflictions induced by onset of oral candidiasis are inflammatory symptoms accompanied by discomfort, pain of the tongue, dysgeusia and bad breath all of which greatly lower the quality of life of the patient. In most cases, oral candidiasis is effectively treated with anti-fungal drugs,2–4) but recurrence after termination of chemotherapy which arises in favor of C. albicans, the commensal yeast, and emergence of an anti-fungal drug resistant species as a result of repeated application of that drug have become serious issues.5,6) Therefore, development of a new type of treatment is highly sought after.
As reported previously, some of the essential oils,7,8) fatty acids9) or other substances10,11) are therapeutically efficacious to murine oral candidiasis. In preceding papers, we have reported that oral treatment with tea tree oil (TTO) and its main component, terpinen-4-ol (T-4-ol), improved Candida infected lesions on the tongue’s surface and lowered the number of Candida cells in the murine oral cavity.7,12) Interestingly, although TTO and T-4-ol showed an efficacy in oral candidiasis, the recovery of lesion score and the decrease in viable Candida cell number were not correlated7); that led us to assume that there might be several factors affecting the therapeutic results other than the direct anti-Candida efficacy.
In our previous histopathological research, oral treatment with T-4-ol was suggested to inhibit accumulation of inflammatory cells at local sites in Candida infected tongue mucosa.7) Experimental findings in vitro indicated that TTO and/or T-4-ol suppress hyphal growth of C. albicans at low concentrations12,13) or possess anti-inflammatory activity.14,15) In this study, we investigated quantitatively: 1. inflammatory responses in murine oral candidiasis, and 2. suppressive efficacy of T-4-ol against the inflammation. We believe that our findings about the anti-inflammatory activity of T-4-ol on murine oral candidiasis as given here explain part of the mechanism of the therapeutic efficacy of this terpenoid.
Materials and Methods
OrganismsThe Candida albicans strain TIMM 1768 was clinically isolated from patients and maintained at Teikyo University Institute of Medical Mycology. This strain was stored at −80°C in Sabouraud dextrose broth (Becton Dickinson, MD, U.S.A.) containing 0.5% yeast extract (Becton Dickinson) and 10% glycerol until the experiment was performed.
AgentsTerpinen-4-ol was purchased from Tokyo Chemical Industry Co. (Tokyo, Japan). Lipopolysaccharide (LPS), hexadecyltrimethylammonium bromide (HTAB), human myeloperoxidase (MPO) and tetramethylbenzidine (TMB) were purchased from Sigma-Aldrich Japan (Tokyo, Japan).
AnimalsAnimal experiments were performed according to the guidelines for the care and use of animals approved by Teikyo University and guidelines for animal experiments conducted at research institutions by the Ministry of Education, Culture, Sports, Science and Technology of Japan. Six week-old female ICR mice (Charles River Japan, Kanagawa, Japan) were used for all animal experiments. The photoperiods were adjusted to 12 h of light and 12 h darkness daily, and the environmental temperature was constantly maintained at 21°C. The mice were kept in cages housing 3 animals and given access to food and water ad libitum.
Murine Oral CandidiasisExperimental procedure of the murine oral candidiasis model was described previously16) and partly modified. Briefly, immunosuppressed mice were induced by subcutaneous treatment with a 100 mg/kg dose of prednisolone (Kyoritsu Seiyaku Corporation, Tokyo, Japan) 1 d prior to oral infection. Tetracycline hydrochloride (Sumika Enviro-Science, Tokyo, Japan) in drinking water at a dose of 4.2 mg/mL was given to the animals from 1 d before infection until the experiment ended. On the day of Candida inoculation, mice were anesthetized by intramuscular injection with 14.4 mg/kg of chlorpromazine chloride (Wako Pure Chemical Industries, Ltd., Osaka, Japan) in the left femur; subsequently, oral infection was induced with about 2×108 cells/mL viable cells of C. albicans (TIMM1768) in RPMI1640 medium containing fetal bovine serum (FBS), the complete medium. The oral infection was performed by means of a cotton swab (baby cotton buds; Johnson & Johnson, Tokyo, Japan) rolled over the entire tongue in the mouth. The cell number of C. albicans inoculated in the oral cavity was calculated to be approximately 1×106 cells/mouse by the difference in viable cell number adhering to the cotton swabs before and just after oral inoculation.16)
Oral TreatmentT-4-ol was suspended in 1% Tween 80 (Wako Pure Chemical Industries, Ltd.) and applied to the oral cavity with feeding needles 3 h after C. albicans inoculation as previously reported.7,17) One percent Tween 80 was applied to the control group in the same manner.
Quantitation of Viable C. albicans from Oral Cavity and Homogenization of Infected TonguesTwenty-four hours after Candida inoculation, groups of mice were sacrificed and the Candida cells in the oral cavity were collected with a cotton swab. After swabbing this cavity, the end of the swab was cut off and placed in a tube containing 5 mL sterile saline. The Candida cells were re-suspended by mixing with a vortex mixer and diluted with a series of 20 and 100-fold dilution of sterile saline. Fifty microliters of each dilution was incubated on a Candida GS agar plate for 20 h at 37°C followed by counting of the CFU (colony forming units).
To measure MPO activity as well as the quantity of cytokines in the Candida infected tongues, the supernatants of the homogenized tongues were frozen until the experiments. The resected tongues were placed in 2 mL of 0.5% HTAB for MPO assay or the complete medium for enzyme-linked immunosorbent assay (ELISA) and were homogenized followed by centrifugation at 3400 rpm for 10 min. The retrieved supernatant was stored at −80°C until measured.
MPO AssayMeasurement of MPO activity was based on the method of Maruyama et al.18) and partly modified. The samples from −80°C were thawed and poured into 96 well micro plates (30 µL/well). Subsequently, 200 µL of a mixture containing 100 µL of phosphate buffered saline, 85 µL of 0.22 m sodium phosphate buffer and 15 µL of 0.017% hydrogen peroxide was added to the wells. The reaction was started by the addition of 20 µL of 18.4 mm TMB in 8% aqueous dimethylformamide. Plates were incubated at 37°C for 3 min and then placed on ice where the reaction was stopped by addition to each well of 30 µL of 1.46 m sodium acetate. The absorbance of samples was measured with 620 nm (optical density (OD) value) and converted into MPO values per 0.1 g of murine tongues.
ELISAAn ELISA assay kit was purchased from BD Biosciences (CA, U.S.A.) for tumor necrosis factor-alpha (TNF-α) and R&D Systems (Minnesota, U.S.A.) for macrophage inflammatory protein-2 (MIP-2). The supernatants of tongues were adjusted as described earlier and ELISA was conducted according to the manufacturer’s instructions.
Preparation of Heat-Killed Candida albicans for in Vitro AssayHeat-killed Candida albicans (HKCA) prepared using the following protocol was applied for in vitro experiments. C. albicans was grown on Sabouraud dextrose for 16h and harvested in RPMI1640 medium containing 2.5% FBS. Half of the culture medium containing Candida cells was immersed in warmed 95°C water for 20 min and stored at −80°C until experiments (HKCA yeast). The other half was adjusted to 1×107 cells/mL and poured into petri dishes followed by incubation at 37°C, 5% CO2 for 3 h. This incubation allowed the Candida to form germ tubes. The germinated cells were collected, heated and stored in the same manner until further experiments (HKCA hyphae).
Preparation of Murine Macrophage MonolayerMurine macrophages were prepared from peritoneal exudate by the method described previously19) and partly modified. Briefly, ICR mice were intraperitoneally injected with 2 mL of 3% Fluid thioglycollate medium (Becton Dickinson) in distilled water 3 d before the collection of peritoneal exudate cells (PEC). The collected PEC were centrifuged at 1500 rpm, 4°C for 5 min and adjusted to 1×106 cells/mL with a hemocytometer followed by applying 200 µL/well into 96-well microplates. The cells were incubated for 2.5 h at 37°C in 5% CO2; non-adherent cells were gently washed out with 37°C PBS resulting in a monolayer which consisted of macrophages (>95%).19)
Measurement of Anti-inflammatory Activity of T-4-ol in VitroHKCA, LPS and T-4-ol were prepared with the complete medium and added to the macrophage-monolayer followed by 37°C in 5% CO2 incubation. The tested concentration of T-4-ol was adjusted to 200, 400 and 800 µg/mL, since a previous report20) showed that epithelial cells or fibroblasts were remained viable at least after short time exposure of less than 1000 µg/mL of T-4-ol. After the incubation, collected culture supernatants were centrifuged for 2000 rpm at 4°C for 5 min and the supernatants were stored at −80°C until the measurement.
Histopathological ObservationThe murine tongues were fixed in 4% paraformaldehyde solution and embedded in paraffin for histopathological study. Five-micron sections were obtained from the paraffin block and stained with periodic acid-Schiff (PAS) stain.
Statistical AnalysisAll results of statistical analysis were analyzed using unpaired t-tests. In all cases, p<0.05 was considered to be significant.
Results
Kinetics of MPO Activity in Murine Oral Candidiasis and MPO Suppression of T-4-olFirst, the neutrophil accumulation was evaluated by measuring MPO activity, the marker of neutrophils, as an inflammatory response in Candida-infected tongues; these tongues showed a clear increase of MPO activity on average although it varied among individual mice. The mean value of MPO activity was 101.21±59.39 munits/mouse tongue (n=6) in a Candida-infected group compared with 7.11±3.64 munits/mouse tongue (n=6) in an uninfected group 24 h after Candida inoculation, which means that MPO activity of Candida infected tongues increase 14 folds to the uninfected group.
Oral treatments with 50 µL of T-4-ol solution at the concentration of 10 mg/mL significantly decreased MPO activity of infected tongues (Fig. 1a). T-4-o1 was applied once 3h after the Candida inoculation. In this experiment, we observed that viable Candida cell numbers in the oral cavity were also decreased significantly at 40 mg/mL, but not at 10 mg/mL, although a decreasing tendency was appeared (Fig. 1b).
Histopathological Observation of Candida Infected Tongues Treated by T-4-olPAS stained sections of tongues, prepared 24 h after Candida infection, were observed as in Fig. 2. Hyphal extension of C. albicans and accumulation of inflammatory cells were observed in the control group (Fig. 2b), but not in the uninfected group (Fig. 2a). The amount of hyphae was diminished dose dependently by oral application of T-4-ol although some were still present on the mucosal surface (Figs. 2c, d). No accumulation of inflammatory cells was observed in the local infected site, which seemed clearly different from the control group.
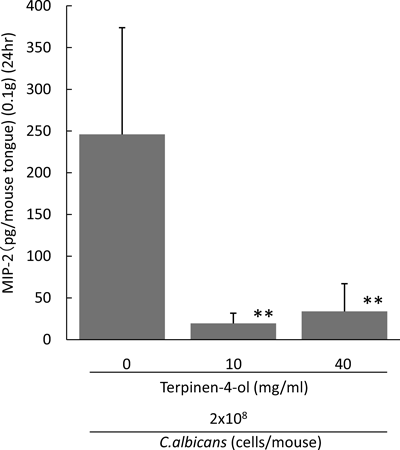
T-4-ol Suppresses Production of MIP-2 in Murine Oral CandidiasisSince it is known that MIP-2 functions as a major chemotactic factor for neutrophils to inflammatory sites in a murine body, the kinetics of the amount of MIP-2 produced in Candida-infected tongues was evaluated. In a control experiment, the tongues of Candida-uninfected mice with or without T-4-ol (40 mg/mL) were homogenized to determine their MIP-2 amount followed by the ELISA assay. The average quantity of MIP-2 was 0±1.01 or 6.95±20.89 pg/tongue (0.1 g) (n=6), which indicated that T-4-ol treatment to uninfected tongues did not significantly affect the level of MIP-2. The amount produced in Candida-infected groups treated with 10 or 40 mg/mL of T-4-ol was suppressed significantly comparing to the control group as shown in Fig. 3.
Measurement of TNF-α Production by Murine Macrophages Stimulated with Heat-Killed Candida Cells in VitroAs described above, oral application of T-4-ol seemed to suppress local inflammation in Candida infected tongues. From these results, we performed in vitro studies to examine whether T-4-ol possesses direct suppressive activity to production of inflammatory cytokines from Candida-stimulated macrophages. It is reported that tissue invasion of Candida hyphae causes production of inflammatory cytokines.21) As a major inflammatory cytokine, TNF-α which is reportedly secreted in a short time (2 h) after exposure to C. albicans,22) was evaluated by supernatants from murine peritoneal macrophages cultured with HKCA.
To determine whether there is a difference of production amount of the cytokine between HKCA yeast and hyphae, each of stimulatory activities was tested to murine macrophages. Adjusted concentrations of HKCA yeast or hyphae were added to a macrophage monolayer followed by 2 h incubation at 37°C, and TNF-α amount in the culture supernatants was then measured with ELISA. The results showed that TNF-α was produced more effectively after 2 h culture of macrophages stimulated with HKCA hyphae, than with HKCA yeast (Fig. 4) .

The effects of T-4-ol on TNF-α production by murine macrophages were examined. Macrophage monolayer was cultured in the presence of HKCA hyphae with or without T-4-ol for 2, 6, or 20 h cultivation and then the culture supernatants were obtained for measurement of TNF-α secretion (Fig. 5a). T-4-ol suppressed the production of TNF-α dose dependently and its 50% inhibitory concentration (IC50) at 2 or 6 h culture was estimated to be under 800 µg/mL. The level of TNF-α in 20 h of culture supernatant seemed to be lowered. Figure 5b shows that stimulation with LPS elicited a greater production of TNF-α than HKCA hyphae and that TNF-α production in the case of LPS-stimulation was not similarly suppressed by 800 µg/mL of T-4-ol. Lowered suppression efficacy of T-4-ol (800 µg/mL) at with 6 or 20 h incubation was also confirmed.
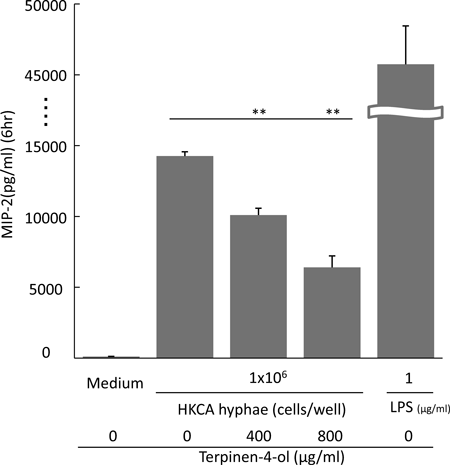
T-4-ol Suppresses MIP-2 Production in VitroThe effects of T-4-ol on MIP-2 production by macrophages was also evaluated in vitro. In the presence of T-4-ol, macrophages were cultured with 1×106 cells/mL of HKCA hyphae for 6h and the production amount of MIP-2 in the supernatants was evaluated with ELISA. Figure 6 clearly shows that a non-stimulated macrophage monolayer produced little MIP-2 in contrast to the groups that cultured with HKCA hyphae. T-4-ol suppressed these MIP-2 productions to less than 50% at the concentration of 800 µg/mL.
Discussion
Candida albicans, especially Candida cells with hyphal growth, causes oral candidiasis by invading oral mucosal tissues; the Candida-infected lesions are often accompanied by pain in the tongue, redness and swelling.23) In experimental murine oral candidiasis, it was reported previously that inflammatory cells, mainly consisting of neutrophils, accumulated in Candida-infected lesions.7,24) Since inflammatory responses with these cells would cause major pathological changes in oral candidiasis, we believe that the inflammatory severity of infected tongues should be evaluated quantitatively in order to understand the pathological conditions of this disease. In this study, the following two findings were obtained: 1. a marker of neutrophils, the MPO activity, was increased in the tongue homogenates after 24 h of oral Candida infection; 2. oral treatment with T-4-ol suppressed this increase of MPO activity, which might be caused by inhibition of the production of MIP-2, a mouse homologue of human IL-8 which could stimulate the migration of neutrophils. The decrease of MPO activity in tongue tissues by T-4-ol application indicates that this monoterpene alcohol possesses not only anti-Candida activity but also anti-inflammatory efficacy in its therapy against oral candidiasis.
Since cellular distribution of MPO is limited mainly to neutrophils,25) it can be used as a marker of these cells. Our experiments showed that MPO activity in Candida-infected tongues at 24 h increased to more than 14 times that of a non-infected group. This means that neutrophils, the inflammatory cells, were accumulated in the local sites of Candida infection. As far as we know, this is the first report enabling quantitation of the degree of inflammatory symptoms of tongues in murine oral candidiasis.
Treatment of T-4-ol suppressed the increase of MPO activity in tongues 24 h after Candida infection (Fig. 1a), which might correspond to the histological observation suggesting inhibition of the local accumulation of neutrophils. We speculated that this suppression might be caused by the inhibited production of MIP-2 in the tongue tissue, which may be produced by macrophages, epithelial cells and other inflammatory cells in the event of Candida infection. Our experimental results showed that T-4-ol treatment suppressed production of MIP-2 in tongues with murine oral candidiasis (Fig. 3), which is apparently related to diminution of MPO activity. These findings indicate that T-4-ol exerts anti-inflammatory activity in oral candidiasis therapy.
Cytokine production by macrophages, which is considered to play an essential role in the early stage of inflammation,26) and the effects of T-4-ol on the production were examined in vitro. Comparison of TNF-α production from murine macrophages stimulated by HKCA yeast and hyphae revealed that the hyphae form more strongly induced the cytokine production than did the yeast form (Fig. 4). The secretion of TNF-α and MIP-2 from macrophages stimulated with HKCA hyphae was suppressed by T-4-ol in vitro. This suppression was demonstrated in the case of 2 h culture for TNF-α and 6 h culture for MIP-2 (Figs. 5a, 6). It was previously reported that Candida hyphae started to invade the mucosal tissues of tongues 3 h after Candida-oral inoculation.27) This evidence appears to explain the mechanism of prompt suppression of inflammatory responses in vivo, that is, T-4-ol treatment 3 h after Candida inoculation suppresses the increase of MIP-2 and MPO activity very rapidly for up to 24 h after the infection. In this context, we wish to note that the low molecular weight of this terpenoid must enable its quick infiltration into the mucosal tissues of the tongues, as true of cinnamaldehyde.8)
We have assumed that the therapeutic activities of terpenoids including T-4-ol on candidiasis depend on its anti-Candida-cell actions. In fact, we have reported that minimum inhibitory concentration (MIC) of T-4-ol for yeast-form growth of TIMM 1768 is about 40 mg/mL and even 800 µg/mL of T-4-ol inhibits mycelial growth of the Candida cells in vitro.12) Therapeutical activity of T-4-ol, especially decrease of viable candida cell numbers collected from oral cavity, should attribute to inhibition of mycelial growth of this fungus.7) In this report, we showed that T-4-ol application at a dose of 50 µL of 40 mg/mL, significantly decreased the viable Candida cell number in the oral cavity (Fig. 1b). Therefore, we can speculate that decrease of the Candida burden on the tongue surface by T-4-ol might negatively affect the inflammatory responses including MIP-2 production. Although this speculation is possible, we postulate the importance of the anti-inflammatory activity of T-4-ol in our therapeutic model, because the increase of MPO activity in the infected tongues was inhibited even with a lower dose (10 mg/mL) of T-4-ol. Moreover, histopathological findings indicate that the existence of mycelia of Candida cells in tongue tissue treated with T-4-ol did not induce an accumulation of inflammatory cells as shown in Figs. 2c and d. Since suppression of accumulation of inflammatory cells by T-4-ol was demonstrated at lower concentrations, we may consider that the impact of anti-inflammatory activity of T-4-ol must be equal to or more potent than direct anti-Candida action in improving the symptoms of oral candidiasis. To support this assumption, we found there was no correlation between MPO activity and viable Candida cell number in the oral cavity of individual mice treated with T-4-ol (data not shown).
Since T-4-ol possesses anti-inflammatory activity in vitro and in vivo, we must discuss the possibility that this terpene alcohol may suppress the self-defense mechanisms against microbial infection. As shown in Fig. 5b, the presence of T-4-ol (800 µg/mL) did not suppress TNF-α production from the macrophage monolayer stimulated by LPS. This result indicates that anti-inflammatory activity of T-4-ol might be varied by different stimulants such as HKCA hyphae or LPS, a water-soluble material. We also wish to note that in the production amount of TNF-α in LPS, the control group did not differ from the T-4-ol treated groups cultured for 6 or 20 h. This means that T-4-ol did not exert cytotoxicity non-specifically to macrophages, at least under our experimental conditions. Therefore, we can speculate that T-4-ol does not non-specifically affect defense mechanisms to various infections; this opinion can also be supported by excellent traditional estimates of the therapeutic efficacy of TTO on various cutaneous infectious diseases.28–30) However, in this context we should note that further precise study on influences of T-4-ol to killing activity of macrophages and neutrophils is needed.
Our study showed that the anti-inflammatory activity of T-4-ol is one of the key roles of its therapeutic efficacy against oral candidiasis. TTO, the essential oil mainly composed of T-4-ol, has been employed empirically against various diseases that include inflammation-related disorders.28,31) The anti-inflammatory activity of T-4-ol could explain the reason why this terpene alcohol has been used for those treatments. Although further study is needed, we hope that T-4-ol can serve not only in oral candidiasis therapy but against various inflammatory diseases; the significance of this agent would be increased in terms of its prevention or therapy of disorders and amelioration of the weaker QOL caused by inflammation.
REFERENCES
- 1) Odds FC. Candida and Candidosis, a review and bibliography. 2nd ed., Bailliere Tindale, London, pp. 115–152 (1988).
- 2) Garber GE. Fluconazole: a new option in the treatment of Candida mucositis and esophageal candidiasis. J. Otolaryngol., 21, 92–94 (1992).
- 3) Lewis MA, Samaranyake LP, Lamey PJ. Diagnosis and treatment of oral candidosis. J. Oral Maxillofac. Surg., 49, 996–1002 (1991).
- 4) Ghannoum MA, Kuhn DM. Voriconazole—better chances for patients with invasive mycoses. Eur. J. Med. Res., 31, 242–256 (2002).
- 5) Masci JR. Complete response of severe, refractory oral candidiasis to mouthwash containing lactoferrin and lysozyme. AIDS, 14, 2403–2404 (2000).
- 6) Rautemaa R, Richardson M, Pfaller M, Koukila-Kähkölä P, Perheentupa J, Saxén H. Decreased susceptibility of Candida albicans to azole antifungals: a complication of long-term treatment in autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) patients. J. Antimicrob. Chemother., 60, 889–892 (2007).
- 7) Ninomiya K, Maruyama N, Inoue S, Ishibashi H, Takizawa T, Oshima H, Abe S. The essential oil of Melaleuca alternifolia (tea tree oil) and its main component, terpinen-4-ol protect mice from experimental oral candidiasis. Biol. Pharm. Bull., 35, 861–865 (2012).
- 8) Taguchi Y, Takizawa T, Ishibashi H, Sagawa T, Arai R, Inoue S, Yamaguchi H, Abe S. Therapeutic effects on murine oral candidiasis by oral administration of cassia (Cinnamomum cassia) preparation. Nippon Ishinkin Gakkai Zasshi, 51, 13–21 (2010).
- 9) Takahashi M, Inoue S, Hayama K, Ninomiya K, Abe S. Inhibition of Candida mycelia growth by a medium chain fatty acids, capric acid in vitro and its therapeutic efficacy in murine oral candidiasis. Med. Mycol. J., 53, 255–261 (2012), in Japanese.
- 10) Kamagata-Kiyoura Y, Abe S, Yamaguchi H, Nitta T. Protective effects of human saliva on experimental murine oral candidiasis. J. Infect. Chemother., 10, 253–255 (2004).
- 11) Ishijima SA, Hayama K, Burton JP, Reid G, Okada M, Matsushita Y, Abe S. Effect of Streptococcus salivarius K12 on the in vitro growth of Candida albicans and its protective effect in an oral candidiasis model. Appl. Environ. Microbiol., 78, 2190–2199 (2012).
- 12) Ninomiya K, Hayama K, Ishijima SA, Takahashi M, Kurihara J, Abe S. Effects of inhibitory activity on mycelial growth of Candida albicans and therapy for murine oral candidiasis by the combined use of terpinen-4-ol and a middle-chain fatty acid, capric acid. Yakugaku Zasshi, 133, 133–140 (2013).
- 13) Hammer KA, Carson CF, Riley TV. Melaleuca alternifolia (tea tree) oil inhibits germ tube formation by Candida albicans. Med. Mycol., 38, 355–362 (2000).
- 14) Brand C, Ferrante A, Prager RH, Riley TV, Carson CF, Finlay-Jones JJ, Hart PH. The water-soluble components of the essential oil of Melaleuca alternifolia (tea tree oil) suppress the production of superoxide by human monocytes, but not neutrophils, activated in vitro. Inflamm. Res., 50, 213–219 (2001).
- 15) Hart PH, Brand C, Carson CF, Riley TV, Prager RH, Finlay-Jones JJ. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res., 49, 619–626 (2000).
- 16) Takakura N, Sato Y, Ishibashi H, Oshima H, Uchida K, Yamaguchi H, Abe S. A novel murine model of oral candidiasis with local symptoms characteristic of oral thrush. Microbiol. Immunol., 47, 321–326 (2003).
- 17) Hisajima T, Maruyama N, Tanabe Y, Ishibashi H, Yamada T, Makimura K, Nishiyama Y, Funakoshi K, Oshima H, Abe S. Protective effects of farnesol against oral candidiasis in mice. Microbiol. Immunol., 52, 327–333 (2008).
- 18) Maruyama N, Sekimoto Y, Ishibashi H, Inouye S, Oshima H, Yamaguchi H, Abe S. Suppression of neutrophil accumulation in mice by cutaneous application of geranium essential oil. J. Inflamm. (Lond.), 2, 1–11 (2005).
- 19) Tokuda Y, Tsuji M, Yamazaki M, Kimura S, Abe S, Yamaguchi H. Augmentation of murine tumor necrosis factor production by amphotericin B in vitro and in vivo. Antimicrob. Agents Chemother., 37, 2228–2230 (1993).
- 20) Ramage G, Milligan S, Lappin DF, Sherry L, Sweeney P, Williams C, Bagg J, Culshaw S. Antifungal, cytotoxic, and immunomodulatory properties of tea tree oil and its derivative components: potential role in management of oral candidosis in cancer patients. Front Microbiol., 3, 1–8 (2012).
- 21) Blander JM, Sander LE. Beyond pattern recognition: five immune checkpoints for scaling the microbial threat. Nat. Rev. Immunol., 12, 215–225 (2012).
- 22) Kim HS, Choi EH, Khan J, Roilides E, Francesconi A, Kasai M, Sein T, Schaufele RL, Sakurai K, Son CG, Greer BT, Chanock S, Lyman CA, Walsh TJ. Expression of genes encoding innate host defense molecules in normal human monocytes in response to Candida albicans. Infect. Immun., 73, 3714–3724 (2005).
- 23) Yamaguchi H. Byougenshinkin to Shinkinsho. 4th ed., Nanzando, Tokyo, pp. 236–241 (2007).
- 24) Okada M, Hisajima T, Ishibashi H, Miyasaka T, Abe S, Satoh T. Pathological analysis of the Candida albicans-infected tongue tissues of a murine oral candidiasis model in the early infection stage. Arch. Oral Biol., 58, 444–450 (2013).
- 25) Klebanoff SJ. Myeloperoxidase: friend and foe. J. Leukoc. Biol., 77, 598–625 (2005).
- 26) Romani L. Immunity to fungal infections. Nat. Rev. Immunol., 4, 1–23 (2004).
- 27) Hisajima T, Ishibashi H, Yamada T, Nishiyama Y, Yamaguchi H, Funakoshi K, Abe S. Invasion process of Candida albicans to tongue surface in early stages of experimental murine oral candidiasis. Med. Mycol., 46, 697–704 (2008).
- 28) Inoue S, Abe S. An invitation to anti-infectious aromatherapy. 1st ed., Fragrance Journal, Tokyo, pp. 141–224 (2011).
- 29) Markum E, Baillie J. Combination of essential oil of Melaleuca alternifolia and iodine in the treatment of molluscum contagiosum in children. J. Drugs Dermatol., 11, 349–354 (2012).
- 30) Soukoulis S, Hirsch R. The effects of a tea tree oil-containing gel on plaque and chronic gingivitis. Aust. Dent. J., 49, 78–83 (2004).
- 31) Schnaubelt K. Advanced Aromatherapy: the science of essential oil therapy. 1st ed., Healing Arts Press, Vermont, pp. 31–124 (1998).