Abstract
Growth and differentiation factor 3 (GDF3), a mammalian-specific transforming growth factor β ligand, and OCT4, one of key stem cell transcription factors, are expressed in testicular germ cell tumors (TGCTs) as well as pluripotent stem cells. To understand the molecular mechanism by which OCT4 and GDF3 function in tumorigenesis as well as stemness, we investigated the transcriptional regulation of GDF3 mediated by OCT4 in human embryonic carcinoma (EC) NCCIT cells, which are pluripotent stem cells of TGCTs. GDF3 and OCT4 was highly expressed in undifferentiated NCCIT cells and then significantly decreased upon retinoic acid-induced differentiation in a time-dependent manner. Moreover, GDF3 expression was reduced by short hairpin RNA-mediated knockdown of OCT4 and increased by OCT4 overexpression, suggesting that GDF3 and OCT4 have a functional relationship in pluripotent stem cells. A promoter-reporter assay revealed that the GDF3 promoter (−1721-Luc) activity was significantly activated by OCT4 in a dose-dependent manner. Moreover, the minimal promoter (−183-Luc) was sufficient for OCT4-mediated transcriptional activation and provided a potential binding site for the direct interaction with OCT4. Collectively, this study provides the evidence about the regulatory mechanism of GDF3 mediated by OCT4 in pluripotent EC cells.
Human testicular germ cell tumors (TGC Ts) originate from primordial germ cells, presumably due to some dysfunction in their normal development, and are classified as either non-seminomas, including teratocarcinomas with embryonic carcinoma (EC) components, or seminomas, which resemble undifferentiated primitive germ cells.1,2) Among them, undifferentiated EC and embryonic stem (ES) cells shares the high degree of similarity in gene expression pattern. In particular, OCT4, the major transcriptional regulator in stem cells, is highly expressed in EC and ES cells, reflecting its role as a functionally important regulator in pluripotent cells.1) OCT4 is a member of the octamer-binding subgroup of the Pit-Oct-Unc (POU) family of transcription factors, which bind to an octamer motif (consensus sequence, ATG CAA AT) using a bipartite POU DNA-binding domain.3,4) OCT4 is a master regulator of the induction and maintenance of cellular pluripotency and its expression must be tightly regulated since dysregulation of OCT4 expression leads to a loss of pluripotency and induces differentiation in ES cells.5–7) OCT4 is also expressed in histologically heterogeneous TGCTs.8–12) In recent studies, OCT4 enhances the migration and invasion of bladder cancer cells and also promotes colony formation of glioma cells.13,14) And also, OCT4 increases the transmigration capacity of melanoma cells, leading to increased invasiveness and aggressiveness.15)
Growth and differentiation factor 3 (GDF3), which is the member of the transforming growth factor β superfamily and a pluripotency-associated factor, is expressed in ES cells and the early embryo, and regulates both the pluripotency and differentiation potentials of stem cells.16–18) Moreover, GDF3 is suggested to be required during embryonic development due to the developmental abnormality of GDF3-null mice.19) Human GDF3 was first identified in EC cells and is also expressed in several cancer types, including TGCTs, breast carcinoma, and melanoma,12,19–22) suggesting it has a potential role in tumorigenesis. Moreover, GDF3 maps to chromosome 12p, which is a hotspot for teratocarcinoma.17) In fact, amplification of 12p is found in human neoplasms of esophageal and ovarian carcinomas.23–25) GDF3 seems to have contradictory roles in a cellular context-dependent manner. For example, GDF3 increases the proliferation of B16 myeloma, while it inhibits the proliferation of breast carcinoma MCF7 cells and promotes the neuronal differentiation of PC12 cells.21,22,26) Also, GDF3 induces differentiation-related genes, including several potent tumor suppressors, without affecting the proliferation capacity, and protects cancer stem cells from retinoic acid (RA)-induced apoptosis, suggesting it is a potent therapeutic target along with RA treatment.27)
Nevertheless, there is no evidence to explain how OCT4 is involved in the transcriptional regulation of GDF3 expression, even though both OCT4 and GDF3, as ES cell genes, are expressed in TGCTs as well as pluripotent stem cells.12,17,28) Germ cell-derived NCCIT cells are pluripotent stem cells of TGCTs, which have a phenotype intermediate between seminoma and EC cells.29) Therefore, we investigate whether OCT4 regulates the expression of GDF3 gene in NCCIT cells and provide the first evidence that OCT4 acts as a transcriptional activator for the oncogenic growth factor GDF3 expression in pluripotent EC cells.
MATERIALS AND METHODS
Cell Culture and DifferentiationNCCIT and HEK293T cells (American Type Cell Collection, Manassas, VA, U.S.A.) were cultured, as previously described.30) To induce differentiation, NCCIT cells were treated with 10 µM RA (Sigma, St. Louis, MO, U.S.A.) solubilized in pure ethyl alcohol and harvested at different time points (0, 2, 4, 6, 8, 10 d).
Plasmid ConstructionWe used luciferase reporter constructs containing the upstream region of the human GDF3 promoters (−1721-Luc, −183-Luc, −183*-Luc), which were previously reported by Park et al.30) We also used constructs harboring human full-length OCT4 and deletion variants [N-terminal domain (ND), ND to POU, and POU to C-terminal domain (CD)], which were previously reported by Lim et al.31) All cloned PCR products and reporter plasmids were verified by sequencing. To prepare the short hairpin RNA (shRNA) expression vector for RNA interference of OCT4, two double-stranded oligonucleotides were generated, based on a previous report32) for maximum silencing efficiency by annealing 5′-GAT CCGCGATCAAGCAGCGACTATGAAGCTTGATAGTCGCTGCTTGATCGCTTT TTT GGA AGC-3′ (sense) and 5′-GGC CGC TTC CAA AAA AGCGATCAAGCAGCGACTATCAAGCTTCATAGTCGCTGCTTGATCGCG-3′ (antisense). The OCT4 sense and antisense sequences are underlined. The hairpin loop structure containing the HindIII sequence is italicized. The resultant double-stranded oligos were inserted into the pGSH1-green fluorescent protein (pGSH1-GFP) shRNA vector (Genlantis, San Diego, CA, U.S.A.). The pGSH1-GFP-luciferase shRNA vector as a control was provided by the manufacturer (Genlantis).
Transient Transfection, RNA Preparation, and Quantitative Reverse Transcription (qRT)-PCRNCCIT cells were transfected with pGSH1-GFP-OCT4 shRNA or a FLAG-tagged OCT4 expression vector using the ViaFect™ transfection reagent (Promega, Madison, WI, U.S.A.). Total RNA was isolated from naïve, differentiated, or transfected NCCIT cells with TRIzol reagent (Invitrogen, La Jolla, CA, U.S.A.). cDNA was amplified using qRT-PCR mix (Bioneer, Daejeon, South Korea), as previously reported.30) The forward and reverse primers were as follows: GDF3, 5′-AGA CTT ATG CTA CGT AAA GGA GCT-3′ and 5′-CTT TGA TGG CAG ACA GGT TAA AGT A-3′; OCT4, 5′-CCC CTG GTG CCG TGA A-3′ and 5′-GCA AAT TGC TCG AGT TCT TTC TG-3′; β-actin, 5′-GCG GGA AAT CGT GCG TGA CAT T-3′ and 5′-GAT GGA GTT GAA GGT AGT TTC GTG-3′. Relative quantification of the expression levels was determined by the 2−ΔΔCT method.33)
Transient Transfection and Reporter AssaysNCCIT and HEK293T cells were transfected with a plasmid DNA mixture using the ViaFect™ transfection reagent (Promega) and harvested 48 h later. Reporter assays were performed, as described in previous reports.34,35)
Western Blot AnalysisNCCIT cells were treated with RA for different amounts of time (0, 2, 4, 6, 8, 10 d) and harvested, or transfected with OCT4-targeting shRNA or the Flag-tagged OCT4 expression vector using the ViaFect™ transfection reagent (Promega) and harvested 48 h later. Western blotting was performed as previously described36,37) with a minor modification. Membranes were blocked in 1% bovine serum albumin and incubated with anti-OCT4 polyclonal (1 : 2000, Cell Signaling Technology, Inc., Beverly, MA, U.S.A.), anti-GDF3 polyclonal (1 : 2000, Abcam, Cambridge, MA, U.S.A.), anti-Flag (1 : 10000, Medical & Biological Laboratories, Nagoya, Japan), and anti-β-actin monoclonal (1 : 2000, Santa Cruz Biotechnology, Dallas, TX, U.S.A.) antibodies. Each primary antibody-complexed membrane was further incubated with a horseradish peroxidase-conjugated secondary antibody (1 : 5000, Santa Cruz Biotechnology). Immunoreactive proteins were visualized by the West-Q Chemiluminescent Substrate Kit, Plus (GenDEPOT, CA, U.S.A.) and the LAS 4000 system (FUJIFILM, Tokyo, Japan).
Electrophoretic Mobility Shift Assay (EMSA)OCT4 was in vitro synthesized using the TNT Quick Coupled Transcription/Translation System (Promega) according to the manufacturer’s protocols. For the EMSA, complementary oligonucleotides were labeled using the Biotin 3′ End DNA Labeling Kit (Pierce Biotechnology, Inc., Rockford, IL, U.S.A.). For DNA-binding reactions, 8 µg of in vitro translated product was added to a 20 µL reaction containing 20 fmol of biotin-labeled oligonucleotide and 1 µg of poly(dI·dC) in binding buffer (10 mM Tris–HCl, pH 7.5, 500 mM NaCl, 1 mM ethylenediaminetetraacetic acid (EDTA), 10% glycerol, 1 mM dithiothreitol, and 1 nM phenylmethylsulfonyl fluoride). For competition experiments, reactions were incubated with excess amounts of an unlabeled double-stranded oligonucleotide probe. DNA–protein complexes were separated from the free probe on 6% non-denaturing polyacrylamide gels. The gels were transferred to a positive nylon membrane (GE Healthcare Life Sciences, United Kingdom) and signals were detected using a streptavidin-horseradish peroxidase conjugate (Thermo Scientific, Rockford, IL, U.S.A.). The oligonucleotide probe sequences used for the EMSA were as follows: WT, 5′-−107GAG CAA TTC ACACTTGATTATCTTACA TCA GAT A−74-3′; Mut, 5′-−107GAG CAA TTC ACcaTgGAggATCcTACATCA GATA−74-3′. The putative binding site is underlined, and the mutated sequences are shown in lower case.
Chromatin Immunoprecipitation (ChIP)The ChIP assay was performed as previously described30) with a minor modification. Precleared naïve NCCIT chromatin was incubated with a rabbit monoclonal anti-OCT4 antibody (Cell Signaling) or normal rabbit immunoglobulin G (IgG) (Cell Signaling) or without an antibody for more than 12 h. Immunoprecipitated DNA was then used as the template for real-time PCR using the GDF3 promoter-specific primers. The primer sequences were as follows: GDF3 promoter [designated as a, −624 to −422], 5′-GTC TAA TCC AGG GGT GTC CAA-3′ (forward) and 5′-GTC ACA TGA GGC TCA CGA TGG-3′ (reverse); GDF3 promoter [b, −254 to −163], 5′-ACT GCT GAC AAC CTC ACA TTG-3′ (forward) and 5′-CTC TTG CTC TAC TCT TCC AGG-3′ (reverse); GDF3 promoter [c, −144 to −47], 5′-CTC TTC ACA TCC AAT GGC CTT-3′ (forward) and 5′-TGC CCA ACA ATT CAG AGG CTG-3′ (reverse); and GDF3 promoter [d, +685 to +819], 5′-CTT CAT GCT TCC CTG CTG GTG-3′ (forward) and 5′-GAA TAG CTG GTG ACG GTG GCA-3′ (reverse).
Statistical AnalysisThe data were analyzed by the t-test and Duncan’s multiple range procedure for multiple comparisons using the SigmaPlot 11.2 program (Systat Software, Inc., San Jose, CA, U.S.A.). In all experiments, p<0.05 was considered statistically significant.
RESULTS AND DISCUSSION
Both OCT4 and GDF3, as ES cell markers, are expressed in seminomas, which morphologically resemble transformed primordial germ cells, and in breast cancer cells, suggesting they are valuable markers of tumorigenesis.20) In addition, shRNA-mediated OCT4 repression results in the down-regulation of GDF3 gene expression in human EC cells.38) We also previously reported that OCT4 and GDF3 are highly expressed in pluripotent human EC NCCIT cells and that this expression is reduced upon RA treatment in a time-dependent manner.30,36,37,39,40) However, there is no report concerning the interaction between OCT4 and GDF3 at the transcriptional level. Therefore, to investigate their transcriptional relationship in pluripotent germ cell-derived EC cells, we first examined the expression of GDF3 and OCT4 in NCCIT cells during RA-mediated differentiation. NCCIT cells were treated with 10 µM RA for different amounts of time (0, 2, 4, 6, 8, 10 d) and subjected to qRT-PCR and Western blot analyses (Fig. 1A). Both mRNA and protein levels of GDF3 and OCT4 significantly decreased during RA-mediated differentiation in a time-dependent manner. RA plays important roles in a variety of cellular processes such as differentiation, embryogenesis, and homeostasis.41) In particular, treatment of EC cells with RA represses the expression of several oncogenic factors and suppresses tumorigenesis, suggesting a link between enforced pluripotency and transformation.30) To further verify the direct relationship between GDF3 and OCT4 expression, shRNA-mediated knockdown and exogenous overexpression of OCT4 were performed using NCCIT cells, and relative expression was examined by qRT-PCR and Western blot analyses (Fig. 1B and Supplementary Fig. 1). The pGSH1-GFP-luciferase shRNA and Flag-tagged expression vectors were used as negative controls. At transcriptional levels, the expression of endogenous OCT4 and GDF3 mRNAs was significantly reduced by shRNA-mediated knockdown of OCT4 up to 80%, respectively, compared with control, whereas overexpression of exogenous OCT4 induced the expression of OCT4 and GDF3 (more than 8 and 2 folds, respectively). In addition, we also examined the protein levels by Western blotting and semi-quantitation using image J program (https://imagej.nih.gov/ij/). The OCT4 shRNA-mediated knockdown significantly reduced the expression of OCT4 and GDF3 to 70 and 50%, whereas exogenous OCT4 expression slightly increased the expression of OCT4 and GDF3 up to 3.6 and 2.2 folds, respectively, compared with control. It was also reported that RNA interference-mediated silencing of OCT4 reduces GDF3 expression in NCCIT cells.42) Therefore, these data suggest that OCT4 is involved in the regulation of GDF3 expression in NCCIT cells.
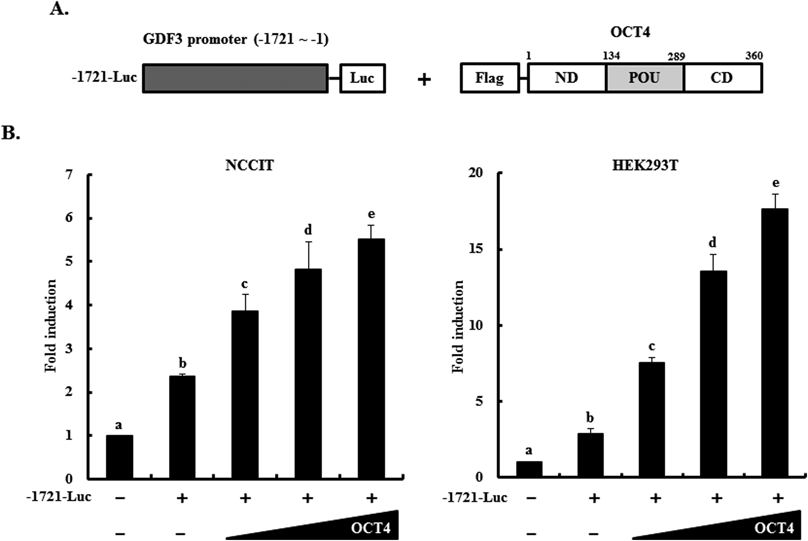
Three stem cell master regulators, OCT4, NANOG, and SRY (sex determining region Y)-box 2, have been reported to share target genes in ES and EC cells.28,43) Moreover, our previous study reported that the NANOG induces GDF3 expression.30) Based on the expression analysis data and previous studies, we examined whether OCT4 acts as a transcriptional activator of GDF3 expression. The GDF3 promoter-reporter vector (−1721-Luc) was co-transfected with the OCT4 expression vector in a dose-dependent manner into NCCIT and HEK293T cells (Fig. 2A). Promoter assays revealed that the GDF3 promoter (−1721-Luc) was significantly activated by dose-dependent OCT4 overexpression in both cell lines (Fig. 2B), suggesting that OCT4 acts as a transcriptional activator of GDF3 gene expression.
In our previous study,30) we found that the GDF3 minimal promoter region (−183 to −1 from the translation site) contains two NANOG-binding sites. A computational database (http://alggen.lsi.upc.es) revealed that one putative OCT4-binding site within the human GDF3 minimal promoter is located within the minimal promoter upstream region (−97 to −84, CAC TTG ATT ATC TTA), which, interestingly, overlaps with one of the NANOG-binding sites, despite the lack of octamer consensus sequences. Therefore, to examine whether OCT4 can interact with the putative binding site, wild type and mutant GDF3 minimal promoter constructs (−183-Luc and −183*-Luc, respectively) were co-transfected with the OCT4 expression vector into NCCIT and HEK293T cells (Fig. 3A). Transcriptional activity of the GDF3 minimal promoter (−183-Luc) was significantly activated by increasing amounts of the OCT4 gene expression vector in both NCCIT and HEK293T cells. By contrast, the mutant GDF3 promoter (−183*-Luc) with disruption of the putative OCT4-binding site that overlapped with the second NANOG-binding site was not activated even in the presence of exogenous OCT4. Taken together, the GDF3 minimal promoter region may contain the shared binding site for NANOG and OCT4, which is required for transcriptional activation of GDF3. In our previous study, we found two NANOG-binding sites, both of which are required for completing the activation of the GDF3 minimal promoter activity.30) To validate the data indicating OCT4-mediated induction of the GDF3 transcriptional activity through an interaction with the binding site that is shared with NANOG, we performed the promoter-reporter assay and confirmed that the mutant GDF3 promoter with disruption of the first NANOG-binding site was significantly activated when exogenous OCT4 was overexpressed (data not shown), supporting the idea that the putative OCT4-binding site within the GDF3 minimal promoter is effective for an interaction with OCT4 as well as NANOG. Further, we found that GDF3 minimal promoter has no significant effect on synergistic activation by co-transfection of OCT4 and NANOG (Supplementary Fig. 2). However, further studies may be required to determine the relationship between OCT4 and NANOG in their interaction with the same binding site within the GDF3 minimal promoter.
Next, we examined whether the forced transcriptional activation of the GDF3 minimal promoter is specific to OCT4. OCT4 contains two proline-rich domains within the N- and C-termini based on the POU DNA-binding domain, and both ND and CD play a transactivational role as transactivation domains.31) To examine the functional effect of OCT4 transactivation domains, constructs harboring full-length OCT4 and ND to POU, POU, and POU to CD of OCT4 were co-transfected with the GDF3 minimal promoter-reporter vector (−183-Luc) into HEK293T cells (Fig. 3B). OCT4 ND and CD were both independently capable of mediating the transcriptional activity of the GDF3 minimal promoter, supporting our previous finding and further indicating that both ND and CD mediate transcriptional activation of the GDF3 promoter.
To examine whether OCT4 can directly bind to the putative regulatory element in the GDF3 minimal promoter, an oligonucleotide spanning the binding site sequence was synthesized to perform an EMSA (Fig. 4A). In vitro synthesized OCT4 protein was added to the biotin-labeled probe, and protein–DNA complexes were observed (lane 1). By contrast, protein-DNA complexes were not observed with the mutant probe and were weak in the presence of an unlabeled competitor (lanes 2 and 3). These results strongly indicate that OCT4 directly binds to the putative binding site within the GDF3 minimal promoter region and upregulates GDF3 transcriptional activity. Therefore, we further verified whether OCT4 can bind to the region containing the putative binding element in native chromatin of undifferentiated NCCIT cells (Fig. 4B). Chromatin was prepared from undifferentiated NCCIT cells and subjected to the ChIP assay using an anti-OCT4 antibody. The enrichment of OCT4-bound chromatin was analyzed by real-time PCR using GDF3 promoter-specific primers to amplify the region (−144 to −47 depicted as c) spanning the putative binding site, and three other sites (−624 to −422 depicted as a; −254 to −163 depicted as b; and +685 to +819 depicted as d) as upstream and downstream negative controls. Region c (−144 to −47) of chromatin was significantly enriched, supporting that OCT4 can interact with the putative binding site within the GDF3 minimal promoter. It was reported that repression of OCT4 in ES cells results in marked loss of GDF3 transcripts as well as NANOG transcripts, both of which are located on the NANOG locus, and chromatin occupancy analysis revealed that OCT4 expression may be required to maintain chromatin structure within this locus.44)
Investigating the molecular regulatory mechanisms of stem cell transcription factors and their target gene(s) is crucial for identifying diagnostic biomarkers in tumorigenesis and molecular markers in stem cell pluripotency, which is mediated by coordinated transcription factor networks and multiple signaling molecules. Aberrant functioning of these stem cell regulators may influence malignant transformation and tumorigenesis.5) Therefore, this study demonstrates that OCT4 plays a role as a transcriptional activator for GDF3 transcription in pluripotent NCCIT cells and contributes the understanding of the molecular networks of stem cell regulators in germ cell-derived pluripotency and tumorigenesis.
Acknowledgment
This research was supported through the Next-Generation BioGreen21 Program (No. PJ01122202) funded by the Rural Development Administration, Republic of Korea.
Conflict of Interest
The authors declare no conflict of interest.
Supplementary Materials
The online version of this article contains supplementary materials.
REFERENCES
- 1) Sperger JM, Chen X, Draper JS, Antosiewicz JE, Chon CH, Jones SB, Brooks JD, Andrews PW, Brown PO, Thomson JA. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc. Natl. Acad. Sci. U.S.A., 100, 13350–13355 (2003).
- 2) Honecker F, Oosterhuis JW, Mayer F, Hartmann JT, Bokemeyer C, Looijenga LH. New insights into the pathology and molecular biology of human germ cell tumors. World J. Urol., 22, 15–24 (2004).
- 3) Klemm JD, Pabo CO. Oct-1 POU domain–DNA interactions: cooperative binding of isolated subdomains and effects of covalent linkage. Genes Dev., 10, 27–36 (1996).
- 4) Jerabek S, Merino F, Schöler HR, Cojocaru V. OCT4: dynamic DNA binding pioneers stem cell pluripotency. Biochim. Biophys. Acta, 1839, 138–154 (2014).
- 5) Kashyap V, Rezende NC, Scotland KB, Shaffer SM, Persson JL, Gudas LJ, Mongan NP. Regulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the NANOG, OCT4, and SOX2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAs. Stem Cells Dev., 18, 1093–1108 (2009).
- 6) Tysnes BB. Tumor-initiating and -propagating cells: cells that we would like to identify and control. Neoplasia, 12, 506–515 (2010).
- 7) Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat. Genet., 24, 372–376 (2000).
- 8) Looijenga LH, Stoop H, de Leeuw HP, de Gouveia Brazao CA, Gillis AJ, van Roozendaal KE, van Zoelen EJ, Weber RF, Wolffenbuttel KP, van Dekken H, Honecker F, Bokemeyer C, Perlman EJ, Schneider DT, Kononen J, Sauter G, Oosterhuis JW. POU5F1 (OCT3/4) identifies cells with pluripotent potential in human germ cell tumors. Cancer Res., 63, 2244–2250 (2003).
- 9) Almstrup K, Hoei-Hansen CE, Wirkner U, Blake J, Schwager C, Ansorge W, Nielsen JE, Skakkebaek NE, Rajpert-De Meyts E, Leffers H. Embryonic stem cell-like features of testicular carcinoma in situ revealed by genome-wide gene expression profiling. Cancer Res., 64, 4736–4743 (2004).
- 10) Clark AT. The stem cell identity of testicular cancer. Stem Cell Rev., 3, 49–59 (2007).
- 11) Santagata S, Ligon KL, Hornick JL. Embryonic stem cell transcription factor signatures in the diagnosis of primary and metastatic germ cell tumors. Am. J. Surg. Pathol., 31, 836–845 (2007).
- 12) Gopalan A, Dhall D, Olgac S, Fine SW, Korkola JE, Houldsworth J, Chaganti RS, Bosl GJ, Reuter VE, Tickoo SK. Testicular mixed germ cell tumors: a morphological and immunohistochemical study using stem cell markers, OCT3/4, SOX2 and GDF3, with emphasis on morphologically difficult-to-classify areas. Mod. Pathol., 22, 1066–1074 (2009).
- 13) Chang CC, Shieh GS, Wu P, Lin CC, Shiau AL, Wu CL. Oct-3/4 expression reflects tumor progression and regulates motility of bladder cancer cells. Cancer Res., 68, 6281–6291 (2008).
- 14) Du Z, Jia D, Liu S, Wang F, Li G, Zhang Y, Cao X, Ling EA, Hao A. Oct4 is expressed in human gliomas and promotes colony formation in glioma cells. Glia, 57, 724–733 (2009).
- 15) Borrull A, Ghislin S, Deshayes F, Lauriol J, Alcaide-Loridan C, Middendorp S. Nanog and Oct4 overexpression increases motility and transmigration of melanoma cells. J. Cancer Res. Clin. Oncol., 138, 1145–1154 (2012).
- 16) McPherron AC, Lee SJ. GDF-3 and GDF-9: two new members of the transforming growth factor-beta superfamily containing a novel pattern of cysteines. J. Biol. Chem., 268, 3444–3449 (1993).
- 17) Clark AT, Rodriguez RT, Bodnar MS, Abeyta MJ, Cedars MI, Turek PJ, Firpo MT, Reijo Pera RA. Human STELLAR, NANOG, and GDF3 genes are expressed in pluripotent cells and map to chromosome 12p13, a hotspot for teratocarcinoma. Stem Cells, 22, 169–179 (2004).
- 18) Levine AJ, Brivanlou AH. GDF3, a BMP inhibitor, regulates cell fate in stem cells and early embryos. Development, 133, 209–216 (2006).
- 19) Caricasole AA, van Schaik RH, Zeinstra LM, Wierikx CD, van Gurp RJ, van den Pol M, Looijenga LH, Oosterhuis JW, Pera MF, Ward A, de Bruijn D, Kramer P, de Jong FH, van den Eijnden-van Raaij AJ. Human growth-differentiation factor 3 (hGDF3): developmental regulation in human teratocarcinoma cell lines and expression in primary testicular germ cell tumours. Oncogene, 16, 95–103 (1998).
- 20) Ezeh UI, Turek PJ, Reijo RA, Clark AT. Human embryonic stem cell genes OCT4, NANOG, STELLAR, and GDF3 are expressed in both seminoma and breast carcinoma. Cancer, 104, 2255–2265 (2005).
- 21) Ehira N, Oshiumi H, Matsumoto M, Kondo T, Asaka M, Seya T. An embryo-specific expressing TGF-β family protein, growth-differentiation factor 3 (GDF3), augments progression of B16 melanoma. J. Exp. Clin. Cancer Res., 29, 135 (2010).
- 22) Li Q, Ling Y, Yu L. GDF3 inhibits the growth of breast cancer cells and promotes the apoptosis induced by Taxol. J. Cancer Res. Clin. Oncol., 138, 1073–1079 (2012a).
- 23) Yen CC, Chen YJ, Chen JT, Hsia JY, Chen PM, Liu JH, Fan FS, Chiou TJ, Wang WS, Lin CH. Comparative genomic hybridization of esophageal squamous cell carcinoma: correlations between chromosomal aberrations and disease progression/prognosis. Cancer, 92, 2769–2777 (2001).
- 24) Schraml P, Schwerdtfeger G, Burkhalter F, Raggi A, Schmidt D, Ruffalo T, King W, Wilber K, Mihatsch MJ, Moch H. Combined array comparative genomic hybridization and tissue microarray analysis suggest PAK1 at 11q13.5-q14 as a critical oncogene target in ovarian carcinoma. Am. J. Pathol., 163, 985–992 (2003).
- 25) Korkola JE, Houldsworth J, Chadalavada RS, Olshen AB, Dobrzynski D, Reuter VE, Bosl GJ, Chaganti RS. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res., 66, 820–827 (2006).
- 26) Li Q, Liu X, Wu Y, An J, Hexige S, Ling Y, Zhang M, Yang X, Yu L. The conditioned medium from a stable human GDF3-expressing CHO cell line, induces the differentiation of PC12 cells. Mol. Cell. Biochem., 359, 115–123 (2012b).
- 27) Tykwinska K, Lauster R, Knaus P, Rosowski M. Growth and differentiation factor 3 induces expression of genes related to differentiation in a model of cancer stem cells and protects them from retinoic acid-induced apoptosis. PLoS ONE, 8, e70612 (2013).
- 28) Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK, Melton DA, Jaenisch R, Young RA. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell, 122, 947–956 (2005).
- 29) Damjanov I, Horvat B, Gibas Z. Retinoic acid-induced differentiation of the developmentally pluripotent human germ cell tumor-derived cell line, NCCIT. Lab. Invest., 68, 220–232 (1993).
- 30) Park SW, Lim HY, Do HJ, Sung B, Huh SH, Uhm SJ, Song H, Chung HJ, Kim JH, Kim NH, Kim JH. Regulation of human growth and differentiation factor 3 gene expression by NANOG in human embryonic carcinoma NCCIT cells. FEBS Lett., 586, 3529–3535 (2012).
- 31) Lim HY, Do HJ, Lee WY, Kim DK, Seo HG, Chung HJ, Park JK, Chang WK, Kim JH, Kim JH. Implication of human OCT4 transactivation domains for self-regulatory transcription. Biochem. Biophys. Res. Commun., 385, 148–153 (2009).
- 32) Rodriguez RT, Velkey JM, Lutzko C, Seerke R, Kohn DB, O’Shea KS, Firpo MT. Manipulation of OCT4 levels in human embryonic stem cells results in induction of differential cell types. Exp. Biol. Med. (Maywood), 232, 1368–1380 (2007).
- 33) Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods, 25, 402–408 (2001).
- 34) Yang HM, Do HJ, Oh JH, Kim JH, Choi SY, Cha KY, Chung HM, Kim JH. Characterization of putative cis-regulatory elements that control the transcriptional activity of the human Oct4 promoter. J. Cell. Biochem., 96, 821–830 (2005).
- 35) Yang HM, Do HJ, Kim DK, Park JK, Chang WK, Chung HM, Choi SY, Kim JH. Transcriptional regulation of human Oct4 by steroidogenic factor-1. J. Cell. Biochem., 101, 1198–1209 (2007).
- 36) Park SW, Do HJ, Ha WT, Han MH, Park KH, Song H, Kim NH, Kim JH. Transcriptional activation of OCT4 by the ETS transcription factor PEA3 in NCCIT human embryonic carcinoma cells. FEBS Lett., 588, 3129–3136 (2014a).
- 37) Park SW, Do HJ, Ha WT, Han MH, Yang HM, Lee SH, Song H, Kim NH, Kim JH. Transcriptional regulation of OCT4 by the ETS transcription factor ESE-1 in NCCIT human embryonic carcinoma cells. Biochem. Biophys. Res. Commun., 450, 984–990 (2014b).
- 38) Giuliano CJ, Kerley-Hamilton JS, Bee T, Freemantle SJ, Manickaratnam R, Dmitrovsky E, Spinella MJ. Retinoic acid represses a cassette of candidate pluripotency chromosome 12p genes during induced loss of human embryonal carcinoma tumorigenicity. Biochim. Biophys. Acta, 1731, 48–56 (2005).
- 39) Sung B, Do HJ, Park SW, Huh SH, Oh JH, Chung HJ, Kang MJ, Kim JH, Kim NH, Kim JH. Regulation of OCT4 gene expression by liver receptor homolog-1 in human embryonic carcinoma cells. Biochem. Biophys. Res. Commun., 427, 315–320 (2012).
- 40) Park SW, Do HJ, Ha WT, Han MH, Song H, Uhm SJ, Chung HJ, Kim JH. Differential expression of ETS family transcription factors in NCCIT human embryonic carcinoma cells upon retinoic acid-induced differentiation. Biol. Pharm. Bull., 37, 659–665 (2014c).
- 41) Mendelsohn C, Ruberte E, Chambon P. Retinoid receptors in vertebrate limb development. Dev. Biol., 152, 50–61 (1992).
- 42) Jung M, Peterson H, Chavez L, Kahlem P, Lehrach H, Vilo J, Adjaye J. A data integration approach to mapping OCT4 gene regulatory networks operative in embryonic stem cells and embryonal carcinoma cells. PLoS ONE, 5, e10709 (2010).
- 43) Greber B, Lehrach H, Adjaye J. Silencing of core transcription factors in human EC cells highlights the importance of autocrine FGF signaling for self-renewal. BMC Dev. Biol., 7, 46 (2007).
- 44) Levasseur DN, Wang J, Dorschner MO, Stamatoyannopoulos JA, Orkin SH. Oct4 dependence of chromatin structure within the extended Nanog locus in ES cells. Genes Dev., 22, 575–580 (2008).