Abstract
The pathogenesis of Alzheimer’s disease (AD) has been linked to the deficiency of neurotransmitter acetylcholine (ACh) in the brain, and the main treatment strategy for improving AD symptoms is the inhibition of acetylcholinesterase (AChE) activity. In the present study, we aimed to identify potent AChE inhibitors from Cinnamomum loureirii extract via bioassay-guided fractionation. We demonstrated that the most potent AChE inhibitor present in the C. loureirii extract was 2,4-bis(1,1-dimethylethyl)phenol. To confirm the antiamnesic effects of the ethanol extract of C. loureirii, mice were intraperitoneally injected with the neurotoxin trimethyltin (2.5 mg/kg) to induce cognitive dysfunction, and performance in the Y-maze and passive avoidance tests was assessed. Treatment with C. loureirii extract significantly improved performance in both behavioral tests, suggesting that this extract may be neuroprotective and therefore beneficial in preventing or ameliorating the degenerative processes of AD, potentially by restoring cholinergic function.
The pathophysiology of Alzheimer’s disease (AD), a type of dementia in the elderly, is highly complex. This progressive degenerative disorder was first discovered by Dr. Alois Alzheimer in 1907 and is characterized by incapacitating memory and language losses and impairments in cognitive and behavioral functions. A neuropathological diagnosis of AD includes deposition of extracellular β-amyloid (Aβ) plaques in the cerebral cortex and hippocampus areas, accumulation of intracellular neurofibrillary tangles of abnormally phosphorylated τ, astrocytic gliosis, inflammatory cascades, and degeneration of basal forebrain cholinergic neurons.1–3)
Cholinergic neurotransmission in the central nervous system (CNS) involves release of acetylcholine (ACh) from presynaptic terminals, binding of ACh to cognate receptors on postsynaptic cells, and downstream signal transduction.4) To date, the most significant and best approach for the prevention and treatment of AD is the inhibition of acetylcholinesterase (AChE).5) AChEs are present in the synaptic cleft of cholinergic synapses and rapidly degrade ACh into choline and acetic acid.6) These enzymes decrease the concentration of ACh in the synapse and downregulate ACh-induced signaling pathways.7) Currently, many studies have focused on the use of AChE inhibitors (AChEIs) to reverse abnormally elevated AChE activity in patients with AD, thereby restoring ACh levels and ameliorating AD.8,9)
Traditionally, various edible plants have been used as medicinal remedies in Asian countries, including Korea, since they are rich sources of diverse secondary metabolites, often called phytochemicals. Notably, Cinnamomum loureirii is one of the most widely used herbal medicines in Korea. The inner bark of C. loureirii is obtained from the trees belonging to the family Lauraceae10) and is often used as a spice and flavoring agent. C. loureirii contains large amounts of bioactive molecules, including essential oils, tannin, mucus, and coumarins.11) Previous studies demonstrated that C. loureirii might be effective in the treatment of gastritis, dyspepsia, inflammatory diseases, and blood circulation disturbances and may possess anti-pyretic, analgesic, anti-allergic, and anti-ulcerogenic effects.12,13) To the best of our knowledge, however, no study has investigated the inhibitory effects of C. loureirii on AChE activity utilizing in vitro and in vivo models.
The aim of the present study was to identify AChEIs from C. loureirii. In order to isolate a candidate AChEI, the ethanol extract of C. loureirii was subjected to solvent partitioning, open column chromatography, TLC, and HPLC.14,15) The chemical structure of the purified active compound was determined using GC-MS and NMR spectroscopy. In addition, the memory enhancing effect of C. loureirii in trimethyltin (TMT)-induced cognitive dysfunction was assessed using two behavioral tests, the Y-maze test and passive avoidance test.
MATERIALS AND METHODS
MaterialsRoswell Park Memorial Institute (RPMI)-1640 medium, heat-inactivated horse serum (HS), fetal bovine serum (FBS), and antibiotic-antimycotic were purchased from Gibco-Invitrogen (Grand Island, NY, U.S.A.). Acetylthiocholine iodide, 5,5′-dithiobis-(2-nitro)benzoic acid (DTNB), 1,5-bis-(4-allydimethyllammoniumphenyl)-pentane-3-one dibromide (BW 284c51), tacrine (9-amino-1,2,3,4-tetrahydroacridine), dimethyl sulfoxide (DMSO), and TMT chloride were purchased from Sigma (St. Louis, MO, U.S.A.). All other chemicals used were of analytical grade purity unless otherwise specified.
Cell CultureRat pheochromocytoma (PC12; ATC C, Manassas, VA, U.S.A.) cells were cultured in RPMI-1640 medium supplemented with 10% HS (v/v), 5% FBS (v/v), and 1% antibiotic-antimycotic (v/v). Cultures were maintained at 37°C with 5% CO2 and water saturation. When the culture was 80–90% confluent, the cells were subcultured. The medium was replaced every 2 to 3 d.
Measurement of AChE ActivityAChE activity was determined using the modified spectrophotometric method of Ellman et al.16) For the enzyme source, PC12 cells were homogenized in Tris–HCl buffer [20 mM Tris–HCl (pH 7.5) containing 150 mM NaCl, 10 mM MgCl, and 0.5% Triton X-100] and then centrifuged at 10000×g for 15 min. The supernatant was used as the enzyme source, and ACh iodide was used as the reaction substrate. DTNB was selected to quantify the thiocholine produced from the enzymatic hydrolysis of AChE. Briefly, 10 µL of each sample was mixed with 10 µL of enzyme solution and added to 70 µL of reaction mixture (50 mM sodium phosphate buffer [pH 8.0] containing 0.5 mM ACh iodide and 1 mM DTNB). This reaction mix was incubated at 37°C for 15 min. The final enzyme reactions were monitored at a wavelength of 405 nm using a 96-well microplate reader (Model 550; Bio-Rad, Hercules, CA, U.S.A.). Tacrine was utilized as a reference standard in order to assess the AChE inhibitory effects of each group.
Extraction of C. loureiriiDried C. loureirii (4 kg) was purchased from Gyung-dong market, an oriental medicine store in Seoul, South Korea. The plant material was finely pulverized to powder, and 4 kg of the powder was blended with five volumes of ethanol (100 L) by shaking for 24 h at room temperature. The resulting supernatant was filtered through Whatman ashless filter paper No. 42 (GE Healthcare Life Sciences, Chalfont St. Giles, U.K.), and the residue was extracted again with fresh ethanol. This procedure was repeated five times, and the solution was completely dried using a rotary evaporator at 39°C. The percentage yield of C. loureirii extract was 26.1% (w/w).
Isolation of the AChE Inhibitor from C. loureiriiThe C. loureirii ethanol extract (998 g) was dissolved in distilled water (1560 mL), and then respectively partitioned with n-hexane (4680 mL), chloroform (4680 mL), and ethyl acetate (4680 mL), repeated three times, for 24 h. Of nine fractions, the fraction with the most pronounced inhibitory effect on AChE was chosen and subjected to 33 sub-fractions through gradient elution with a mixture of the solvents (chloroform–ethanol; 100 : 0, 90 : 10, 80 : 20, 70 : 30, 60 : 40, 50 : 50, 40 : 60, 30 : 70, 20 : 80, 10 : 90, 0 : 100, v/v, respectively, repeated three times per each elution) using gradient-elution silica-gel open column chromatography. The most potent fraction was subjected to TLC and then visualized under visible and ultraviolet light (254 and 365 nm, Vilber Lourmatm, Collégien, France). Each band separated on TLC was scraped off and extracted with ethanol and/or chloroform for the enzyme assay. Amongst all TLC bands scraped and examined, a band representing the most potent inhibitory effects against AChE was selected and subjected to HPLC analysis. Waters HPLC system (Waters Co., Milford, MA, U.S.A.) equipped with a 2690 pump, a 2996 photodiode array detector, and Empower2 software was utilized. The analytical column was a C18 µ-Bondapak™ reverse phase column (Waters Co., 3.9×300 mm, 125 Å, 10 µm). Mobile phases were employed for gradient elution with water (A) and HPLC-grade ethanol (B) under the following conditions: initially 100% A for 10 min, linear to 0% A over 50 min, followed by isocratic elution at 0% A for an additional 10 min, to 100% A over 10 min, and 100% A isocratic for 10 min at a flow rate of 0.5 mL/min. The structure of the purified active compound was then determined by GC-MS and 13C/1H-NMR spectroscopic techniques (Korea Basic Science Institute, Western Seoul Center).
AnimalsMale Institute of Cancer Research (ICR) mice (5-week-old) were purchased from DBL Co. (Chungbuk, South Korea). Mice were housed in groups of eight and maintained under a 12 h light/dark cycle at a constant temperature of 23±1°C and relative humidity of 55%. The C. loureirii ethanol extract was supplemented in commercial diet (RodFeed; DBL Co., Chungbuk, South Korea) at concentrations of 400, 800, and 1200 mg/kg body weight (wt). All animals were allowed free access to diets and water. All experimental procedures were approved by the Animal Care and Use Committee of Korea University and performed in compliance with the guideline thereof.
TMT InjectionAfter animals received the diets for 3 weeks, mice of the TMT group and sample groups received a single intraperitoneal injection of TMT. The TMT was dissolved in 0.85% (v/v) sodium chloride, and dosage was determined previously, as described (2.5 mg of TMT/kg body wt).17) The negative control group was injected with sodium chloride solution alone. The behavioral tests were conducted 1–2 d after TMT injection.
Y-Maze TestThe maze was made of black plastic with three equal angled arms (33 cm long×10 cm wide×15 cm high) in a Y shape. Each mouse was initially placed within an arm and allowed to explore the maze over 8 min. The sequence and number of arm entries visited were monitored and manually recorded. The spontaneous alternation behavior score (%) was calculated by dividing the possible alternation by the total number of arm entries minus 2, multiplied by 100. The entrance numbers per trial was used as an indicator of locomotor activity.18)
Passive Avoidance TestThe test was carried out in a chamber with two compartments (illuminated/dark) with a stainless grid floor that had electrical conductibility. For the acquisition trials, the mouse was placed in the illuminated compartment. When a mouse entered the dark chamber, an electrical foot shock (0.5 mA, 1 s) was applied through the grid floor. In the retention trial, the same procedure was performed 24 h after training, and the mouse was again placed in the illuminated compartment. The time required to enter the dark chamber was recorded up to 5 min.19)
Statistical AnalysisData are presented as mean±standard deviation (S.D.). Statistical analyses were performed using the Statistical Analysis System software package (Cary, NC, U.S.A.). Significant differences were calculated by one-way ANOVA followed by the Scheffe’s multiple range test. A p value less than 0.05 was considered statistically significant.
RESULTS
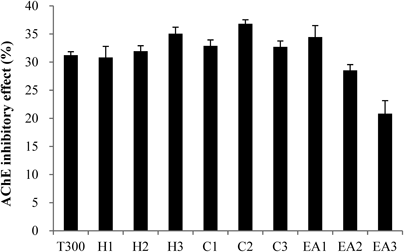
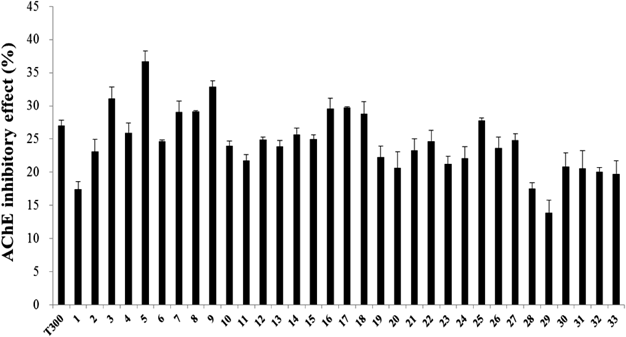
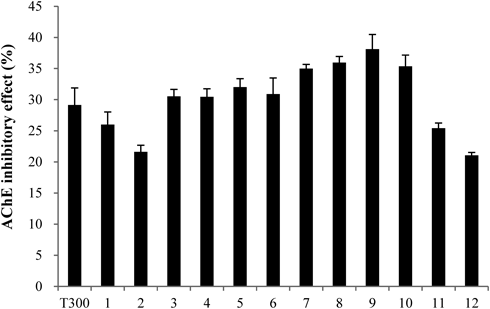
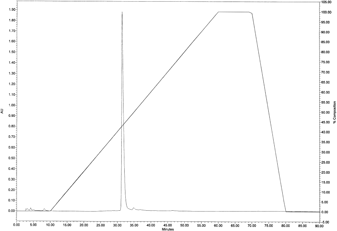
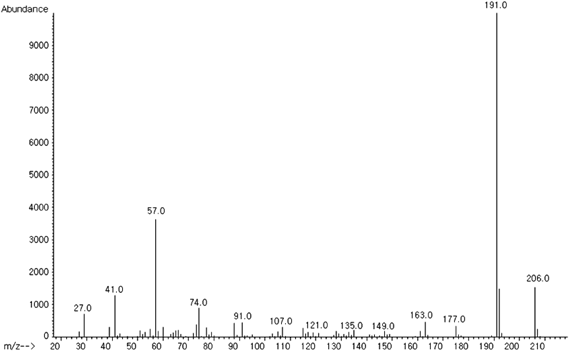
Purification of a Candidate AChEI from C. loureirii ExtractThe C. loureirii ethanol extract (total weight: 998 g) was subjected to liquid–liquid partition. Among the nine fractions obtained, the second fraction of chloroform layer was selected for further isolation since it exhibited the highest inhibitory activity against AChE relative to the other layers (Fig. 1). This chloroform extract (total weight: 11.9 g) was then partitioned into 33 fractions by open column chromatography, and these fractions were tested for their ability to inhibit AChE. Among all the fractions, the fifth fraction showed the highest inhibitory effect on AChE (Fig. 2). This fraction (total weight: 4.5 g) was then subjected to TLC to separate the active compound. The bands clustered near the intermediate range of Rf value, i.e., 0.09–0.92, and these bands were extracted with ethanol and/or chloroform. Of all the TLC bands scraped, the ninth band (Rf value=0.64) showed the most effective inhibitory effect against AChE activity (Fig. 3), and hence, it subsequently was subjected to HPLC (total weight: 128 mg). As shown in Fig. 4, a significant single peak was seen at 31.43 min of the chromatogram. The single peak was collected at the retention time depending on the chromatography seen, and the final yield of the isolated component from C. loureirii (4 kg) was about 7 mg.
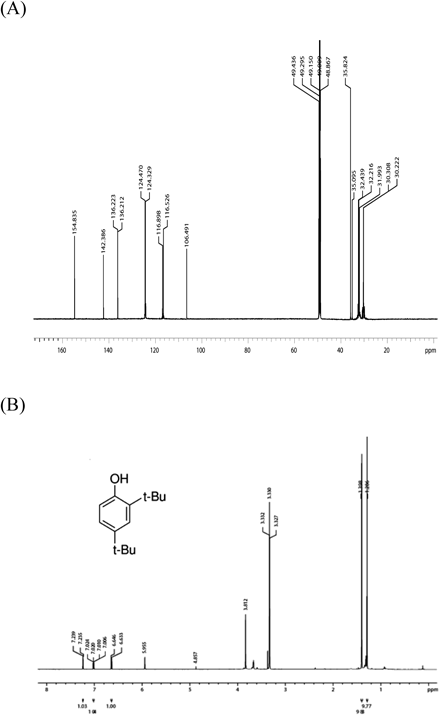
The structure of the candidate AChEI was elucidated by GC-MS and 13C/1H-NMR and was identified as a 2,4-bis(1,1-dimethylethyl)phenol (BP) (m/z=206), which is a natural polyphenol with a phenolic structure (Figs. 5, 6).
The Inhibitory Effect of BP on AChEThe AChE inhibitory activity of BP was tested. As shown, AChE inhibitory activity was increased by the BP with a dose-dependent manner (Table 1).
Table 1. Inhibition of Acetylcholinesterase (AChE) Activity under Different Concentrations of 2,4-Bis(1,1-dimethylethyl)phenol
| Concentration of BP | AChE inhibitory activity (%) |
|---|
| 10 mM | 17.12±1.33 |
| 20 mM | 21.19±1.39 |
| 40 mM | 30.45±3.41 |
The value (%) of AChE activity for each fraction was calculated relative to control activity (100%). Each value represents the mean±standard deviation (S.D.) (n=4).
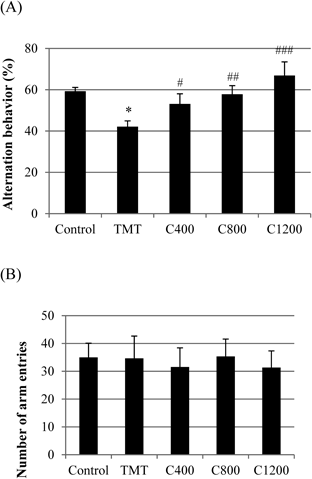
The effect of C. loureirii extract on immediate spatial working memory was investigated using the Y-maze test (Fig. 7A). The TMT-treated group showed a significant deficit (17.2% decrease) in spontaneous alternation behavior compared to the negative control group. However, dietary supplementation of C. loureirii extract significantly increased the alternation behavior compared with that of the TMT-treated group. Even though the C. loureirii extract efficiently restored spatial working memory in the Y-maze test, the total number of arm entries was not significantly different among all the groups, suggesting that TMT and the samples affected nonspecific processes, including arousal, attention, or sensory motor functions (Fig. 7B).
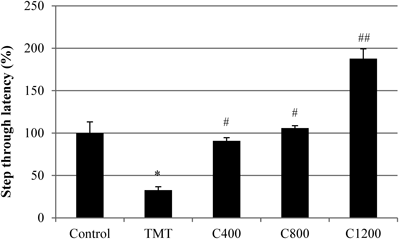
Learning and short-term memory impairment of TMT-treated mice were assessed by the passive avoidance test (Fig. 8). The step-through latency of the TMT group was 67.2% less than that of the negative control group (i.e., TMT-free control group), and the administration of C. loureirii extract significantly improved this reduction in TMT treated animals. Importantly, TMT treatment impaired retention of the single-trial passive avoidance test. Furthermore, mice treated with TMT and samples did not exhibit any significant differences in body or brain weight and acute liver toxicity when compared to control mice (data not shown). Thus, improvements in memory performance with C. loureirii were not due to changes in weight or levels of serum aminotransferases.
DISCUSSION
In order to isolate a potent AChEI from natural sources, often considered more safe, the ethanol extract of C. loureirii was subjected to bioassay guided purification: solvent partitioning, open column chromatography, TLC, and HPLC.17) The structure of a candidate compound was elucidated using GC-MS and NMR; the structure of the purified compound was determined as BP. In our subsequent in vitro assay, this constituent inhibited the AChE activity with a dose dependent manner. Previously, it was reported that this phenolic compound is present in wild plants and mushrooms.20–22) Of note, BP has been widely used as a raw material for phosphite antioxidants and as an intermediate for the preparation of ultraviolet stabilizers as well.23) According to previous reports, BP is an antioxidant with remarkable cytotoxic activity against cancer cells,24,25) and it has been shown to decrease the degree of low-density lipoprotein (LDL)-oxidation.26) Moreover, BP was reported to be the active compound in the ethanol extract of Ipomoea batatas that was shown to be protective against Aβ-induced neurotoxicity.27)
TMT, a potent neurotoxicant, has been used to induce experimental models of AD.28) The administration of TMT in rodents has been reported to cause neuropathological and behavioral alternations, including aggression, tremor, and hyperexcitability.29) These changes were correlated with degeneration of cholinergic axons and changes in ACh levels and their receptors.30) Kobayashi et al. founded that TMT toxicity causes a decrease of ACh release in cerebral tissue.31) These changes might be due to hippocampal loss of neurons, leading to deficits in learning and memory. As shown in Figs. 7A and 8, supplementation of C. loureirii extract to TMT-treated mice attenuated TMT-induced impairment in a dose-dependent manner. Thus, C. loureirii extract possibly improves learning and memory function in TMT-treated animals by inhibiting AChE thereby restoring ACh levels.
Co-administration of C. loureirii extract and TMT had no effect on the body weight or brain weight of mice. In addition, we found that C. loureirii extract was not hepatotoxic and exerted no adverse effect on liver function. Future investigation might be needed to examine the effect of exposure to BP on TMT-induced memory deficiency in vivo; further, it would be interesting to compare the biological potency between crude C. loureirii extract and the purified constituent, BP.
This study demonstrated that C. loureirii extract inhibited AChE in vitro and was neuroprotective against cognitive dysfunction resulting from TMT exposure. Overall, C. loureirii, including bioactive polyphenol, is a potent source of AChEIs that might be beneficial in slowing or preventing the degenerative processes of AD by restoring the cholinergic basal forebrain system.
Acknowledgments
This research was supported by High Value-Added Food Technology Development Program, Ministry of Agriculture, Food and Rural Affairs, Republic of Korea.
Conflict of Interest
The authors declare no conflict of interest.
REFERENCES
- 1) Geula C. Abnormalities of neural circuitry in Alzheimer’s disease: hippocampus and cortical cholinergic innervation. Neurology, 51 (Suppl. 1), S18–S29, discussion, S65–S67 (1998).
- 2) McGeer PL, McGeer EG. Inflammation, autotoxicity and Alzheimer’s disease. Neurobiol. Aging, 22, 799–809 (2001).
- 3) Lee VM, Goedert M, Trojanowski JQ. Neurodegenerative tauopathies. Annu. Rev. Neurosci., 24, 1121–1159 (2001).
- 4) Mesulam MM, Guillozet A, Shaw P, Levey A, Duysen EG, Lockridge O. Acetylcholinesterase knockouts establish central cholinergic pathways and can use butyrylcholinesterase to hydrolyze acetylcholine. Neuroscience, 110, 627–639 (2002).
- 5) Bartus RT. On neurodegenerative diseases, models, and treatment strategies: lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp. Neurol., 163, 495–529 (2000).
- 6) Fujii T, Mori Y, Tominaga T, Hayasaka I, Kawashima K. Maintenance of constant blood acetylcholine content before and after feeding in young chimpanzees. Neurosci. Lett., 227, 21–24 (1997).
- 7) DeKosky ST, Harbaugh RE, Schmitt FA, Bakay RA, Chui HC, Knopman DS, Reeder TM, Shetter AG, Senter HJ, Markesbery WR, Intraventricular Bethanecol Study Group. Cortical biopsy in Alzheimer’s disease: diagnostic accuracy and neurochemical, neuropathological, and cognitive correlations. Intraventricular Bethanecol Study Group. Ann. Neurol., 32, 625–632 (1992).
- 8) Nordberg A, Svensson AL. Cholinesterase inhibitors in the treatment of Alzheimer’s disease: a comparison of tolerability and pharmacology. Drug Saf., 19, 465–480 (1998).
- 9) Loizzo MR, Tundis R, Menichini F, Menichini F. Natural products and their derivatives as cholinesterase inhibitors in the treatment of neurodegenerative disorders: an update. Curr. Med. Chem., 15, 1209–1228 (2008).
- 10) Li R, Wang Y, Jiang ZT, Jiang S. Chemical composition of the essential oils of Cinnamomum loureirii Nees. From China obtained by hydrodistillation and microwave-assisted hydrodistillation. J. Essent. Oil Res., 22, 129–131 (2010).
- 11) Burdock G. Fenaroli’s Handbook of Flavor Ingredients, Fifth Edition, CRC Press, Boca Raton (2005).
- 12) Kurokawa M, Kumeda CA, Yamamura J, Kamiyama T, Shiraki K. Antipyretic activity of cinnamyl derivatives and related compounds in influenza virus-infected mice. Eur. J. Pharmacol., 348, 45–51 (1998).
- 13) Zhu ZP, Zhang MF, Shen YQ, Chen GJ. Pharmacological study on spleen-stomach warming and analgesic action of Cinnamomum cassia Presl. Zhongguo Zhong Yao Za Zhi, 18, 553–557, 514–515 (1993).
- 14) Sasidharan S, Chen Y, Saravanan D, Sundram KM, Yoga Latha L. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med., 8, 1–10 (2011).
- 15) Ong ES. Extraction methods and chemical standardization of botanicals and herbal preparations. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 812, 23–33 (2004).
- 16) Ellman GL, Courtney KD, Andres V Jr, Feather-Stone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7, 88–95 (1961).
- 17) Kim CR, Choi SJ, Oh SS, Kwon YK, Lee NY, Park GG, Kim YJ, Heo HJ, Jun WJ, Park CS, Shin DH. Rubus coreanus Miquel inhibits acetylcholinesterase activity and prevents cognitive impairment in a mouse model of dementia. J. Med. Food, 16, 785–792 (2013).
- 18) Kim MJ, Choi SJ, Lim ST, Kim HK, Heo HJ, Kim EK, Jun WJ, Cho HY, Kim YJ, Shin DH. Ferulic acid supplementation prevents trimethyltin-induced cognitive deficits in mice. Biosci. Biotechnol. Biochem., 71, 1063–1068 (2007).
- 19) Heo HJ, Kim MJ, Lee JM, Choi SJ, Cho HY, Hong B, Kim HK, Kim E, Shin DH. Naringenin from Citrus junos has an inhibitory effect on acetylcholinesterase and a mitigating effect on amnesia. Dement. Geriatr. Cogn. Disord., 17, 151–157 (2004).
- 20) Rana VS, Blazquez MA. Chemical constituents of Gynura cusimbua aerial parts. J. Essent. Oil Res., 19, 21–22 (2007).
- 21) Xuan TD, Toyama T, Fukuta M, Khanh TD, Tawata S. Chemical interaction in the invasiveness of cogongrass (Imperata cylindrica (L.) Beauv.). J. Agric. Food Chem., 57, 9448–9453 (2009).
- 22) Zhou J, Feng T, Ye R. Differentiation of eight commercial mushrooms by electronic nose and gas chromatography-mass spectrometry. J. Sens., 2015, 1–14 (2015).
- 23) Nerín C, Fernández C, Domeño C, Salafranca J. Determination of potential migrants in polycarbonate containers used for microwave ovens by high-performance liquid chromatography with ultraviolet and fluorescence detection. J. Agric. Food Chem., 51, 5647–5653 (2003).
- 24) Russo A, Cardile V, De Ioannes A, Garbarino J. Effect of litreol on the viability of human cancer cells. Chem. Biol. Interact., 179, 178–184 (2009).
- 25) Kozubek A, Tyman JH. Resorcinolic lipids, the natural non-isoprenoid phenolic amphiphiles and their biological activity. Chem. Rev., 99, 1–26 (1999).
- 26) Yoon MA, Jeong TS, Park DS, Xu MZ, Oh HW, Song KB, Lee WS, Park HY. Antioxidant effects of quinoline alkaloids and 2,4-di-tert-butylphenol isolated from Scolopendra subspinipes. Biol. Pharm. Bull., 29, 735–739 (2006).
- 27) Choi SJ, Kim JK, Kim HK, Harris K, Kim CJ, Park GG, Park CS, Shin DH. 2,4-Di-tert-butylphenol from sweet potato protects against oxidative stress in PC12 cells and in mice. J. Med. Food, 16, 977–983 (2013).
- 28) O’Connell A, Earley B, Leonard BE. The neuroprotective effect of tacrine on trimethyltin induced memory and muscarinic receptor dysfunction in the rat. Neurochem. Int., 25, 555–566 (1994).
- 29) Brown AW, Aldridge WN, Street BW, Verschoyle RD. The behavioral and neuropathologic sequelae of intoxication by trimethyltin compounds in the rat. Am. J. Pathol., 97, 59–82 (1979).
- 30) Dyer RS. Physiological methods for assessment of trimethyltin exposure. Neurobehav. Toxicol. Teratol., 4, 659–664 (1982).
- 31) Kobayashi H, Saito F, Yuyama A. Effects of organotins on the cholinergic system in the chicken brain in vitro. Toxicol. In Vitro, 6, 337–343 (1992).