Abstract
External stimuli, such as radiation, induce inflammatory cytokine and chemokine production in skin, but the mechanisms involved are not completely understood. We previously showed that the P2Y11 nucleotide receptor, p38 mitogen-activated protein kinase (MAPK) and nuclear factor-kappa B (NF-κB) all participate in interleukin (IL)-6 production induced by γ-irradiation. Here, we focused on the transient receptor potential vanilloid 4 (TRPV4) channel, which is expressed in skin keratinocytes and has been reported to play a role in inflammation. We found that irradiation of human epidermal keratinocytes HaCaT cells with 5 Gy of γ-rays (137Cs: 0.75 Gy/min) induced IL-6 and IL-8 production. HaCaT cells treated with TRPV4 channel agonist GSK1016790A also showed increased IL-6 and IL-8 production. In both cases, IL-6/IL-8 production was not increased at 24 h after stimulation, but was increased at 48 h. ATP was released from cells exposed to γ-irradiation or TRPV4 channel agonist, and the release was suppressed by TRPV4 channel inhibitors. The γ-irradiation-induced increase in IL-6 and IL-8 production was suppressed by apyrase (ecto-nucleotidase), NF157 (selective P2Y11 receptor antagonist) and SB203580 (p38 MAPK inhibitor). GSK1016790A-induced inhibitor of kappa B-alpha (IκBα) decomposition, which causes NF-κB activation was suppressed by NF157 and SB203580, and γ-irradiation-induced IκBα decomposition was suppressed by TRPV4 channel inhibitors. Our results suggest that γ-irradiation of keratinocytes induces ATP release via activation of the TRPV4 channel, and then ATP activates P2Y11 receptor and p38 MAPK-NF-κB signaling, resulting in IL-6/IL-8 production.
It is well-known that stimuli such as ultraviolet or gamma (γ-) irradiation can cause skin inflammation due to overproduction of cytokines and chemokines.1) Indeed, γ-irradiation, which is commonly used as a treatment for cancer, can cause a range of side effects, such as depilation, skin dryness, erythema, pigmentation, bubbles, erosion, skin ulcer, necrosis and even carcinogenesis.2–5) However, the mechanism through which γ-irradiation induces production of cytokines/chemokines in epidermal cells is not fully understood.
ATP is released from human epidermal keratinocyte-derived HaCaT cells in response to various stimuli6) and induces epidermal cell proliferation, differentiation and apoptosis7) through autocrine and paracrine signal transduction via the P2 receptor, which is expressed on the cell membrane.8,9) Extracellular ATP also induces production of interleukin (IL)-6 (pro-inflammatory cytokine) and IL-8 (chemokine) in epidermal cells.10) In addition, we have reported an important role in P2Y11 receptor in lipopolysaccharide (LPS)-induced IL-6 production in human monocyte THP-1 cells.11) Further, we have also reported the involvement of P2Y11 receptor in IL-6 production by interferon-γ, silica nanoparticles, or γ-irradiation in HaCaT cells.12–14) P2Y11 receptors are also involved in the induction of IL-6 production in response to UVA irradiation in HaCaT cells,15) and activation of P2Y11 receptors induces IL-8 production in human monocyte-derived dendritic cells.16) However, little work has been done on the mechanism of γ-irradiation-induced IL-8 production or the mediators of ATP release in HaCaT cells.
We previously reported the involvement of P2Y11 receptor, p38 mitogen-activated protein kinase (MAPK), and nuclear factor-kappa B (NF-κB) in γ-irradiation-induced IL-6 production in HaCaT cells.14) γ-Irradiation induces release of ATP, which activates P2Y11 receptor, although we found that P2Y11 receptor is not activated immediately after γ-irradiation, but only at about 18 h after irradiation. Taking account of these findings, we focused here on the transient receptor potential vanilloid 4 (TRPV4) channel, which is expressed in skin keratinocytes and has been reported to play a role in inflammation, as a candidate for mediating ATP release in response to γ-irradiation.
The TRPV4 channel is a nonselective cation channel that was identified as an osmotic pressure sensor in 2000.17,18) Subsequent studies revealed not only hypotonic stimulation, but also resulted in the discovery of other irritant receptors activated by stimulants such as arachidonic acid and its metabolites,19) endocannabinoid,20) and mechanical stimulation.21) TRPV4 is also activated by thermal stimulation22,23) (27–35°C). It is expressed in various locations, including the nervous system, kidneys, skin, blood vessels, lungs and bladder,24,25) and was recently shown to be involved in production of inflammatory mediators,26) including IL-627–29) and IL-8.30) It has also been reported to be involved in ATP release.31–33) Therefore, in this work we investigated the role of TRPV4 channel in ATP release after γ-irradiation of HaCaT cells, as well as the signalling pathway leading to IL-6/IL-8 production.
MATERIALS AND METHODS
ReagentsApyrase, SB203580, and GSK2193874 were purchased from Sigma-Aldrich (U.S.A.). NF157 was purchased from Tocris Bioscience (U.K.). RN-1734 and GSK1016790A were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan).
Cell Culture and IrradiationThe culture of immortalized human-derived epidermal keratinocytes HaCaT cells and γ-irradiation were performed as described previously.14) HaCaT cells were kindly supplied by Drs. M. Ichihashi and M. Ueda (Kobe University School of Medicine, Kobe, Japan) with the permission of Dr. N. E. Fusening (German Cancer Research Center, Heidelberg, Germany).34,35) The cells were irradiated with γ-rays from a Gammacell 40 (137Cs source) (Nordin International, Inc., Japan; 0.75 Gy/min) at room temperature.
ImmunoblottingImmunoblotting was performed as described previously.14) In brief, cells were washed with phosphate-buffered saline (PBS) (Wako, Japan) twice and incubated in lysis buffer containing 1% Triton X-100, protease inhibitor cocktail, and PHOSSTOP® (Sigma-Aldrich) at 4°C for 30 min. The cell lysate was centrifuged at 10000×g for 15 min, and the supernatant was mixed with 2x Laemmli sample buffer (Wako) and 10 mM dithiothreitol. Samples were incubated at 95°C for 10 min. Samples (1.5 µg protein) were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the bands were transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were incubated overnight at 4°C in TBST (0.1% Tween-20, 10 mM Tris–HCl, 0.1 M NaCl) containing 1% bovine serum albumin (BSA) (Wako), further incubated overnight at 4°C with anti-TRPV4 antibody (Sigma-Aldrich) or mouse anti-inhibitor of kappa B-alpha (IκBα) (L35A5) monoclonal antibody (mAb) (1 : 1000) (Cell Signaling Technology, Inc., U.S.A.), washed with TBST for 30 min, incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit immunoglobulin G (IgG) antibody (1 : 20000) (Cell Signaling Technology, Inc.) or HRP-conjugated anti-mouse IgG antibody (1 : 20000) (Cell Signaling Technology, Inc.) for 1.5 h at room temperature, and then washed again with TBST for 30 min. Membranes were also incubated for 1 h at room temperature with peroxidase-conjugated anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mAb (1 : 50000) (Wako), to confirm equal loading. Specific proteins were visualized by using ImmunoStar®LD (Wako), and bands were analyzed with Image Studio 4.0 for C-DiGit Scanner (LI-COR, U.K.).
Cytokine ProductionHaCaT cells (1.0×105 cells/mL) were seeded and incubated for 24 h. After incubation, the cells were stimulated by exposure to γ-irradiation or GSK1016790A. The concentration of IL-6 in culture medium was measured by enzyme-linked immunosorbent assay (ELISA) as described previously.14) The concentration of IL-8 was measured with human IL-8 ELISA Ready-SET-Go!®(2nd Generation) (eBioscience, U.S.A.) according to the manufacturer’s instructions.
Measurement of Extracellular ATPThe concentration of ATP in culture medium was measured as described previously.14) In brief, HaCaT cells (1.5×105 cells/mL) were seeded and incubated for 24 h. The culture medium was changed to phenol red-free medium containing 0.5% FBS for 1 h before irradiation. The culture medium was harvested at 24 h after irradiation. The medium was centrifuged at 600×g for 1 min at 4°C. The supernatant (10 µL) was added to wells of a white 96-well plate, and then 100 µL of Luciferase/Luciferin Reagent (Promega) was injected into each well. The chemiluminescence was measured with a WALLAC ARVO SX multilabel counter (PerkinElmer, Inc., U.S.A.).
StatisticsResults were expressed as mean±standard error (S.E.). The statistical significance of differences between control and other groups was calculated using Dunnett’s test. Multiple groups were compared using ANOVA followed by pairwise comparisons with Bonferroni’s post hoc analysis. Calculations were done with the Instat version 3.0 statistical software package (Graph Pad Software, U.S.A.). The criterion of significance was p<0.05.
RESULTS AND DISCUSSION
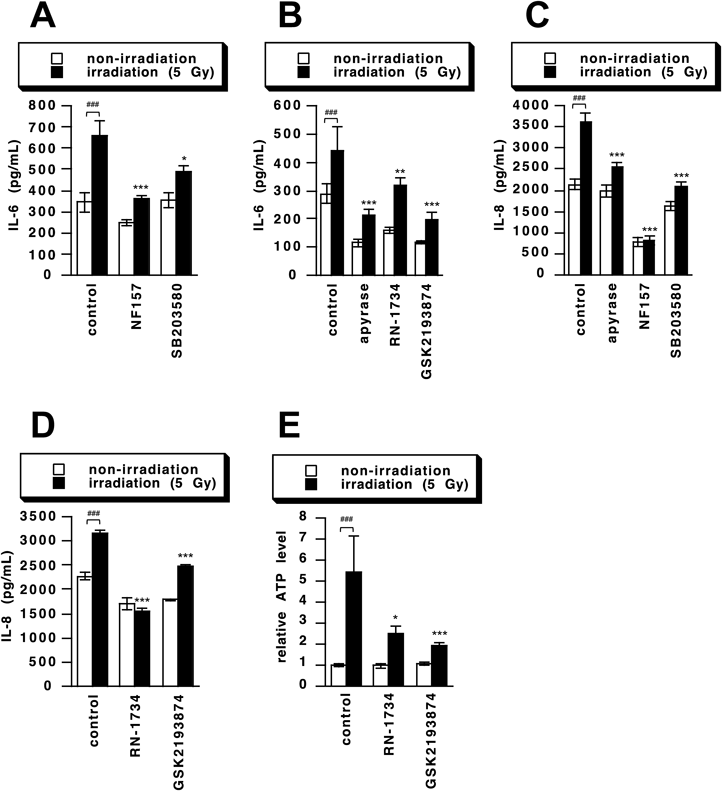
First, expression of TRPV4 channel in HaCaT cells was confirmed (Fig. 1A). Then, HaCaT cells were irradiated with 5 Gy of γ-rays or stimulated with TRPV4 channel agonist GSK1016790A, and we measured IL-6 and IL-8 production. Though IL-6 and IL-8 production was unchanged at 24 h after stimulation, it was significantly increased at 48 h (Figs. 1B, C). This is in agreement with the results of our previous study on IL-6 production,14) in which we demonstrated the involvement of ATP-mediated P2Y11 receptor activation and p38 MAPK signaling in γ-irradiation-induced IL-6 production. Since IL-8 production induced by γ-irradiation showed similar time dependence, we speculated that the similar pathways were involved. This was confirmed by the findings that the γ-irradiation-induced increase in IL-6/IL-8 production was suppressed by apyrase (ecto-nucleotidase), NF157 (a selective P2Y11 receptor antagonist) and SB203580 (a p38 MAPK inhibitor) (Figs. 2A–C).
We have reported that ATP release from γ-irradiated cells,14) and it was recently shown that TRPV4 channel stimulation causes ATP release in esophageal keratinocytes, gastric epithelia and odontoblasts.31–33) Therefore, we next investigated the role of TRPV4 in γ-irradiation-induced IL-6 and IL-8 production by using two TRPV4 channel inhibitors, RN-1734 and GSK2193874. Both inhibitors suppressed the γ-irradiation-induced increase in production of IL-6 and IL-8 (Figs. 2B, D). Further, the amount of ATP released from HaCaT cells was increased at 24 h after γ-irradiation, and the γ-irradiation-induced ATP release was suppressed by RN-1734 and GSK2193874 (Fig. 2E). As reported in previous paper,14) the concentration of ATP in culture medium was detected at pM order, because the released ATP was metabolized by ecto-nucleotidase and diluted in culture medium though pericellular ATP concentration would be much higher. To determine the involvement of TRPV4 channel in the ATP release, we here showed ATP release as a ratio of control. These results indicate that the TRPV4 channel is involved in γ-irradiation-induced IL-6 and IL-8 production and ATP release. Thus, we next investigated whether stimulation of the TRPV4 channel would induce production of IL-6 and IL-8. Indeed, the amount of ATP released from HaCaT cells were increased at 24 h after stimulation with a TRPV4 agonist, GSK1016790A. The GSK1016790A-induced ATP release was suppressed by RN-1734 and GSK2193874 (Fig. 3A). We also investigated the signaling pathways, and found that the GSK1016790A-induced IL-6 and IL-8 production was suppressed by apyrase, NF157, SB203580, RN-1734 and GSK2193874 (Figs. 3B–E). These results strongly suggest that the TRPV4 channel is involved in IL-6 and IL-8 production by regulating γ-irradiation-induced ATP release.
It has been reported that NF-κB is involved in γ-irradiation-induced IL-6 production in various cells,36–39) including HaCaT cells.14) Therefore, we investigated whether TRPV4 channel is involved in the activation of NF-κB in irradiated cells. Activation of NF-κB occurs via decomposition of IκBα, which is an inhibitor of NF-κB. Therefore, we evaluated the activation of NF-κB by measuring relative IκBα level. We found that GSK1016790A promotes the decomposition of IκBα, supporting the idea that activation of the TRPV4 channel leads to activation of NF-κB. GSK1016790A-induced IκBα decomposition was suppressed by NF157 and SB203580 (Figs. 4A, B). In addition, IκBα was decreased by γ-irradiation, and the decrease was suppressed by RN-1734 and GSK2193874 (Figs. 4C, D). These results suggest that TRPV4 channel activation induces P2Y11 receptor activation and p38 MAPK-mediated NF-κB activation. Since we have previously shown that the IL-6 production is mediated through P2Y11 receptor-p38 MAPK-NF-κB pathway in γ-irradiatted HaCaT cells,14) activation of the TRPV4 channel appears to act as a trigger of this pathway by regulating ATP release.
ATP release pathways include maxi anion channel, pannexin hemichannel, connexin hemichannel, exocytosis concentrated into vesicle by vesicle nucleotide transporter (VNUT), and other anion channel(s).40) Among them, involvement of anion channels or hemichannels in radiation-induced ATP release has been reported. γ-Irradiation-induced ATP release has also been reported to involve maxi anion channels in HaCaT cells.41) On the other hand, hemichannels and VNUT are involved in TRPV4 channel-induced ATP release,31,42) and there is one report describing the involvement of TRPV4 as a regulator of intracellular Ca2+ in human esophageal epithelial cells.30) Here, we provide the first evidence that TRPV4 channel-regulated ATP release is involved in γ-irradiation-induced IL-6 and IL-8 production in keratinocytes. However, further work will be needed to determine the mechanism of γ-irradiation-induced TRPV4 activation and the pathway of ATP release.
Since cytokines and chemokines promote cancer and metastasis,43,44) our present findings could be helpful in developing ways to reduce these undesired side effects of radiotherapy. In addition, the TRPV4 channel might be a novel therapeutic target for γ-irradiation-induced inflammation.
Acknowledgment
Parts of this work were supported by Grants-in-Aid for Pharmaceutical Scientific Research (to MT) from Takeda Science Foundation.
Conflict of Interest
The authors declare no conflict of interest.
REFERENCES
- 1) Zhao W, Robbins MEC. Inflammation and chronic oxidative stress in radiation-induced late normal tissue injury: therapeutic implications. Curr. Med. Chem., 16, 130–143 (2009).
- 2) Coussens LM, Werb Z. Inflammation and cancer. Nature, 420, 860–867 (2002).
- 3) Ancrile B, Lim K-H, Counter CM. Oncogenic Ras-induced secretion of IL6 is required for tumorigenesis. Genes Dev., 21, 1714–1719 (2007).
- 4) Kuilman T, Michaloglou C, Vredeveld LCW, Douma S, van Doorn R, Desmet CJ, Aarden LA, Mooi WJ, Peeper DS. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell, 133, 1019–1031 (2008).
- 5) Yang G-Y, Taboada S, Liao J. Inflammatory Bowel Disease: A model of chronic inflammation-induced cancer. Methods Mol. Biol., 511, 193–233 (2009).
- 6) Bakondi E, Gönczi M, Szabó É, Bai P, Pacher P, Gergely P, Kovács L, Hunyadi J, Szabó C, Csernoch L, Virág L. Role of intracellular calcium mobilization and cell-density-dependent signaling in oxidative-stress-induced cytotoxicity in HaCaT keratinocytes. J. Invest. Dermatol., 121, 88–95 (2003).
- 7) Bowman PD, Schuschereba ST, Lawlor DF, Gilligan GR, Mata JR, DeBaere DR. Survival of human epidermal keratinocytes after short-duration high temperature: synthesis of HSP70 and IL-8. Am. J. Physiol., 272, C1988–C1994 (1997).
- 8) Burnstock G. Introduction: P2 receptors. Curr. Top. Med. Chem., 4, 793–803 (2004).
- 9) Khakh BS, North RA. P2X receptors as cell-surface ATP sensors in health and disease. Nature, 442, 527–532 (2006).
- 10) Kawakami M, Kaneko N, Anada H, Terai C, Okada Y. Measurement of interleukin-6, interleukin-10, and tumor necrosis factor-alpha levels in tissues and plasma after thermal injury in mice. Surgery, 121, 440–448 (1997).
- 11) Sakaki H, Tsukimoto M, Harada H, Moriyama Y, Kojima S. Autocrine regulation of macrophage activation via exocytosis of ATP and activation of P2Y11 receptor. PLOS ONE, 8, e59778 (2013).
- 12) Ishimaru M, Tsukimoto M, Harada H, Kojima S. Involvement of P2Y11 receptor in IFN-γ-induced IL-6 production in human keratinocytes. Eur. J. Pharmacol., 703, 67–73 (2013).
- 13) Nagakura C, Negishi Y, Tsukimoto M, Itou S, Kondo T, Takeda K, Kojima S. Involvement of P2Y11 receptor in silica nanoparticles 30-induced IL-6 production by human keratinocytes. Toxicology, 322, 61–68 (2014).
- 14) Ohsaki A, Miyano Y, Tanaka R, Tanuma S, Kojima S, Tsukimoto M. A novel mechanism of γ-irradiation-induced IL-6 production mediated by P2Y11 receptor in epidermal keratinocytes. Biol. Pharm. Bull., 41, 925–936 (2018).
- 15) Kawano A, Kadomatsu R, Ono M, Kojima S, Tsukimoto M, Sakamoto H. Autocrine regulation of UVA-induced IL-6 production via release of ATP and activation of P2Y receptors. PLOS ONE, 10, e0127919 (2015).
- 16) Meis S, Hamacher A, Hongwiset D, Marzian C, Wiese M, Eckstein N, Royer H-D, Communi D, Boeynaems J-M, Hausmann R, Schmalzing G, Kassack MU. NF546 [4,4′-(carbonylbis(imino-3,1-phenylene-carbonylimino-3,1-(4-methyl-phenylene)-carbonylimino))-bis(1,3-xylene-alpha,alpha′-diphosphonic acid) tetrasodium salt] is a non-nucleotide P2Y11 agonist and stimulates release of interleukin-8 from human monocyte-derived dendritic cells. J. Pharmacol. Exp. Ther., 332, 238–247 (2010).
- 17) Strotmann R, Harteneck C, Nunnenmacher K, Schultz G, Plant TD. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol., 2, 695–702 (2000).
- 18) Liedtke W, Choe Y, Martí-Renom MA, Bell AM, Denis CS, Sali A, Hudspeth AJ, Friedman JM, Heller S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell, 103, 525–535 (2000).
- 19) Nilius B, Vriens J, Prenen J, Droogmans G, Voets T. TRPV4 calcium entry channel: a paradigm for gating diversity. Am. J. Physiol. Cell Physiol., 286, C195–C205 (2004).
- 20) Watanabe H, Vriens J, Prenen J, Droogmans G, Voets T, Nilius B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature, 424, 434–438 (2003).
- 21) Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Transient receptor potential vanilloid 4 is essential in chemotherapy-induced neuropathic pain in the rat. J. Neurosci., 24, 4444–4452 (2004).
- 22) Güler AD, Lee H, Iida T, Shimizu I, Tominaga M, Caterina M. Heat-evoked activation of the ion channel, TRPV4. J. Neurosci., 22, 6408–6414 (2002).
- 23) Watanabe H, Vriens J, Suh SH, Benham CD, Droogmans G, Nilius B. Heat-evoked activation of TRPV4 channels in a HEK293 cell expression system and in native mouse aorta endothelial cells. J. Biol. Chem., 277, 47044–47051 (2002).
- 24) Shibasaki K, Suzuki M, Mizuno A, Tominaga M. Effects of body temperature on neural activity in the hippocampus: regulation of resting membrane potentials by transient receptor potential vanilloid 4. J. Neurosci., 27, 1566–1575 (2007).
- 25) Sokabe T, Fukumi-Tominaga T, Yonemura S, Mizuno A, Tominaga M. The TRPV4 channel contributes to intercellular junction formation in keratinocytes. J. Biol. Chem., 285, 18749–18758 (2010).
- 26) Ye L, Kleiner S, Wu J, Sah R, Gupta RK, Banks AS, Cohen P, Khandekar MJ, Boström P, Mepani RJ, Laznik D, Kamenecka TM, Song X, Liedtke W, Mootha VK, Puigserver P, Griffin PR, Clapham DE, Spiegelman BM. TRPV4 is a regulator of adipose oxidative metabolism, inflammation, and energy homeostasis. Cell, 151, 96–110 (2012).
- 27) Nayak PS, Wang Y, Najrana T, Priolo LM, Rios M, Shaw SK, Sanchez-Esteban J. Mechanotransduction via TRPV4 regulates inflammation and differentiation in fetal mouse distal lung epithelial cells. Respir. Res., 16, 60 (2015).
- 28) Walter BA, Purmessur D, Moon A, Occhiogrosso J, Laudier DM, Hecht AC, Iatridis JC. Reduced tissue osmolarity increases TRPV4 expression and pro-inflammatory cytokines in intervertebral disc cells. Eur. Cell. Mater., 32, 123–136 (2016).
- 29) Dalsgaard T, Sonkusare SK, Teuscher C, Poynter ME, Nelson MT. Pharmacological inhibitors of TRPV4 channels reduce cytokine production, restore endothelial function and increase survival in septic mice. Sci. Rep., 6, 33841 (2016).
- 30) Ueda T, Shikano M, Kamiya T, Joh T, Ugawa S. The TRPV4 channel is a novel regulator of intracellular Ca2+ in human esophageal epithelial cells. Am. J. Physiol. Liver Physiol., 301, G138–G147 (2011).
- 31) Mihara H, Boudaka A, Sugiyama T, Moriyama Y, Tominaga M. Transient receptor potential vanilloid 4 (TRPV4)-dependent calcium influx and ATP release in mouse oesophageal keratinocytes. J. Physiol., 589, 3471–3482 (2011).
- 32) Mihara H, Suzuki N, Boudaka AA, Muhammad JS, Tominaga M, Tabuchi Y, Sugiyama T. Transient receptor potential vanilloid 4-dependent calcium influx and ATP release in mouse and rat gastric epithelia. World J. Gastroenterol., 22, 5512–5519 (2016).
- 33) Egbuniwe O, Grover S, Duggal AK, Mavroudis A, Yazdi M, Renton T, Di Silvio L, Grant AD. TRPA1 and TRPV4 activation in human odontoblasts stimulates ATP release. J. Dent. Res., 93, 911–917 (2014).
- 34) Boukamp P, Petrussevska RT, Breitkreutz D, Hornung J, Markham A, Fusenig NE. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol., 106, 761–771 (1988).
- 35) Takasawa R, Nakamura H, Mori T, Tanuma S. Differential apoptotic pathways in human keratinocyte HaCaT cells exposed to UVB and UVC. Apoptosis, 10, 1121–1130 (2005).
- 36) Brennenstuhl H, Armento A, Braczysnki AK, Mittelbronn M, Naumann U. IκBζ, an atypical member of the inhibitor of nuclear factor kappa B family, is induced by γ-irradiation in glioma cells, regulating cytokine secretion and associated with poor prognosis. Int. J. Oncol., 47, 1971–1980 (2015).
- 37) Mangoni M, Sottili M, Gerini C, Desideri I, Bastida C, Pallotta S, Castiglione F, Bonomo P, Meattini I, Greto D, Olmetto E, Terziani F, Becherini C, Delli Paoli C, Trombetta L, Loi M, Biti G, Livi L. A PPAR gamma agonist protects against oral mucositis induced by irradiation in a murine model. Oral Oncol., 64, 52–58 (2017).
- 38) Zaher NH, Salem AAM, Ismail AFM. Novel amino acid derivatives bearing thieno[2,3-d]pyrimidine moiety down regulate NF-κB in γ-irradiation mediated rat liver injury. J. Photochem. Photobiol. B, 165, 328–339 (2016).
- 39) Linard C, Marquette C, Mathieu J, Pennequin A, Clarençon D, Mathé D. Acute induction of inflammatory cytokine expression after gamma-irradiation in the rat: effect of an NF-kappaB inhibitor. Int. J. Radiat. Oncol. Biol. Phys., 58, 427–434 (2004).
- 40) Tsukimoto M. Purinergic signaling is a novel mechanism of the cellular response to ionizing radiation. Biol. Pharm. Bull., 38, 951–959 (2015).
- 41) Tsukimoto M, Homma T, Ohshima Y, Kojima S. Involvement of purinergic signaling in cellular response to γ radiation. Radiat. Res., 173, 298–309 (2010).
- 42) Shahidullah M, Mandal A, Delamere NA. TRPV4 in porcine lens epithelium regulates hemichannel-mediated ATP release and Na-K-ATPase activity. Am. J. Physiol. Physiol., 302, C1751–C1761 (2012).
- 43) Liu Q, Yu S, Li A, Xu H, Han X, Wu K. Targeting interlukin-6 to relieve immunosuppression in tumor microenvironment. Tumour Biol., 39, 1010428317712445 (2017).
- 44) Dominguez C, David JM, Palena C. Epithelial–mesenchymal transition and inflammation at the site of the primary tumor. Semin. Cancer Biol., 47, 177–184 (2017).