Abstract
Anticancer drugs exhibit many side effects, including skin pigmentation, which often lowers patient QOL. However, the mechanism of pigmentation caused by anticancer drugs remains unknown. The purpose of this study was to elucidate the mechanism of anticancer drug-induced skin pigmentation using 5-fluorouracil (5-FU), a widely used anticancer drug. Specific pathogen-free, 9-week-old Hos:HRM-2 male mice were intraperitoneally administered 5-FU daily for 8 weeks. Skin pigmentation was observed at the end of the study. Mice treated with 5-FU were also administered inhibitors of cAMP, α-melanocyte-stimulating hormone (α-MSH), and adrenocorticotropic hormone (ACTH) for analysis. Administration of oxidative stress, nuclear factor-kappa B (NF-κB), cAMP, and ACTH inhibitors reduced pigmentation in 5-FU-treated mice. These results indicate that the oxidative stress/NF-κB/ACTH/cAMP/tyrosinase pathway plays an important role in pigmentation in 5-FU-treated mice.
INTRODUCTION
The skin, which acts as a barrier to the outside world, is constantly exposed to environmental stressors, such as UV radiation and dryness. Facial skin is particularly prone to the formation of spots, wrinkles, and sagging when exposed to sunlight. Dark spots are pigmentation abnormalities, where melanin pigment deposits on the epidermis. Melanin is produced by melanocytes present in 1 out of 10 cells in the basal layer of the epidermis. Various mechanisms underlying the synthesis of excess melanin due to environmental stress have been reported. Reactive oxygen species (ROS) are generated when the skin is exposed to stressful stimuli. ROS promote the production of proteins, such as endothelin-1, stem cell growth factor, α-melanocyte-stimulating hormone (α-MSH), cyclooxygenase, interleukin (IL)-1α, and macrophage migration inhibitory factor. Among these, the most widely studied is α-MSH, which binds to melanocortin 1 receptor (MC1R) on the membrane, leading to an increase in cAMP and the subsequent activation of protein kinase A/cAMP response element binding protein/microphthalmia-associated transcription factor signaling, thereby increasing tyrosinase enzyme activity.1–4) Melanin synthesis is initiated by the catalytic reaction of tyrosinase with tyrosine as the substrate. Tyrosine is converted to dopaquinone/dopachrome/5,6-dihydoroxyindole-2-carboxylic acid, eventually forming melanin, which is transported to keratinocytes.5) Thus, abnormalities involving cutaneous pigmentation induced by cutaneous irritants, such as UV, are well documented. However, UV is not the only factor that causes skin abnormalities, and skin is also affected by allergies and internal diseases.6,7)
Anticancer drugs are a factor that affects the skin. Skin damage induced by anticancer drugs has a minimal direct impact, but psychological distress due to changes in appearance reduces patient QOL and complicates compliance with anticancer treatment.8) Skin disorders caused by anticancer agents can occur in various forms, including acneiform eruptions, dry skin, and paronychia. Many preventive and therapeutic methods have been studied for skin disorders to improve treatment with anticancer drugs.8–11) However, although there are some case reports on pigmentation, which is among these skin disorders,12,13) the underlying causes have not been investigated, and treatment methods have not been established.
The future development of anticancer drugs should investigate the cause of pigmentation and take countermeasures to facilitate treatment with effective anticancer drugs. The purpose of this study was to investigate the mechanism of pigmentation as a side effect of anticancer drugs using 5-fluorouracil (5-FU), a widely used anticancer agent.
MATERIALS AND METHODS
Animal ExperimentsSpecific pathogen-free (SPF), 9-week-old Hos:HRM-2 male mice (colored hairless mouse; Japan SLC, Inc., Hamamatsu, Shizuoka, Japan) were used for the experiments. These mice have melanocytes in the auricle, eyes, toes, tail, and skin around the buttocks, and they can synthesize melanin. All mice were individually housed in cages at a controlled temperature of 23 ± 1 °C under SPF conditions with a 12-h light/dark cycle. All animals were allowed free access to laboratory chow (rodent diet EQ 5L37; Japan SLC, Inc.) and water during the experiments. We used five animals per group and repeated the experiments three times. This study strictly followed the recommendations and guidelines for the Care and Use of Laboratory Animals of Suzuka University of Medical Science (Approval No. 34/October 7, 2017). All surgical procedures were performed under pentobarbital anesthesia, and every effect was adjusted to minimize animal suffering.
5-FU TreatmentApproximately 15 mg/kg of 5-FU (Kyowa Kirin Co., Ltd., Tokyo, Japan) in saline was intraperitoneally injected to mice once daily for 8 weeks. The solvent-injected animals were administered saline. Based on the method described by Huang et al.,14) concentrations were set to 5, 15, and 30 mg/kg, and the lowest effective concentration given to mice was adopted.
MC1R Antagonist [Agouti Signaling Protein (ASIP)] TreatmentApproximately 10 nM of ASIP (R&D Systems, Minneapolis, MN, U.S.A.) in 1% dimethyl sulfoxide (DMSO) was intraperitoneally injected to mice three times per week for 8 weeks. The solvent-injected animals were administered 1% DMSO.15)
cAMP Antagonist {α-[2-(3-Chlorophenyl) hydrazinylidene]-5-(1,1-dimethylethyl)-β-oxo-3-isoxazolepropanenitrile (ESI-09)} TreatmentApproximately 20 mg/kg of ESI-09 (Cayman, Ann Arbor, MI, U.S.A.) in a solution of DMSO: phosphate-buffered saline (PBS; ratio 1 : 7) was intraperitoneally injected to mice three times per week for 8 weeks. The solvent-injected animals were administered a solution of DMSO : PBS (ratio 1 : 7).16)
Adrenocorticotropic Hormone (ACTH) Receptor Antagonist [ACTH (11-24)] TreatmentApproximately 20 µg/kg of ACTH (11-24) (MedChemExpress, Monmouth Junction, NJ, U.S.A.) in saline was intraperitoneally injected to mice three times per week for 8 weeks. The solvent-injected animals were administered saline.17)
Nuclear Factor-kappa B (NF-κB) Inhibitor [4-Methyl-N1-(3-phenyl-propyl)-benzene-1,2-diamine (JSH-23)] TreatmentApproximately 1 mg/kg of JSH-23 (MedChemExpress) in 0.5% sodium carboxymethyl cellulose (CMC) was orally administered to mice once daily for 8 weeks. The solvent-injected animals were administered 0.5% CMC.18)
ROS Inhibitor (N-Acetyl-L-cysteine (NAC)) TreatmentNAC (200 mg/kg; Nacalai Tesque, Kyoto, Japan) in 0.08% DMSO was intraperitoneally injected to mice once daily for 8 weeks. The solvent-injected animals were administered 0.08% DMSO.19)
Preparation and Staining of the Skin around the ButtocksWe obtained samples from the skin around the buttocks after 8 weeks of starting the experiment. Specimens were fixed in phosphate-buffered paraformaldehyde (4%), embedded in frozen Tissue-Tek, an optimal cutting temperature compound, and cut into 5-µm sections. The sections were subjected to immunostaining; the staining method is described in details in our previous study.20) Briefly, the specimens were incubated with rabbit polyclonal anti-tyrosinase (1 : 100; Cell Signaling Technology Inc., Danvers, MA, U.S.A.) primary antibodies. The samples were then washed and incubated with fluorescein isothiocyanate-conjugated anti-rabbit (1 : 30; Dako Cytomation, Glstrup, Denmark) secondary antibodies. The expression levels were evaluated immunohistochemically using a fluorescence microscope. Other sections were stained using 3,4-dihydroxyphenylalanine (DOPA). DOPA-positive melanocytes in the skin around the buttocks were stained as described in a previous study.21) The skin around the buttocks was washed with PBS and incubated in PBS containing 0.1% L-DOPA at 37 °C (Sigma-Aldrich Chemical Co., St. Louis, MO, U.S.A.).
Measurement of ACTH, α-MSH, and cAMP Levels in the Skin around the Buttocks of 5-FU-Treated MiceSamples from the skin around the buttocks were collected the last day of the experiment, as described in a previous study.22) Briefly, 100 mg of skin was rinsed with PBS and homogenized, and the supernatant was collected for the assay. The ACTH, α-MSH, and cAMP levels in the skin samples were determined using commercial enzyme-linked immunosorbent assay (ELISA) kits (ACTH and α-MSH: Abcam, Cambridge, MA, U.S.A.; cAMP; MyBioSource, San Diego, CA, U.S.A.) according to the manufacturers’ instructions. Optical density was measured using a microplate reader (Molecular Devices, Sunnyvale, CA, U.S.A.).
Statistical AnalysisAll data are presented as means ± standard deviation (S.D.). Microsoft Excel 2010 (Microsoft Corp., Redmond, WA, U.S.A.) was used to analyze the statistical significance of the data, with one-way ANOVA followed by Tukey’s post-hoc test using SPSS software version 20 (SPSS Inc., Chicago, IL, U.S.A.). Results with p values *< 0.05 or **< 0.01 were considered statistically significant.
RESULTS
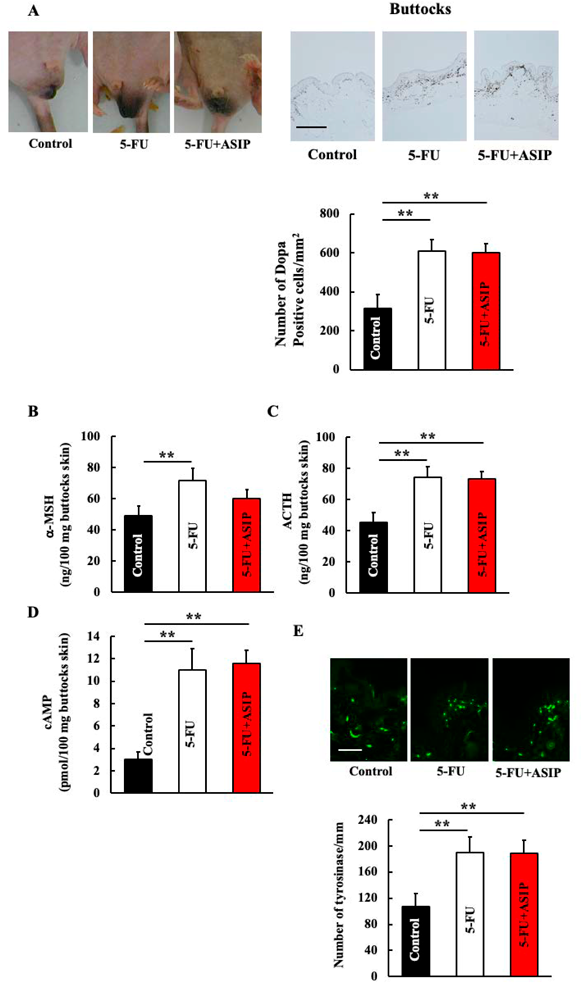
Effect of 5-FU Treatment on Pigmentation and Its Associated Genes in the Skin around the ButtocksEight weeks of 5-FU administration increased DOPA-positive melanocytes in the skin around the buttocks of mice (Fig. 1A). Furthermore, increased ACTH, α-MSH, and cAMP levels and tyrosinase expression were observed following 5-FU treatment (Figs. 1B–E).
Effect of ASIP Administration on Pigmentation and Its Associated Genes in the Skin around the Buttocks of 5-FU-Treated MiceWe administered ASIP, an α-MSH inhibitor, to examine the effects of α-MSH. No difference in pigmentation was observed between 5-FU-treated and ASIP + 5-FU-treated mice (Fig. 2A). In addition, α-MSH, ACTH, and cAMP levels and tyrosinase expression in the skin around the buttocks did not differ between the two groups of mice (Figs. 2B–E).
Effect of ESI-09 Administration on Pigmentation and Its Associated Genes in the Skin around the Buttocks of 5-FU-Treated MiceESI-09, a cAMP inhibitor, was administered to evaluate the effects of cAMP. Pigmentation was reduced in ESI-09 + 5-FU-treated mice compared to 5-FU-treated mice (Fig. 3A). The cAMP level and tyrosinase expression also decreased in ESI-09 + 5-FU-treated mice, compared to 5-FU-treated mice (Figs. 3D, E). However, the levels of ACTH and α-MSH in the skin around the buttocks did not differ (Figs. 3B, C).
Effect of ACTH (11-24) Administration on Pigmentation and Its Associated Genes in the Skin around the Buttocks of 5-FU-Treated MiceWe administered ACTH (11-24), an ACTH inhibitor, to study the effects of ACTH. Pigmentation was reduced in ACTH (11-24) + 5-FU-treated mice compared to 5-FU-treated mice (Fig. 4A). Similarly, cAMP levels and tyrosinase expression also decreased (Figs. 4D, E). However, the amount of α-MSH and ACTH in the skin around the buttocks did not differ between the two groups of mice (Figs. 4B, C).
Effect of JSH-23 Administration on Pigmentation and Its Associated Genes in the Skin around the Buttocks of 5-FU-Treated MiceTo elucidate the mechanism underlying the increase in ACTH levels, we used JSH-23 to inhibit the activity of NF-κB, a transcription factor that increases the ACTH levels. Pigmentation was reduced in JSH-23 + 5-FU-treated mice compared with that in 5-FU-treated mice (Fig. 5A). Concurrently, the ACTH and cAMP levels also decreased in the JSH-23 + 5-FU-treated mice compared with those in 5-FU-treated mice (Figs. 5B, C).
Effect of NAC Administration on Pigmentation and Its Associated Genes in the Skin around the Buttocks of 5-FU-Treated MiceFinally, we used NAC to inhibit oxidative stress, which is considered to be the most upstream factor of 5-FU-induced pigmentation. Pigmentation was reduced in NAC + 5-FU-treated mice, compared with that in 5-FU-treated mice (Fig. 6A). ACTH and cAMP levels also decreased in NAC + 5-FU-treated mice compared with those 5-FU-treated mice, and their levels did not differ from those in the control group (Figs. 6B, C).
DISCUSSION
In this study, mice treated with 5-FU showed an increase in skin pigmentation around the buttocks. In addition, increases in ACTH and cAMP levels and tyrosinase expression were observed in pigmented skin samples.
Pigmentation generally occurs through the production of melanin in melanocytes. Melanin synthesis occurs in melanosomes, a group of intracellular organelles in melanocytes, where tyrosine is converted by tyrosinase to catalyze the reaction.5) α-MSH is known to regulate the activity of melanocytes.22) α-MSH is a POMC-derived hormone that is secreted by the pituitary gland, as well as by keratinocytes in the skin. Melanocortin receptors are expressed in melanocytes and are involved in melanin biosynthesis. The 5-FU formulations (5-FU, capecitabine, and tegafur) used in this experiment have caused many cases of pigmentation.23) Therefore, we first administered ASIP to examine the role of MSH in melanocyte activation. ASIP is an α-MSH inhibitor that acts as an inverse agonist on specific melanocortin receptors (MCR) related to α-MSH.24) The pigmentation effect of 5-FU was not reversed, and α-MSH was not involved in 5-FU-induced pigmentation. Although ASIP has been reported to decrease cAMP levels,24) this was not observed in this study.
Next, ESI-09, a cAMP inhibitor, was administered to examine whether cAMP affects 5-FU-induced pigmentation. ESP-09 targets the cAMP-binding domain of exchange protein directly activated by cAMP (EPAC)/cAMP-regulated guanine nucleotide exchange factor (GEF) and inhibits cAMP-dependent EPAC/GEF activity.25) Administration of ESI-09 decreased the pigmentation induced by 5-FU, indicating the involvement of cAMP activation in this process.
Thus, we continued to investigate substances that activate cAMP. Addison’s disease, which is caused by adrenal insufficiency, increases ACTH production and causes skin pigmentation.26) ACTH is a stress hormone that is excised from POMC and secreted in response to stress. Herein, 5-FU administration increased ACTH production, suggesting its involvement. In addition, the administration of ACTH (11-24), an inhibitor ACTH inhibitor,17) decreased pigmentation and cAMP levels, suggesting that 5-FU causes an increase in pigmentation via ACTH. Oxidative stress is involved in the occurrence of significant 5-FU side effects, such as gastrointestinal toxicity and myelotoxicity.27) Furthermore, intraperitoneal administration of 5-FU increases oxidative stress in local tissues, which activates NF-κB to promote inflammation.28) Inflammation is known to increase the release of ACTH.29,30) Hence, these findings suggest that the administration of 5-FU may increase ACTH secretion by inducing intracellular inflammation.
Based on the above, we investigated the mechanism of the increase in ACTH levels. In this study, 5-FU administration increased blood levels of IL-6 and inducible nitric oxide synthase, which are inflammatory markers (data not shown). This inflammation activates the transcription factor NF-κB and increases the ACTH levels.29,30) Therefore, administering an NF-κB inhibitor suppressed the increase in ACTH levels and alleviated the increase in pigmentation (Fig. 5). Furthermore, 5-FU administration is known to increase oxidative stress,28) which in turn, increases inflammatory markers.28) In this study, inhibiting oxidative stress abrogated the increases in ACTH and cAMP levels (Fig. 6). These results indicate that administration of 5-FU causes an increase in pigmentation, starting with the induction of oxidative stress.
In this study, 5-FU administration induced an increase in pigmentation, and the mechanism involved the activation of the 5-FU/oxidative stress/NF-κB/ACTH/cAMP/tyrosinase pathway. However, inhibition of oxidative stress abolished both ACTH and cAMP, but could not completely suppress the increase in pigmentation. Thus, other mechanisms are also conceivable, which should be further investigated in the future.
Acknowledgments
This study was supported by JSPS KAKENHI (Grant No. 18K06082).
Conflict of Interest
The authors declare no conflict of interest.
REFERENCES
- 1) Gordon PR, Mansur CP, Gilchrest BA. Regulation of human melanocyte growth, dendricity, and melanization by keratinocyte derived factors. J. Invest. Dermatol., 92, 565–572 (1989).
- 2) Cichorek M, Wachulska M, Stasiewicz A, Tymińska A. Skin melanocytes: biology and development. Postepy Dermatol. Alergol., 30, 30–41 (2013).
- 3) Imokawa G, Yada Y, Miyagishi M. Endothelins secreted from human keratinocytes are intrindic mitogens for human melanocytes. J. Biol. Chem., 267, 24675–24680 (1992).
- 4) Hachiya A, Kobayashi A, Yoshida Y, Kitahara T, Takema Y, Imokawa G. Biphasic expression of two paracrine melanogenic cytokines, stem cell factor and endothelin-1, in ultraviolet B-induced human melanogenesis. Am. J. Pathol., 165, 2099–2109 (2004).
- 5) d’Ischia M, Wakamatsu K, Napolitano A, Briganti S, Garcia-Borron JC, Kovacs D, Meredith P, Pezzella A, Picardo M, Sarna T, Simon JD, Ito S. Melanins and melanogenesis: methods, standards, protocols. Pigment Cell Melanoma Res., 26, 616–633 (2013).
- 6) Manabe T, Inagaki Y, Nakagawa S, Miyoshi K, Ueki H. Ripple pigmentation of the neck in atopic dermatitis. Am. J. Dermatopathol., 9, 301–307 (1987).
- 7) Johnston GA, Graham-Brown RAC. The skin and disorders of the alimentary tract, hepatobiliary system, kidney, and cardiopulmonary system. Fitzpatrick’s Dermatology in General Medicine. (7th ed.) McGraw-Hill, New York, p. 1445 (2008).
- 8) Sato A, Hamada K, Imataka H. Skin toxicity. Gan To Kagaku Ryoho, 38, 1767–1772 (2011).
- 9) Lynch TJ Jr, Kim ES, Eaby B, Garey J, West DP, Lacouture ME. Epidermal growth factor receptor inhibitor-associated cutaneous toxicities: an evolving paradigm in clinical management. Oncologist, 12, 610–621 (2007).
- 10) Lacouture ME, Mitchell EP, Piperdi B, Pillai MV, Shearer H, Iannotti N, Xu F, Yassine M. Skin toxicity evaluation protocol with panitumumab (STEPP), a phase II, open-label, randomized trial evaluation the impact of a pre-emptive skin treatment regimen on skin toxicities and quality of life in patients with metastatic colorectal cancer. J. Clin. Oncol., 28, 1351–1357 (2010).
- 11) Jatoi A, Rowland K, Sloan JA, Gross HM, Fishkin PA, Kahanic SP, Novotny PJ, Schaefer PL, Johnson DB, Tschetter LK, Loprinzi CL. Tetracycline to prevent epidermal growth factor receptor inhibitor-induced skin rashes: results of a placebo-controlled trial from the North Central Cancer Treatment Group (NO3CB). Cancer, 113, 847–853 (2008).
- 12) Ohtsu A. Japanese nationwide post-marketing survey of S-1 in patients with advanced gastric cancer. Gan To Kagaku Ryoho, 33 (Suppl. 1), 52–56 (2006).
- 13) Seki T, Higaki S, Morohashi M. Drug eruption by Tegafur. Rinsho Hifuka, 43, 433–438 (1989).
- 14) Huang R, Ai G, Zhong L, Mai L, Chen JN, Liu Y, Li Y, Huang X, Su Z, Zhan JYX. Protective effects of oxyberberine in 5-fluorouracil-induced intestinal mucositis in the mice model. Evid. Based Complement. Alternat. Med., 2022, 1238358 (2022).
- 15) Hida T, Wakamatsu K, Sviderskaya EV, Donkin AJ, Montoliu L, Lynn Lamoreux ML, Yu B, Millhauser GL, Ito S, Barsh GS, Jimbow K, Bennett DC. Agouti protein, mahogunin, and attractin in pheomelanogenesis and melanoblast-like alteration of melanocytes: a cAMP-independent pathway. Pigment Cell Melanoma Res., 22, 623–634 (2009).
- 16) Singhmar P, Huo X, Li Y, Dougherty PM, Mei F, Cheng X, Heijnen CJ, Kavelaars A. Orally active epac inhibitor reverses mechanical allodynia and loss of intraepidermal nerve fibers in a mouse model of chemotherapy-induced peripheral neuropathy. Pain, 159, 884–893 (2018).
- 17) Brain PF, Evans AE. Acute influences of some ACTH-related peptides on fighting and adrenocortical activity in male laboratory mice. Pharmacol. Biochem. Behav., 7, 425–433 (1977).
- 18) Kumar A, Negi G, Sharma SS. JSH-23 targets nuclear factor-kappa B and reverses various deficits in experimental diabetic neuropathy: effect on neuroinflammation and antioxidant defence. Diabetes Obes. Metab., 13, 750–758 (2011).
- 19) Hiramoto K, Yamate Y, Shirasawa T, Sato EF. The role of the active oxygen product from gp91phox NADPH oxidase on the newborn weight of mouse pups. Biol. Med. (Aligarh), 7, 5 (2015).
- 20) Yokoyama S, Hiramoto K, Koyama M, Ooi K. Skin disruption is associated with indomethacin-induced small intestinal injury in mice. Exp. Dermatol., 23, 659–663 (2014).
- 21) Jimbow K, Uesugi T. New melanogenesis and photobiological processes in activation and proliferation of precursor melanocytes after UV-exposure: ultrastractural differentiation of precursor melanocytes from Langerhans cells. J. Invest. Dermatol., 78, 108–115 (1982).
- 22) Mac Neil S, Buffery J, Hill SE, Dobson J, Bleehen SS. Intracellular signalling in the control of melanogenesis. Pigment Cell Res., 3 (Supp2), 154–161 (1992).
- 23) Giménez García RM, Carrasco Molina SC. Drug-induced hyperpigmentation: review and case series. J. Am. Board Fam. Med., 32, 628–638 (2019).
- 24) Gantz I, Fong TM. The melanocortin system. Am. J. Physiol. Endocrinol. Metab., 284, E468–E474 (2003).
- 25) Almahariq M, Tsalkova T, Mei FC, Chen H, Zhou J, Sastry SK, Schwede F, Cheng X. A novel EPAC-specific inhibitor suppresses pancreatic cancer cell migration and invation. Mol. Pharmacol., 83, 122–128 (2013).
- 26) Winters SJ, Vitaz T, Nowacki MR, Craddock DC, Silverman C, Lopez FA. Addison’s disease and pituitary enlargement. Am. J. Med. Sci., 349, 526–529 (2015).
- 27) Numazawa S, Sugihara K, Miyake S, Tomiyama H, Hida A, Hatsuno M, Yamamoto M, Yoshida T. Possible involvement of oxidative stress in 5-fluorouracil-mediated myelosuppression in mice. Basic Clin. Pharmacol. Toxicol., 108, 40–45 (2011).
- 28) Chan KH, Lam KS, Cheng OY, Kwan JS, Ho PW, Cheng KK, Chung SK, Ho JW, Guo VY, Xu A. Adiponectin is protective against oxidative stress induced cytotoxicity in amyloid-beta neurotoxicity. PLOS ONE, 7, e52354 (2012).
- 29) Itoi K, Jiang YQ, Iwasaki Y, Watson SJ. Regulatory mechanisms of corticotropin-releasing hormone and vasopressin gene expression in the hypothalamus. J. Neuroendocrinol., 16, 348–355 (2004).
- 30) Chida D, Nakagawa S, Nagai S, Sagara H, Katsumata H, Imaki T, Suzuki H, Mitani F, Ogishima T, Shimizu C, Kotaki H, Kakuta S, Sudo K, Koike T, Kubo M, Iwakura Y. Melanocortin 2 receptor is required for adrenal gland development, steroidogenesis, and neonatal gluconeogenesis. Proc. Natl. Acad. Sci. U.S.A., 104, 18205–18210 (2007).