Abstract
Newly synthesized proteins destined for the secretory pathway are folded and assembled in the endoplasmic reticulum (ER) and then transported to the Golgi apparatus via COPII vesicles, which are normally 60–90 nm. COPII vesicles must accordingly be enlarged to accommodate proteins larger than 90 nm, such as long-chain collagen. Key molecules involved in this enlargement are Tango1 and Tango1-like (Tali), which are transmembrane proteins in the ER encoded by the MIA3 and MIA2 genes, respectively. Interestingly, two splicing variants are expressed from each of these two genes: Tango1L and Tango1S from the MIA3 gene, and Tali and cTAGE5 from the MIA2 gene. Here, we constructed Tango1L-knockout (KO), Tango1S-KO, Tali-KO, and cTAGE5-KO separately in medaka fish, a vertebrate model organism, and characterized them. Results showed that only Tango1L-KO conferred a lethal phenotype to medaka fish. Only Tango1L-KO medaka fish exhibited a shorter tail than wild-type (WT) fish and showed the defects in the export of type II collagen from the ER, contrary to the previous reports analyzing Tango1-KO or Tali-KO mice and the results of knockdown experiments in human cultured cells. Medaka fish may employ a simpler system than mammals for the export of large molecules from the ER.
Key words: intracellular transport, COPII vesicles, enlargement, endoplasmic reticulum, Golgi apparatus
Introduction
Newly synthesized proteins destined for the secretory pathway are folded and assembled in the ER with assistance from ER-localized molecular chaperones and folding enzymes, and only correctly folded molecules are allowed to move to the Golgi apparatus (Bukau et al., 2006). This transport from the ER is mediated by COPII vesicles, which are covered by two layers of protein complex, namely an inner coat consisting of Sec23 and Sec24 as well as an outer coat consisting of Sec13 and Sec31, and then depart from the ER exit site (see Fig. 1E). The size of COPII vesicles is normally 60–90 nm (Raote et al., 2023).

However, the cell sometimes needs to transport cargo proteins that are larger than 90 nm. For example, during formation of the notochord, notochord cells are smoothly aligned by synthesizing and secreting large amounts of extracellular matrix proteins, including type VIII collagen (Ishikawa et al., 2017). After secretion of hedgehog to regulate patterning of various surrounding tissues (Stemple, 2005), notochord cells receive the Mib-Jag1-Notch signal and then differentiate into two types of cells: one is large vacuolated structural cells that generate turgor and the other is thin non-vacuolated epithelial cells (sheath cells). As the notochord is a primitive form of cartilage, sheath cells synthesize and secrete type II collagen to form the peri-notochordal basement membrane, which covers the notochord as a sheath. Thanks to turgor generated by vacuolated cells and the sheath formed by sheath cells, the notochord extends smoothly and rigidly and thereby functions as an axial skeleton before the formation of vertebra (Ishikawa et al., 2017; Yamamoto et al., 2010).
Long-chain collagen such as type II collagen is longer than 300 nm and therefore cannot be incorporated into standard COPII vesicles. How such large cargo proteins could be transported was a major question in cell biology (Fromme and Schekman, 2005). In 2009, Vivek Malhotra and colleagues discovered an interesting and important molecule termed transport and Golgi organization (Tango) 1, which is localized in the ER as a transmembrane protein (Saito et al., 2009). Its N-terminal SH3 domain binds to long-chain collagen via Hsp47 in the lumen of the ER, whereas its C-terminal proline-rich domain (PRD) binds to Sec23/Sec24 in the cytosol, which blocks the completion of the formation of the outer coat layer, resulting in the enlargement of COPII vesicles to accommodate long-chain collagen (see Fig. 1E) (Malhotra and Erlmann, 2015).
Following the finding that a truncated version of Tango1 which lacked a majority of the N-terminal region was expressed in cultured cells, the full-length and truncated versions of Tango1 (note that they are splicing variants) were designated as Tango1L and Tango1S, respectively (Maeda et al., 2016; Wilson et al., 2011). Interestingly, cutaneous T-cell lymphoma-associated antigen 5 (cTAGE5) was found to be substantially homologous to Tango1S, and accordingly its full-length version was named Tango1-like (Tali); Tali and cTAGE5 are also splicing variants (Santos et al., 2016). Since then, the roles of Tango1L/Tango1S and Tali/cTAGE5 in the export of long-chain collagen as well as triglyceride-rich chylomicron (~150–500 nm in diameter) have been extensively investigated using cultured cells (Raote et al., 2021; Raote et al., 2023).
At the animal levels, Tango1-knockout (KO) mice were constructed and characterized (Wilson et al., 2011). Because a region containing the N-terminal SH3 domain was deleted, these Tango1-KO mice did not express Tango1L but did express Tango1S. These Tango1L-KO mice showed defects in the secretion of various collagens from chondrocytes, fibroblasts, endothelial cells, and mural cells; exhibited short-limbed dwarfism; and died at birth. On the other hand, Tali/cTAGE5-KO mice were constructed by deleting a cytoplasmic small region inside the shared coiled-coiled domain (CCD) 2, which lead to the loss of functionally important PRD capable of binding to Sec23/Sec24 in both Tali and cTAGE5. These KO mice showed embryonic lethalality (Wang et al., 2016).
In zebrafish (Clark and Link, 2021), as a majority of the N-terminal luminal region was deleted, Tango1-KO zebrafish did not express Tango1L but did express Tango1S, similarly to Tango1L-KO mice. These Tango1L-KO zebrafish were shorter than WT fish during the embryonic period, showed craniofacial defects, and did not survive to adulthood. On the other hand, as a small region inside the shared CCD2 was deleted, Tali/cTAGE5-KO zebrafish expressed truncated Tali and cTAGE5 lacking functionally important PRD capable of binding to Sec23/Sec24, similarly to Tali/cTAGE5-KO mice. Tali/cTAGE5-KO zebrafish appeared to be normal during the embryonic period but became shorter than WT fish at 2 months after hatching, and died by 1 year, although WT and heterozygous fish lived longer than 3 years.
Here, we separately knocked out Tango1L, Tango1S, Tali, and cTAGE5 in medaka fish, a vertebrate model organism, using genome editing and characterized their phenotypes for the first time.
Results
Relationship between medaka Tango1 and Tali
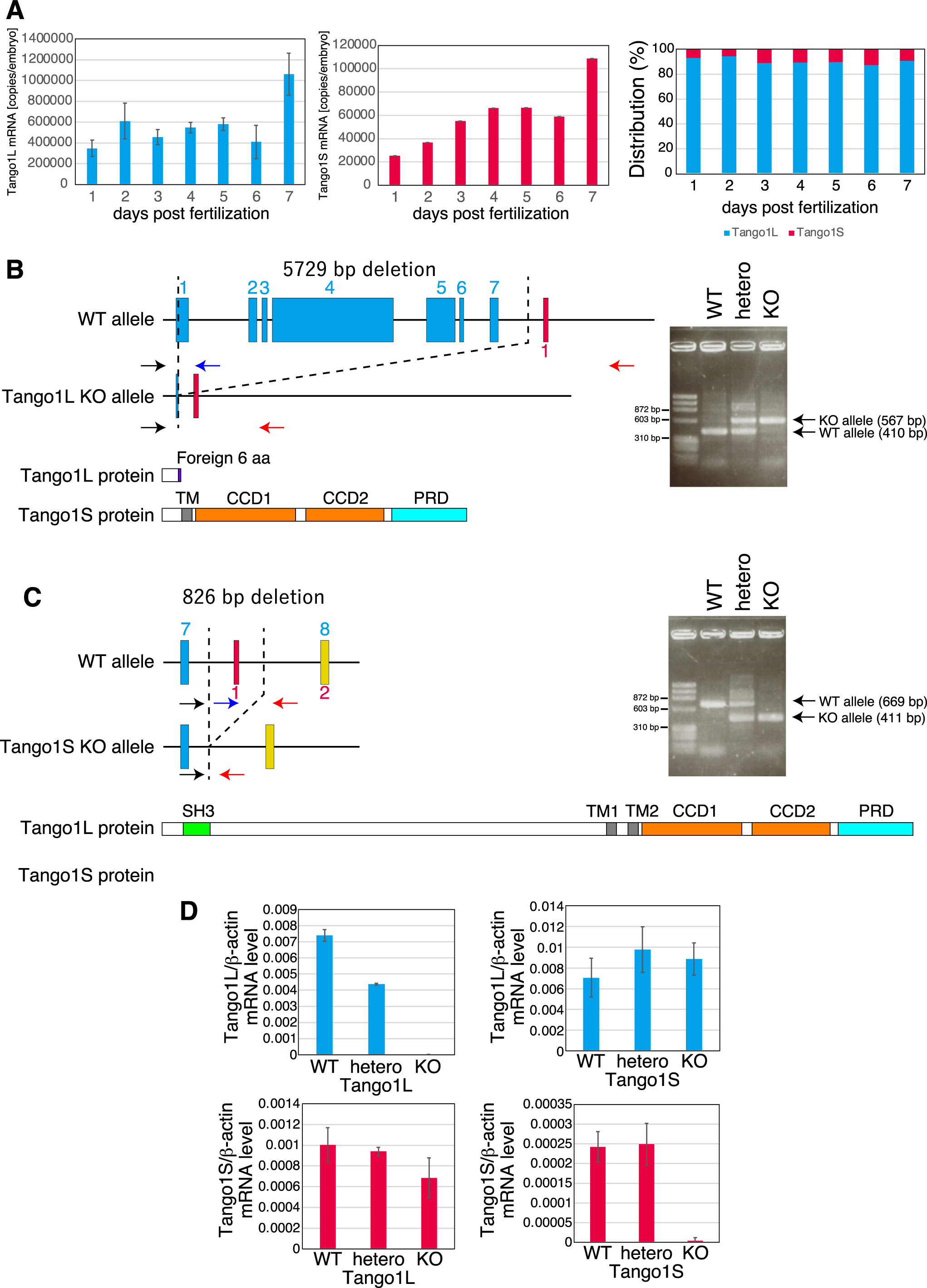
Two splicing variants are expressed from the MIA2 gene (MIA stands for melanoma inhibitory activity member) in medaka fish, namely Tali mRNA and cTAGE5 mRNA. Thus, the MIA2 locus possesses 6 Tali-specific exons, 1 cTAGE5-specific exon, and 22 shared exons from the 5' end to the 3' end (Fig. 1A). Similarly, two splicing variants are expressed from the MIA3 gene in medaka fish, namely Tango1L mRNA and Tango1S mRNA. Thus, the MIA3 locus possesses 6 Tango1L-specific exons, 1 Tango1S-specific exon, and 21 shared exons from the 5' end to the 3' end (Fig. 1B). Both Tali and Tango1L contain SH3, transmembrane (TM) domain, CCD1, CCD2, and PRD, whereas both cTAGE5 and Tango1S lack a majority of the luminal region present in Tali and Tango1L (Fig. 1A, B, E). Sequence comparison shows that human Tango1L is more similar to medaka Tango1L than to medaka Tali, whereas human Tali is more similar to medaka Tali than to medaka Tango1L (Fig. 1C, D).
Construction and characterization of Tali- and cTAGE5-knockout medaka
Determination of the absolute expression levels of Tali mRNA and cTAGE5 mRNA by quantitative RT-PCR using full-length Tali cDNA amplified by PCR as standard showed that cTAGE5 mRNA was more abundantly expressed than Tali mRNA in embryos from 1 to 7 days post-fertilization (dpf) (Fig. 2A, medaka usually hatch at 7 dpf).
We constructed Tali-KO medaka and cTAGE5-KO medaka using CRISPR/Cas9-mediated genome editing. To construct Tali-KO medaka, the exons 1–6 of the MIA2 gene were deleted entirely, which was confirmed by genomic PCR (Fig. 2B). This Tali-KO medaka did not express Tali mRNA but did express cTAGE5 mRNA as expected, when examined at 7 months post-hatching (mph) (Fig. 2D; medaka gain reproductive capacity at 2~3 mph). To construct cTAGE5-KO medaka, the cTAGE5-specific exon of the MIA2 gene was deleted, which was confirmed by genomic PCR (Fig. 2C). This cTAGE5-KO medaka expressed Tali mRNA but did not express cTAGE5 mRNA at 7 mph, as expected (Fig. 2D).
A great majority of Tali-KO medaka and cTAGE5-KO medaka hatched (Fig. 3A), and survived until 15 dpf (Fig. 3B) as well as 2 mph (Fig. 3C). Both Tali-KO medaka and cTAGE5-KO medaka exhibited normal tail length (Fig. 3D).
Construction and characterization of Tango1L- and Tango1S-knockout medaka
Determination of the absolute expression levels of Tango1L mRNA and Tango1S mRNA by quantitative RT-PCR using a plasmid carrying Tango1 cDNA as standard showed that approximately 90% were Tango1L mRNA in embryos from 1 to 7 dpf (Fig. 4A).
We constructed Tango1L-KO medaka and Tango1S-KO medaka using CRISPR/Cas9-mediated genome editing. To construct Tango1L-KO medaka, the exons 1–7 of the MIA3 gene were deleted almost entirely, which was confirmed by genomic PCR (Fig. 4B). This Tango1L-KO medaka did not express Tango1L mRNA but did express Tango1S mRNA, as expected (Fig. 4D). To construct Tango1S-KO medaka, the Tango1S-specific exon of the MIA3 gene was deleted, which was confirmed by genomic PCR (Fig. 4C). This Tango1S-KO medaka expressed Tango1L mRNA but did not express Tango1S mRNA, as expected (Fig. 4D).
Approximately two-thirds of Tango1L-KO medaka did not hatch (Fig. 5A). Hatched Tango1L-KO medaka started to die from 11 dpf and all died by 14 dpf (Fig. 5B, C). In contrast, a great majority of Tango1S-KO medaka hatched (Fig. 5A) and survived until 15 dpf (Fig. 5B). However, it should be noted that Tango1S-KO medaka survived for 6 mph did not show the Mendelian ratio (Fig. 5C). Tango1L-KO medaka but not Tango1S-KO medaka exhibited a shorter tail than WT fish (Fig. 5D).
Export of type II collagen
As only Tango1L-KO medaka exhibited several phenotypes, immunofluorescence analysis of type II collagen was conducted with comparison of calnexin (CNX), an ER marker, in embryos with various genotypes of Tango1L and Tango1S, focusing on the region containing the notochord. Type II collagen was secreted to form the peri-notochordal basement membrane immediately outside CNX-positive ER in Tango1L-WT, Tango1L-hetero, Tango1S-WT, Tango1S-hetero, and Tango1S-KO medaka (Fig. 6A, B). Notably, type II collagen remained punctate and the peri-notochordal basement membrane was not formed in Tango1L-KO medaka (Fig. 6A, panel k).
Discussion
Two splicing variants are expressed from the MIA3 gene, namely Tango1L mRNA and Tango1S mRNA. Our present analysis revealed that Tango1L but not Tango1S is required for the export of type II collagen from the ER (Fig. 6). This is in contrast to previous Tango1S knockdown experiments, which showed that Tango1S is also required for the secretion of type VII collagen from the ER in human cultured cells (Maeda et al., 2016). This discrepancy may be due to the difference in their expression levels: Tango1L mRNA is approximately 9-fold more abundant than Tango1S mRNA in medaka fish (Fig. 4A), whereas immunoblotting showed that Tango1S is abundantly expressed in addition to Tango1L in human cultured cells (Maeda et al., 2016). Considering that Tango1L and Tango1S are interchangeable in human cultured cells (Maeda et al., 2016), medaka Tango1L alone would be able to initiate the enlargement of COPII vesicles to accommodate long-chain collagen without making heterodimers with Tango1S. Accordingly, the absence of Tango1L hampered the formation of the peri-notochordal basement membrane, leading to a short tail (Fig. 5D), because turgor generated by vacuolated cells could not be efficiently utilized to extend the notochord smoothly and rigidly.
Nonetheless, given that Tango1S-KO medaka surviving for 2 months after hatching did not show the Mendelian ratio (Fig. 5C), our study might have underestimated the importance of Tango1S. Resolution of this question requires the construction and analysis of Tango1L/Tango1S-double KO medaka. Also, the cause of the death of Tango1L-KO medaka within 14 dpf should be determined, because it cannot be explained solely by short tail. Furthermore, the question of whether the transport defects in Tango1L-KO medaka could be rescued by overexpression of Tango1S, Tali, or cTAGE5 needs to be answered.
Two splicing variants are expressed from the MIA2 gene, namely Tali mRNA and cTAGE5 mRNA. Our current analysis revealed that neither Tali nor cTAGE5 is required for survival (Fig. 3) and for the export of type II collagen from the ER (data not shown) in medaka fish. This is surprising because it was previously shown that cTAGE5 heterodimerizes with Tango1L and is required for the export of type VII collagen in human cultured cells (Saito et al., 2011). As mentioned above, Tango1L is required for the export of type II collagen in medaka fish, but does not seem to rely on a partner(s) for its functionality in this fish. Biochemical analysis is required to understand the difference between mammalian cells and medaka fish; i.e., the importance of heterodimerization in the export of long-chain collagen from the ER. Further, a conclusive determination of the role of Tali/cTAGE5 in the physiology of medaka fish requires the construction and analysis of Tali/cTAGE5-double KO medaka, as was done in zebrafish (Clark and Link, 2021).
Materials and Methods
Statistics
Statistical analysis was conducted using Student’s t-test, with probability expressed as *p<0.05, **p<0.01 and ***p<0.001 for all figures.
Fish
Medaka southern strain Cab was used as WT fish. Fish were maintained in a recirculating system with a 14:10 hr light:dark cycle at 27.5°C. All experiments were performed in accordance with the guidelines and regulations established by the Animal Research Committee of Kyoto University (approval number: H2819).
CRISPR/Cas9 method
To synthesize sgRNAs for CRISPR/Cas9-mediated genome editing, a PCR product amplified from DR274 vector (Addgene) using T7-sgRNA-Fw and DR274-Rv primers was purified by phenol chloroform extraction and used as template for T7 RNA polymerase. The sequences of transcribed sgRNAs are shown in Table 1.
Table 1
Information of DNA sequences and antibodies
| sgRNAs |
| Target |
Sequence |
| ol Tango1L-up |
GGGGUUCGCAUCGAUGGAUGGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| olTango1L-down |
GGCGUGUGCUUUCACGGUGAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol Tango1S-up |
GGCGUGUGCUUUCACGGUGAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol Tango1S-down |
GGCCUGCUACUAUAGUUCAAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol Tali-up |
GGGCUUCUGCGGUUGAAUUGGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol Tali-down |
GGAUUGGCACCUGUUUCCUCGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol cTAGE5-up |
GGAUUCGCCGUUAACCCAGCGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| ol cTAGE5-down |
GGUUCUCCUGUUCGGCUAACGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU |
| Primers for genotyping |
| Target |
Sequence |
| ol Tango1L |
GAATGAGGTCGTGCTTTCCAGTCTTCGTG |
| CATGTGATCTGGTTCCGCTACAGGTTGCTG |
| CTCCCGTATCAGGATCAACGGAAGGGAAC |
| ol Tango1S |
CATCACGTCTCTCATCAAACGTTGTCAGTG |
| CCATCCTATTTAGCTTGATTGGTTTCAGGC |
| ol Tali |
GGTGTCTTGTCCTAACTCCACCGGAAGCAG |
| GTGACCGTCAAGCATGTACCACAACACTTC |
| CAACGTTGCTCGTAACCTGATTGCAGAAC |
| ol cTAGE5 |
GTTAACCCAGCTGGCAGCTCTACTTTCTAC |
| ATTCAGCATAAGAAACAGCTAACTACTAAC |
| Primers for qRT-PCR |
| Target |
Sequence |
| ol Tango1L-Fw |
CCTGAGCTTGAGCCTTCATT |
| ol Tango1L-Rv |
TCCCACTCCTTCCTCTTGTT |
| ol Tango1S-Fw |
GTAGCGGAACCAGATCACAT |
| ol Tango1S-Rv |
CGGCTTCCACTCTTCTGGTA |
| ol Tali-Fw |
GGAGATTCTGAGTGTGAAAGCCTAATGA |
| ol Tali-Rv |
CTTAGAAAACGGCAGTCGTTACTCTTGT |
| ol cTAGE5-Fw |
ACCCGCGGGCCCACCAGGAAGAGCAGGCACAGACC |
| ol cTAGE5-Rv |
ATTACTTGCGGCCGCTTCTGCTGTTTCCAGGTTCCTCTG |
| Antibody |
| Immunogen |
Source |
Identifier |
| Anti-Collagen II antibody |
abcam |
ab185430 |
| Anti-Calnexin antibody |
Enzo Life Sciences |
ADI-SPA-865-F |
| anti-Mouse IgG (H + L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 488 |
Thermo Fisher Scientific |
A-11017 |
| anti-Rabbit IgG (H + L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 568 |
Thermo Fisher Scientific |
A-11011 |
Cas9 expression vector was linearized with NotI, purified by phenol-chloroform extraction, and used as template to synthesize capped mRNAs using the Message mMachine SP6Kit (Invitrogen).
Synthesized RNAs were purified by phenol chloroform extraction (for sgRNAs) or RNeasy mini kit (Qiagen) (for Cas9 mRNA) and microinjected into one-cell-stage embryos at a concentration of 1 μg/μl for Cas9 and 25 ng/μl for both upstream and downstream targeting sgRNAs. Injection was performed as described previously (Ishikawa et al., 2011).
Genotyping
Embryos or hatched fish were suspended in 50 μl of lysis buffer (10 mM NaOH and 0.2 mM EDTA), boiled for 10 min, and then neutralized by the addition of an equal volume of 40 mM Tris-HCl, pH 8.0. DNA fragment containing a part of the MIA2 or MIA3 gene was amplified by PCR directly from lysates using the primers shown in Table 1.
Quantitative RT-PCR
Total RNA was extracted from embryos or caudal fin at the indicated dpf or mph by the acid guanidinium-phenol-chloroform method using Isogen (Nippon Gene). Quantitative RT-PCR analysis was carried out as described previously (Ishikawa et al., 2013) using the SYBR Green method (Applied Biosystems) and a pair of primers (Fw and Rv) whose names and sequences are described in Table 1.
Microscopy and Tail length measurement
Brightfield microscopic analysis was conducted using Leica M205FA stereomicroscope.
Tail length was determined by analyzing microscopic images using ImageJ (https://imagej.nih.gov/ij/).
Immunofluorescence
Embryos were fixed in 4% paraformaldehyde in PBS overnight at 4°C, washed with PBS, fixed again with 100% methanol for 15 min at room temperature and more than overnight at –30°C, and then washed with PBST (0.1% Tween20 in PBS). Fixed embryos were permeabilized by incubation with 0.1 mg/ml proteinase K in PBSX (0.1% Triton X-100 in PBS) for 22 min at room temperature and washed with PBSDX (1% DMSO and 0.3% Triton X-100 in PBS) three times. Permeabilized embryos were blocked in blocking solution (2% fetal bovine serum and 2 mg/ml BSA in PBSX) for 4 h at 4°C, reacted with primary antibody shown in Table 1 in blocking solution overnight at 4°C, washed two times with PBSDX, reacted with secondary antibody shown in Table 1 and Hoechst 33342 for 1 h at room temperature, and then washed with PBSDX. Immunostained embryos were analyzed using Zeiss LSM880 microscopy.
Acknowledgments
We thank Ms. Kaoru Miyagawa and Ms. Makiko Sawada for their technical and secretarial assistance. This work was financially supported in part by AMED-CREST, Japan (24gm1410005 to K. M.).
References
- Bukau, B., Weissman, J., and Horwich, A. 2006. Molecular chaperones and protein quality control. Cell, 125: 443–451.
- Clark, E.M. and Link, B.A. 2021. Complementary and divergent functions of zebrafish Tango1 and Ctage5 in tissue development and homeostasis. Mol. Biol. Cell, 32: 391–401.
- Fromme, J.C. and Schekman, R. 2005. COPII-coated vesicles: Flexible enough for large cargo? Curr. Opin. Cell Biol., 17: 345–352.
- Ishikawa, T., Okada, T., Ishikawa-Fujiwara, T., Todo, T., Kamei, Y., Shigenobu, S., Tanaka, M., Saito, T.L., Yoshimura, J., Morishita, S., Toyoda, A., Sakaki, Y., Taniguchi, Y., Takeda, S., and Mori, K. 2013. ATF6alpha/beta-mediated adjustment of ER chaperone levels is essential for development of the notochord in medaka fish. Mol. Biol. Cell, 24: 1387–1395.
- Ishikawa, T., Taniguchi, Y., Okada, T., Takeda, S., and Mori, K. 2011. Vertebrate unfolded protein response: Mammalian signaling pathways are conserved in medaka fish. Cell Struct. Funct., 36: 247–259.
- Ishikawa, T., Toyama, T., Nakamura, Y., Tamada, K., Shimizu, H., Ninagawa, S., Okada, T., Kamei, Y., Ishikawa-Fujiwara, T., Todo, T., Aoyama, E., Takigawa, M., Harada, A., and Mori, K. 2017. UPR transducer BBF2H7 allows export of type II collagen in a cargo- and developmental stage-specific manner. J. Cell Biol., 216: 1761–1774.
- Maeda, M., Saito, K., and Katada, T. 2016. Distinct isoform-specific complexes of TANGO1 cooperatively facilitate collagen secretion from the endoplasmic reticulum. Mol. Biol. Cell, 27: 2688–2696.
- Malhotra, V. and Erlmann, P. 2015. The pathway of collagen secretion. Annu. Rev. Cell. Dev. Biol., 31: 109–124.
- Raote, I., Saxena, S., Campelo, F., and Malhotra, V. 2021. TANGO1 marshals the early secretory pathway for cargo export. Biochim. Biophys. Acta Biomembr., 1863: 183700.
- Raote, I., Saxena, S., and Malhotra, V. 2023. Sorting and export of proteins at the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol., 15: a041258.
- Saito, K., Chen, M., Bard, F., Chen, S., Zhou, H., Woodley, D., Polischuk, R., Schekman, R., and Malhotra, V. 2009. TANGO1 facilitates cargo loading at endoplasmic reticulum exit sites. Cell, 136: 891–902.
- Saito, K., Yamashiro, K., Ichikawa, Y., Erlmann, P., Kontani, K., Malhotra, V., and Katada, T. 2011. cTAGE5 mediates collagen secretion through interaction with TANGO1 at endoplasmic reticulum exit sites. Mol. Biol. Cell, 22: 2301–2308.
- Santos, A.J., Nogueira, C., Ortega-Bellido, M., and Malhotra, V. 2016. TANGO1 and Mia2/cTAGE5 (TALI) cooperate to export bulky pre-chylomicrons/VLDLs from the endoplasmic reticulum. J. Cell Biol., 213: 343–354.
- Stemple, D.L. 2005. Structure and function of the notochord: An essential organ for chordate development. Development, 132: 2503–2512.
- Wang, Y., Liu, L., Zhang, H., Fan, J., Zhang, F., Yu, M., Shi, L., Yang, L., Lam, S.M., Wang, H., Chen, X., Wang, Y., Gao, F., Shui, G., and Xu, Z. 2016. Mea6 controls VLDL transport through the coordinated regulation of COPII assembly. Cell Res., 26: 787–804.
- Wilson, D.G., Phamluong, K., Li, L., Sun, M., Cao, T.C., Liu, P.S., Modrusan, Z., Sandoval, W.N., Rangell, L., Carano, R.A., Peterson, A.S., and Solloway, M.J. 2011. Global defects in collagen secretion in a Mia3/TANGO1 knockout mouse. J. Cell Biol., 193: 935–951.
- Yamamoto, M., Morita, R., Mizoguchi, T., Matsuo, H., Isoda, M., Ishitani, T., Chitnis, A.B., Matsumoto, K., Crump, J.G., Hozumi, K., Yonemura, S., Kawakami, K., and Itoh, M. 2010. Mib-Jag1-Notch signalling regulates patterning and structural roles of the notochord by controlling cell-fate decisions. Development, 137: 2527–2537.
Abbreviations
CCDcoiled-coiled domain
dpfday post-fertilization
dphday post-hatching
mphmonth post-hatching
ERendoplasmic reticulum
KOknockout
PRDproline-rich domain
TMtransmembrane
WTwild-type