Abstract
Understanding fruit growth and thermal requirements from anthesis to ripening can provide insights into the developmental process and productivity of whole plants. Fruit growth in sweet peppers (Capsicum annuum) can be estimated using sigmoid functions. The present study aimed to clarify the growth patterns of sweet pepper fruit using these functions. In a greenhouse, we hydroponically grew 10 sweet pepper cultivars that have a wide range of fruit dry matter content. Fruits at different growth stages were destructively sampled and measured for their length, width, dry and fresh weights, as well as dry matter content. Using the data, fruit growth curves in terms of length, width, water content, dry weight, and dry matter content were modeled using the Gompertz function. Results showed that fruit volume (i.e., fruit length and width), fruit water content, and fruit dry weight rapidly increased from 200–600, 400–800, and 600–1,200°C·d days after anthesis (DAA), respectively, in all the cultivar types. However, the increase in fruit dry weight from 600–1,200°C·d DAA differed among the cultivar types. Fruit dry matter content decreased to 0.04–0.05 g·g−1 until about 600°C·d DAA regardless of the cultivar types, and thereafter it increased from 600–1,200°C·d DAA for the cultivars with high dry weight increase, but it barely increased for the cultivars with low dry weight increase. In conclusion, sweet pepper fruit initially began to expand in volume, followed by an increase in water content and dry weight. However, the increase in the fruit dry weight from 600–1,200°C·d DAA showed differences among the cultivar types, and this trait determined the cultivar-specific fruit dry matter content at harvest. The cultivar-specific dry matter increase affects the fruit load; therefore, it is probably related to the number of harvestable fruits (i.e., fresh yield) and plant exhaustion during production. The results of this study are expected to enable more precise growth prediction and plant management.
Introduction
Determining fruit growth and thermal requirements from anthesis to ripening are crucial for understanding the developmental process of whole plants and improving production efficiency. Sigmoid functions have been used to model the daily fruit growth of horticultural crops such as strawberry (Fragaria × ananassa; Saito et al., 2022), tomato (Solanum lycopersicum; Adams et al., 2001; Ho et al., 1987; Watabe et al., 2022), sweet pepper (Capsicum annuum; Marcelis and Baan Hofman-Eijer, 1995; Wubs et al., 2012), pear (Pyrus communis; Mitchell, 1986), and peach (Prunus persica Batsch; Génard and Souty, 1996). Additionally, some studies used these fruit growth curves to predict fruit size (Bozokalfa and Kilic, 2010; Ngouajio et al., 2003; Wubs et al., 2012), fresh fruit yield (Jones et al., 1991), harvest date (Wolf et al., 1986), fruit set (Homma et al., 2022), and fruit load (Saito et al., 2022). Introducing these plant-growth factors in production sites is expected to improve labor management and production efficiency (Higashide, 2022). Thus, fruit growth models help to clarify whole plant growth and improve productivity.
The growth patterns of fruits were different depending on the crop. For example, dry weight of sweet pepper fruit gradually increased until harvest, but the fresh weight barely increased in the later growth stage (Marcelis and Baan Hofman-Eijer, 1995; Wubs et al., 2012). In contrast, Adams et al. (2001) and Watabe et al. (2022) found that dry weight of tomato fruit barely increased during the later growth stage, but its fresh weight and volume gradually increased until harvest. Although immature cucumber fruits were harvested, Marcelis (1992) reported that fresh weight, but not dry weight, gradually increased until harvest. In peach, Fukuda et al. (2021) reported that fruit length (i.e., fruit volume) was represented by a double-sigmoid curve. Thus, fruit growth patterns vary in terms of volume, dry weight, and water content.
Further investigations of these fruit-growth factors help to predict not only fresh yield and fruit quality but also root and shoot growth. Initially, increases in fruit dry weight and water content are related to fruit dry matter content at harvest (i.e., fruit dry weight/[fruit water content + fruit dry weight]). The fruit dry matter content is closely related to soluble solids within the fruit (i.e., fruit quality) and fresh yields (Higashide and Heuvelink, 2009; Higashide et al., 2012). In horticultural crops such as tomato and sweet pepper, fresh yields were different in various cultivars owing to cultivar-specific fruit dry matter content (Higashide et al., 2012; Homma et al., 2024b). Therefore, crop- or cultivar-specific increases in fruit dry weight and water content may influence fruit dry matter content at harvest, thereby affecting yield potential and fruit quality. In sweet pepper, fruit load calculated using the growth of fruit dry weight rather than fruit fresh weight or volume was associated with whole plant growth, including root and shoot growth (González-Real et al., 2008; Hall, 1977). Thus, fruit growth analyses, including volume, dry weight, water content, and dry matter content, can provide insights into not only individual fruit growth but also whole plant growth. However, few studies have reported analyses focusing on these fruit-growth factors, especially using multiple cultivars with different fruit dry matter content values.
Fruit dry matter content of sweet pepper has high and low values under slightly high and low fertilizer supply (Bae and Kim, 2004; Tadesse et al., 1999); however, in hydroponic production, growers can maintain nutrient solutions in the optimal range for plant growth. The differences in fruit dry matter content can thus be considered as cultivar differences. Consequently, clarifying the cultivar-specific fruit dry matter content focused on fruit-growth factors is crucial to understand individual fruit and whole plant growth that are in turn related to productivity.
Using sweet pepper as a case study, the present study aimed to clarify fruit growth patterns with a focus on volume, dry weight, water content, and dry matter content by using 10 cultivars with a wide range of fruit dry matter content values. Fruits at different developmental stages were destructively sampled, and the length, width, dry and fresh weights, and dry matter content were measured. Subsequently, fruit growth patterns were determined and discussed in relation to cultivar differences.
Materials and Methods
Plant materials and growth conditions
Sweet pepper plants were hydroponically grown in a small compartment of a Venlo-type greenhouse (width, 18 m; length, 18 m; height, 5.1 m) in Tsukuba, Japan (36°26′N, 140°10′E). The following red and yellow cultivars were used in this study: ‘Sven’, ‘Fairway’, ‘Mavera’, ‘Morbidelli’, ‘Keessie’, ‘Kaite’, ‘Gialte’, ‘Maldonado’, ‘Nagano’, and ‘Artega’. The ‘Artega’, ‘Maldonado’, ‘Mavera’, ‘Gialte’, and ‘Kaite’ were obtained from Enza Zaden (Enkhuizen, the Netherlands), while the ‘Nagano’, ‘Keessie’, ‘Sven’, ‘Fairway’, and ‘Morbidelli’ were obtained from Rijk Zwaan (Zuid-Holland, the Netherlands). These cultivars were selected based on a relatively small to large fruit size and were characterized by a wide range of fruit dry matter content (Homma et al., 2024b). The greenhouse was divided into five rows with an inter-row width of approximately 1.8 m. Planting beds were arranged in a double-row planting system.
Sweet pepper seeds were sown on 18 August 2021 in seed trays. The seeds were germinated in nursery soil covered with an aluminum foil for three days at a 30°C air temperature. The seedlings were then grown under constant illumination using fluorescent lamps with a photosynthetic photon flux density of 400 μmol·m−2·s−1, a 16-h daytime (25°C)/8-h nighttime (20°C) cycle, and a 1,000 μmol·mol−1 CO2 atmosphere in a growth chamber (Nae-terrace, 4-6T; Mitsubishi Chemical Agri Dream Co., Ltd., Tokyo, Japan). The seedlings were fertilized every day with a commercial nutrient solution (High-Tempo; Sumitomo Chemical Co., Ltd., Tokyo, Japan) at an adjusted electrical conductivity of 1.8 dS·m−1. The solution comprised 9.20 mM NO3−, 3.07 mM H2PO4−, 5.61 mM K+, 1.67 mM Mg2+, and 4.96 mM Ca2+. After four weeks, the seedlings were transferred to the greenhouse and allowed to grow for a week.
On 21 September 2021, the seedlings were transplanted into rockwool cubes (Grodan Delta Block; Rockwool B.V., Limburg, the Netherlands) and placed onto rockwool slabs (Grotop Vital; Grodan, Roermond, the Netherlands) in the greenhouse. The plants were trained on two main stems. The planting density was 3.7 plants·m−2 (i.e., 7.4 stems·m−2). Weak laterals of each dichotomous branch above the first leaf were pruned once a week: i.e., each node had two leaves.
The aboveground environments were recorded and controlled using an integrated environmental control system (Maximizer; Priva. B.V., South Holland, the Netherlands). A system measurement box was set in center of the greenhouse at 2.0 m high. Ventilation windows automatically opened when the air temperature in the greenhouse exceeded 23°C. A heat pump (Green Package; Nepon Co., Ltd., Tokyo, Japan) was used for cooling at night when the air temperature exceeded 18–20°C. A fogging system (LYOHM system, CoolPescon CH; Ikeuchi Co., Ltd., Tokyo, Japan) was used to maintain a relative air humidity of approximately 70–85% during the daytime. A heater (House Kaonki; Nepon) was set to turn on when the air temperature dropped to 18°C at nighttime (from sunset to sunrise) and 21°C in the daytime (from sunrise to sunset). A shade curtain (SLS 50 Harmony; Svensson, Kinna, Sweden) was extended across the roof when the outside solar radiation reached 0.8–1.0 kW·m−2.
Plants were supplied with a commercial nutrient solution (OAT-SA; OAT Agrio Co., Ltd., Tokyo, Japan) at 2.0 dS·m−1 (i.e., optimal range for sweet pepper growth: 1.5–2.5 dS·m−1; Bae and Kim, 2004; Tadesse et al., 1999). The nutrient solution adjusted to an electrical conductivity of 2.6 dS·m−1 contained 17.1 mM NO3−, 4.4 mM H2PO4−, 10.2 mM K+, 1.5 mM Mg2+, and 4.1 mM Ca2+. Irrigation was controlled using a ubiquitous environmental control system (Hoshi et al., 2018). Irrigation frequency was determined by external solar radiation, and irrigation was controlled to maintain a daily drainage rate (drainage/supplied water) over 30%. The drainage rate was calculated based on the total water supply and drainage for 10 cultivars.
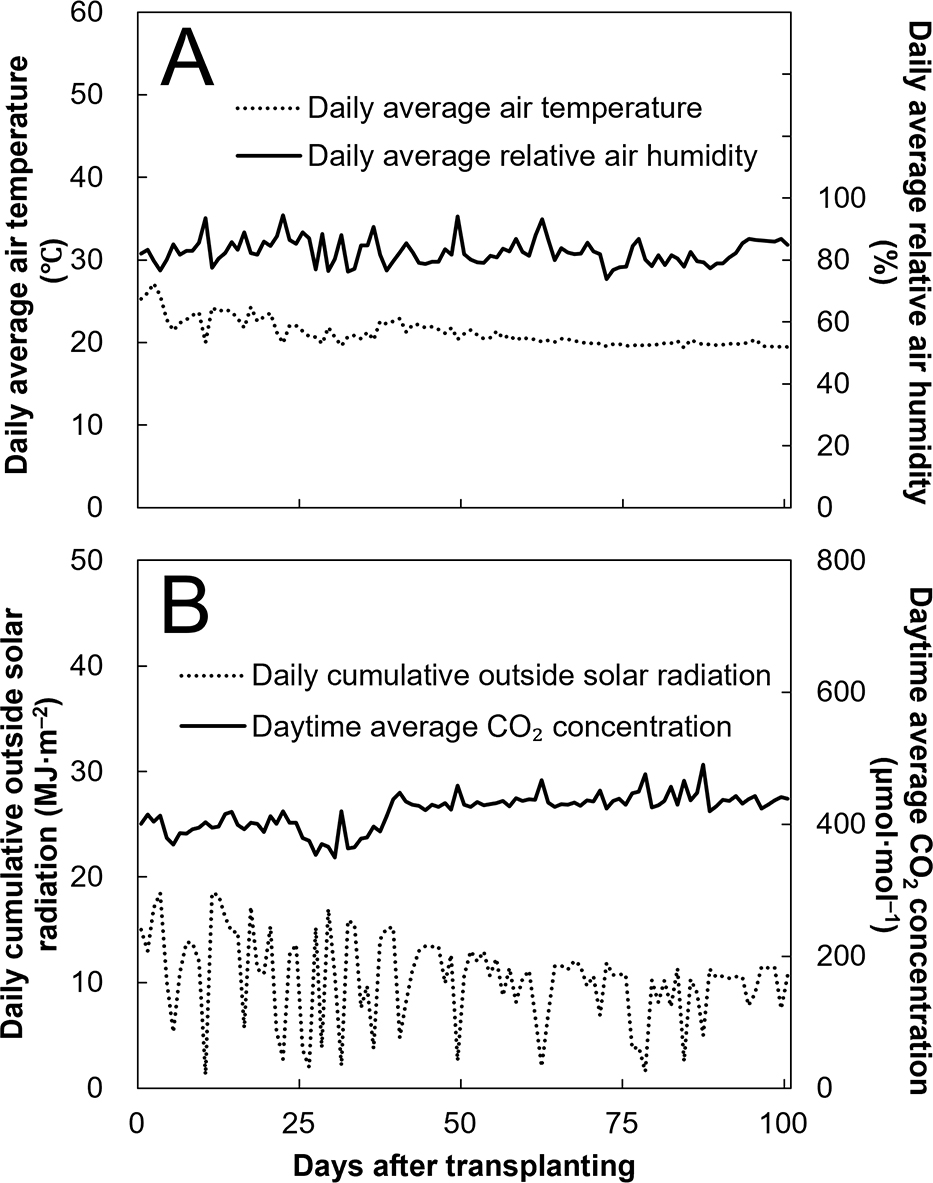
Changes in environmental conditions from 0–100 days after transplanting (DAT) are presented in Figure 1. Daily average air temperature, daily average relative air humidity, daily cumulative outside solar radiation, and daytime average CO2 concentration in the greenhouse were within the range of 21.2 ± 1.6°C, 82.7 ± 4.1%, 10.4 ± 4.1 MJ·m−2, and 420 ± 27 μmol·mol−1, respectively. Daily cumulative outside solar radiation decreased from 0 to 100 DAT. In 2021, the summer and winter solstices in Tsukuba, Japan, where the experiment was performed, were about 21 June (91 days before transplanting) and 22 December (92 DAT), respectively.
Measurements of fruit volume and weight and fitting them to a fruit growth function
To measure fruit growth curves, we first removed all flowers and growing fruits from the plants at 20 DAT, as reported to Chen et al. (2024). Thereafter, only one fruit was allowed to grow on each plant from 27–97 DAT. The fruits were set only on the main stem of the plant. The observed leaf area indices (LAI, m2·m−2) of ‘Artega’ at 0 and 80 DAT were about 0.1 and 4.4 m2·m−2, respectively (data not shown). The plant management applied here (i.e., two leaves per node) led to a linear increase in LAI values, as previously reported (Homma et al., 2022). Therefore, the fruit grew with an LAI of approximately 1.0–5.0 m2·m−2 from 27–97 DAT. We believe that these values were sufficient to grow only one fruit per plant.
Fruit set was defined as when fruit grew over 1.0–2.0 cm wide after anthesis. This definition was based on previous findings that sweet pepper flowers barely abort after 1–2 weeks of anthesis and fruit width reaches 1.0–2.0 cm in this period (Homma et al., 2022, 2024a; Wubs et al., 2007). Each week, approximately five fruits were allowed to set on 5–7 plants, and some fruits were aborted after fruit set. Thus, about 0–7 fruits per week set on the plants of each cultivar, and fruits of different ages grew from 27 to 97 DAT (26–48 fruits per cultivar).
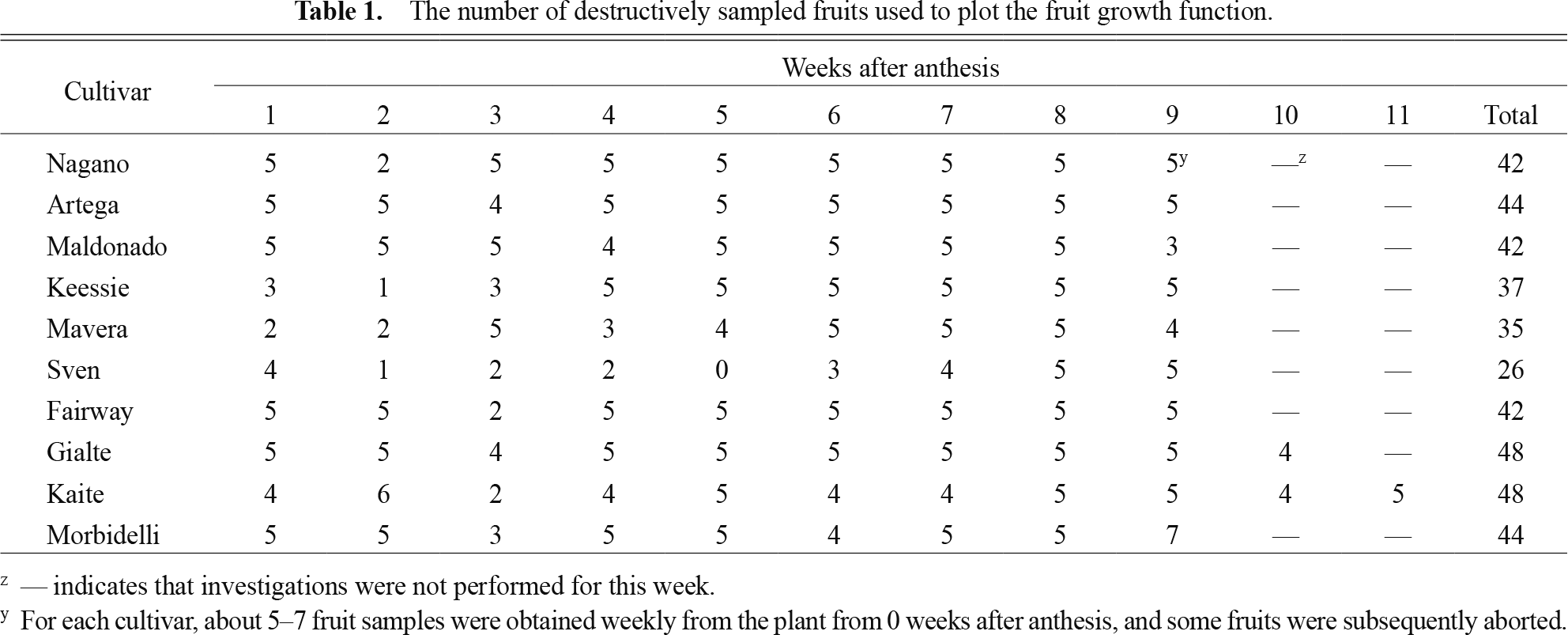
Some fruits aborted in this experiment; therefore, fruit samples were obtained for 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, and 11 weeks from anthesis (Table 1). Growing fruits of different ages were destructively sampled at 87 DAT (‘Fairway’, ‘Morbidelli’, ‘Keessie’, ‘Maldonado’, and ‘Artega’), 90 DAT (‘Mavera’, ‘Kaite’, ‘Gialte’, and ‘Nagano’), and 97 DAT (‘Sven’) for fruit growth modeling. Thus, fruit set investigations were not performed from 91–97 DAT for the nine destructively sampled cultivars at 87 and 90 DAT.
Fruit length (FL, mm per fruit), fruit width (FW, mm per fruit), fruit fresh weight (FFW, g per fruit), and fruit dry weight (FDW, g per fruit) were measured using a digital scale (TE1502S; Sartorius, Goettingen, Germany) and calipers (Digital Nogisu; Niigata Seiki Co., Ltd., Niigata, Japan). In this study, FFW and FDW were measured for only fruit without stalks. We measured not only individual fruit weight but also fruit width and length, because both indicators are used as criteria for categorizing fruit size (e.g., fruit weight: Japan, Vegetable Information, 2019; fruit length: United States, USDA, 2005). The sampled fruits were dried for at least 72 h at 100°C using a ventilation drier (JMB-28DPN-S; Maruto Testing Machine Co., Ltd., Tokyo, Japan) to measure the dry weight.
The fruit growth curve was modeled using sigmoid growth according to Wubs et al. (2012). This function showed good estimation of fruit size in previous studies that used sweet pepper (Homma et al., 2022, 2024a); therefore, we applied this model for estimating sweet pepper fruit size.
|
F
L
,
F
W
,
F
D
W
,
F
F
W
=
A
×
b
e
−
c
×
x
| (1) |
where the coefficients A, b, and c represent an upper asymptote line (FL and FW, mm per fruit; FDW and FFW, g per fruit), intercept of the function (dimensionless), and constant determination of the curvature (1/°C·d), respectively. The x value is the cumulative daily average air temperature after anthesis (°C·d). Anthesis was defined as the state in which the petals were completely opened and pollen was produced (Wubs et al., 2007). In each cultivar, cumulative daily average air temperatures after anthesis to the fruit reaching a length of approximately 1.0–2.0 cm were measured for 3–6 fruits after 97 DAT (‘Artega’: n = 3, 138°C·d, ‘Nagano’: n = 3, 217°C·d, ‘Maldonado’: n = 3, 158°C·d, ‘Keessie’: n = 6, 159°C·d, ‘Mavera’: n = 3, 138°C·d, ‘Sven’: n = 5, 159°C·d, ‘Fairway’: n = 5, 217°C·d, ‘Gialte’: n = 3, 138°C·d, ‘Kaite’: n = 3, 158°C·d, and ‘Morbidelli’: n = 3, 178°C·d). Therefore, the x value in each fruit sample was calculated by adding the cumulative average air temperatures from anthesis until fruit set (138–217°C·d) and those from fruit set until harvest (20–1,475°C·d). Regression curves were obtained by constructing scatter plots between the observed fruit parameters and corresponding air temperatures from anthesis to the sampling date. This study sampled fruits about 1,400–1,500°C·d so they had completely matured. We set 1,200°C·d as the harvest period based on a previous study (Homma et al., 2023b), which showed that completely matured fruits had 1,150–1,250°C·d.
Fruit growth curves of dry matter content (FDMC, g·g−1, Eq. 2) and water content (FWC, g per fruit, Eq. 3) were calculated as follows:
|
F
D
M
C
=
F
D
W
/
F
F
W
| (2) |
|
F
W
C
=
F
F
W
−
F
D
W
| (3) |
Standardized fruit growth curves were obtained by dividing Eq. 1 by the coefficient A obtained here. Instantaneous fruit growth curves were obtained as derivatives of the standardized curves.
Statistical analysis
The significance of the fitted sigmoid function was investigated by calculating P-values for the parameters. To detect cultivar-specific variations in fruit growth curves (i.e., volume, water content, dry weight, and fruit dry matter content), fruit growth functions for each cultivar are shown using the obtained parameters. Statistical analyses were performed using R version 4.1.0 (R Development Core Team, 2020).
Results
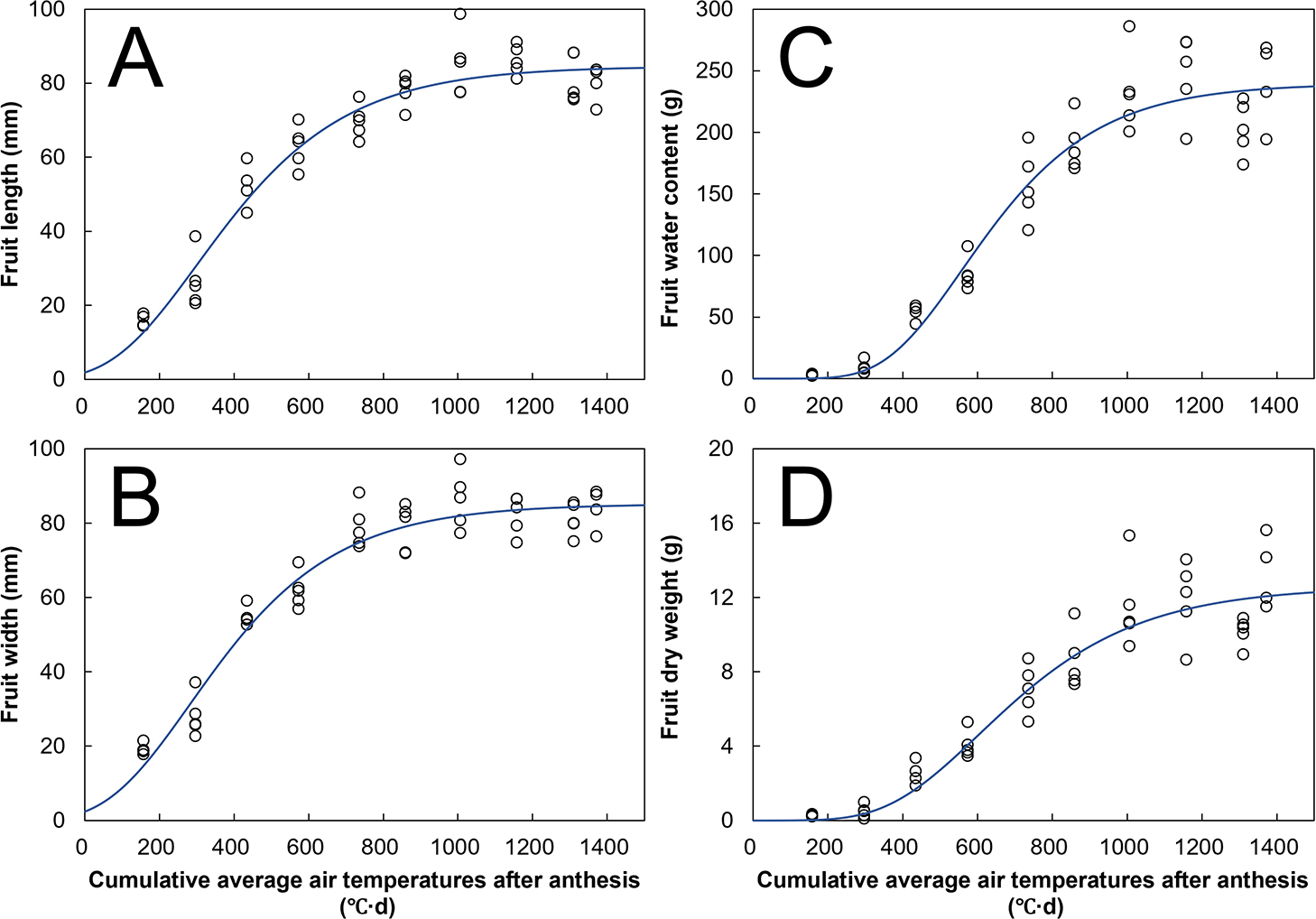
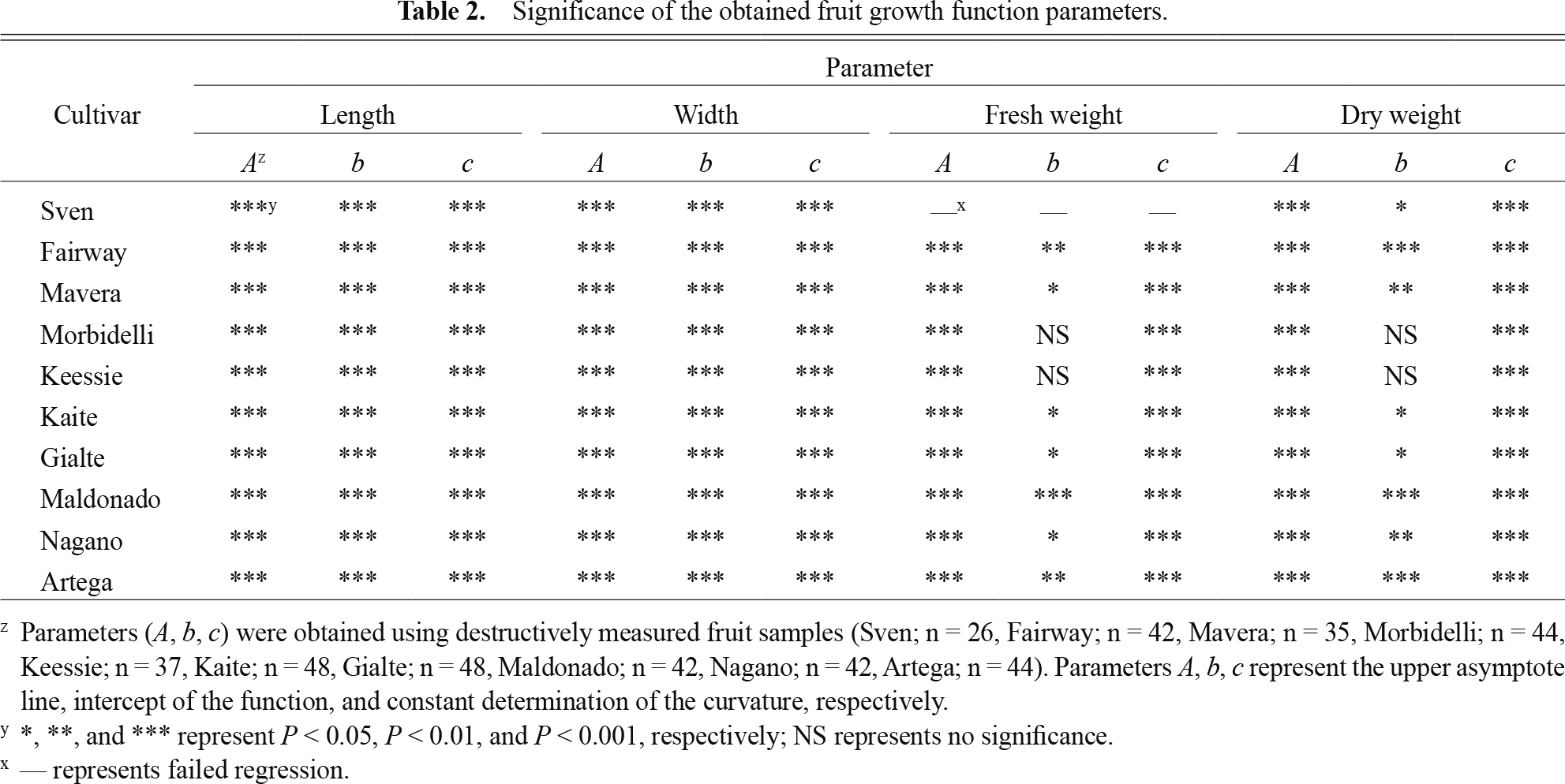
The significances of the obtained Gompertz function parameters are listed in Table 2. Additionally, the obtained parameters are shown in Table S1. The significance of the parameters indicated that fruit growth was appropriately estimated using the Gompertz function and the fruit samples were obtained with sufficient validity. The parameters were significant for almost all cultivars (P < 0.05; Table 2), except for coefficient b for the fruit dry weight and fresh weight of ‘Keessie’ and ‘Morbidelli’ and coefficient A, b, and c for the fruit fresh weight of ‘Sven’. The obtained fruit growth curves are shown in Figures 2 and 3. The fruit length and width at harvest (1,200°C·d days after anthesis [DAA]) were within the range of 80–90 and 70–90 mm per fruit, respectively. The fruit water content at 1,200°C·d DAA was within the range at 200–250 g per fruit, except for ‘Fairway’. The fruit dry weight at 1,200°C·d DAA was within the range at 10–16 g per fruit. Hence, fruit growth varied in terms of volume and weight depending on the cultivar type.
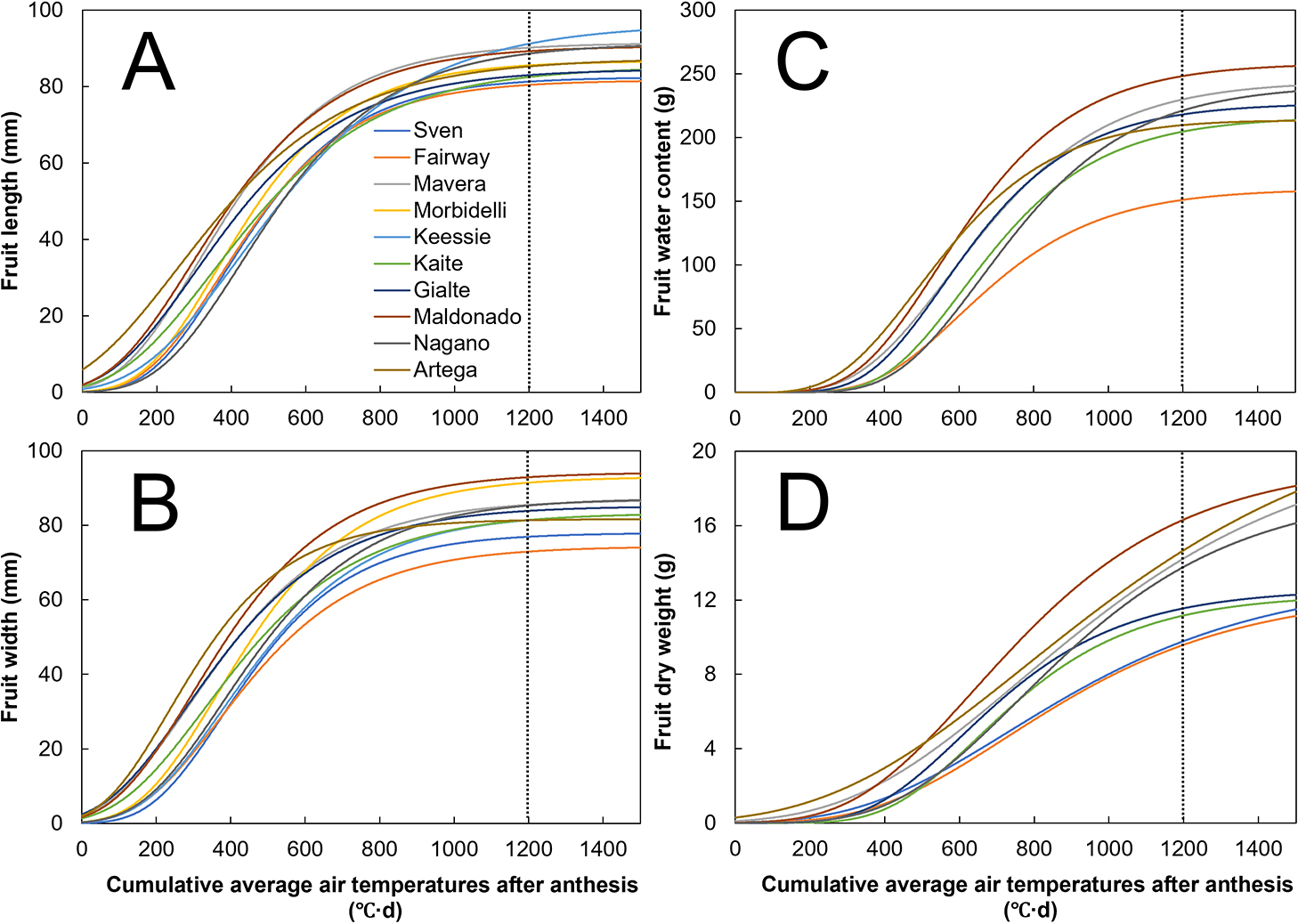
The standardized fruit growth curves for length, width, water content, and dry weight are shown in Figure 4. First, the fruit volume (fruit length: Fig. 4A; fruit width: Fig. 4B) increased to 30–50, 80–90, and 90–100% of its growth potential (i.e., coefficient A in Eq. 1) at 400, 800, and 1,200°C·d DAA, respectively. Second, the fruit water content increased to 5–20, 50–80, and 85–100% of its growth potential at 400, 800, and 1,200°C·d DAA, respectively (Fig. 4C). Finally, the fruit dry weight increased to 5–10, 40–60, and 60–90% of its growth potential at 400, 800, and 1,200°C·d DAA, respectively (Fig. 4D). The growth rate for the fruit dry weight thus showed cultivar-dependent values at 1,200°C·d DAA compared with those for the fruit volume and water content.
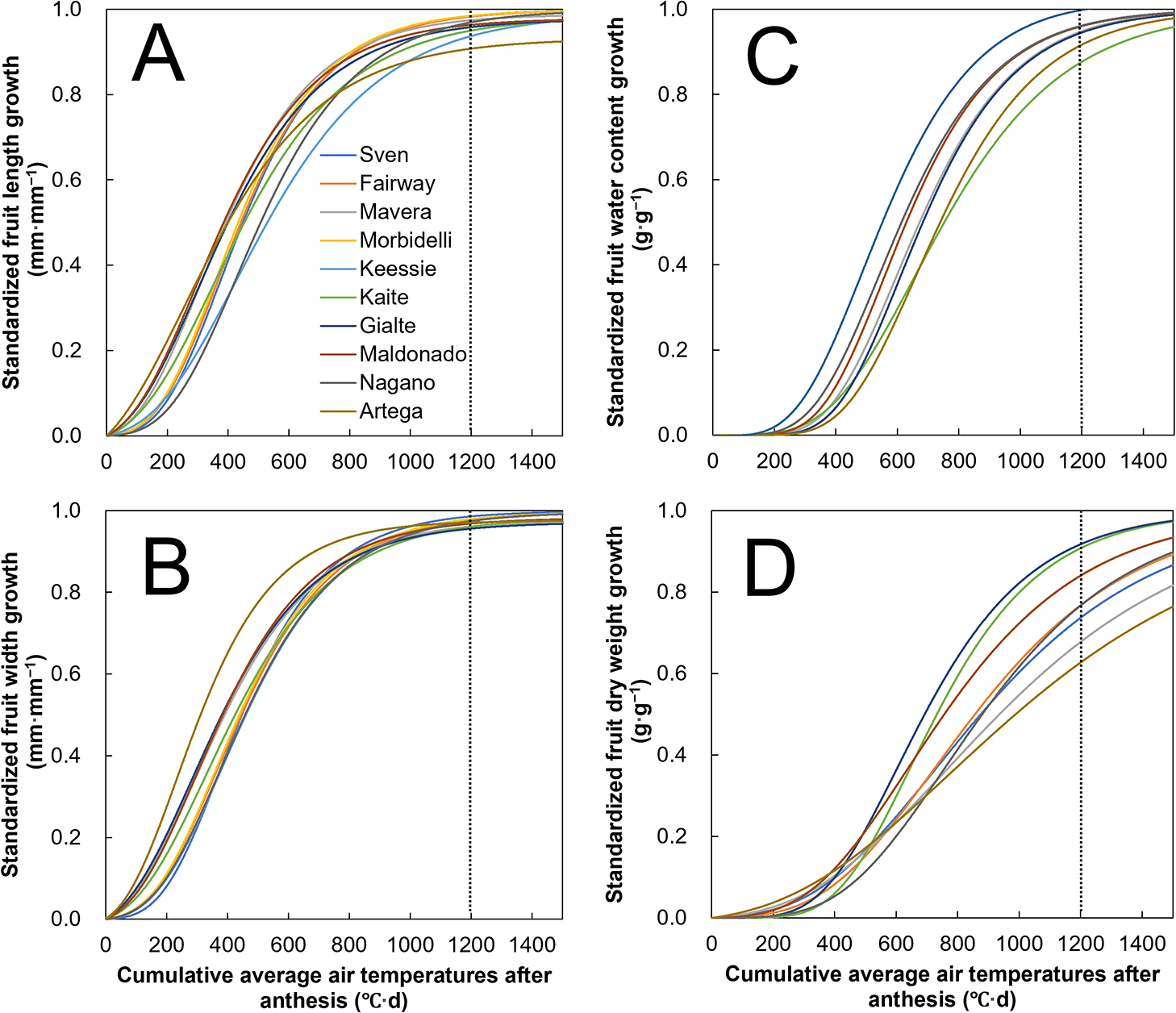
The derivatives of the standardized growth curves for fruit length, width, water content, and dry weight are shown in Figure 5. Totally, rapid growth was observed at 200–600, 400–800, and 600–1,200°C·d DAA for fruit volume (fruit length and width), water content, and dry weight, respectively. In other words, fruit tended to initially increase in volume, followed by an increase in water content and dry weight. Except for fruit dry weight, all other fruit-growth parameters (i.e., fruit length, width, and water content) showed similar growth rates among the cultivars. In particular, dry weight growth curves substantially differed depending on the cultivars at 600–1,200°C·d DAA, whereas the volume and water increase in the same period exhibited only a small difference. For dry weight increase, the convexity of the growth curves was different depending on the cultivar; specifically, that of ‘Gialte’ and ‘Kaite’ had relatively earlier growth periods compared to other cultivars. As a result, fruit dry weight increases in these cultivars mostly occurred during the early growth period.
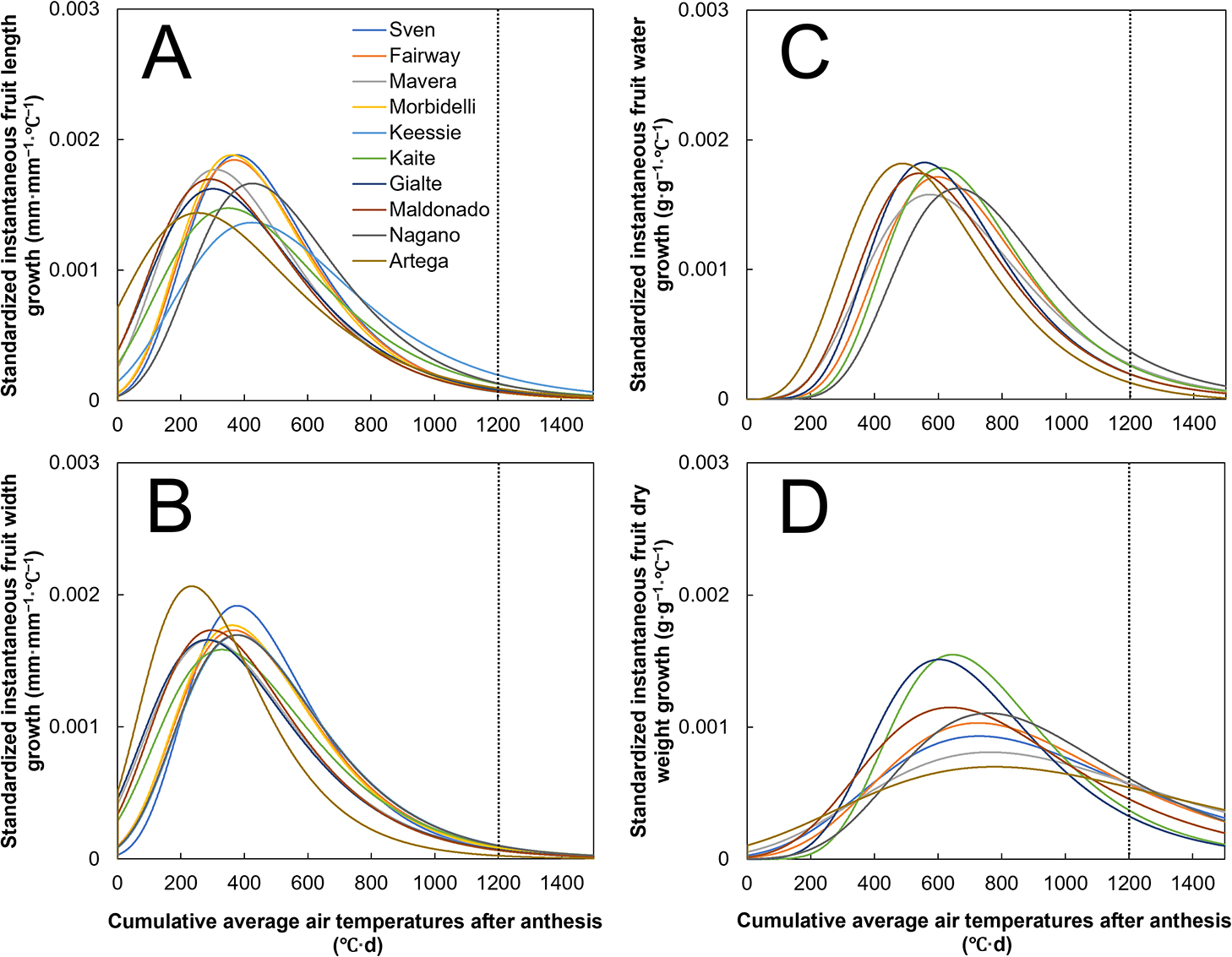
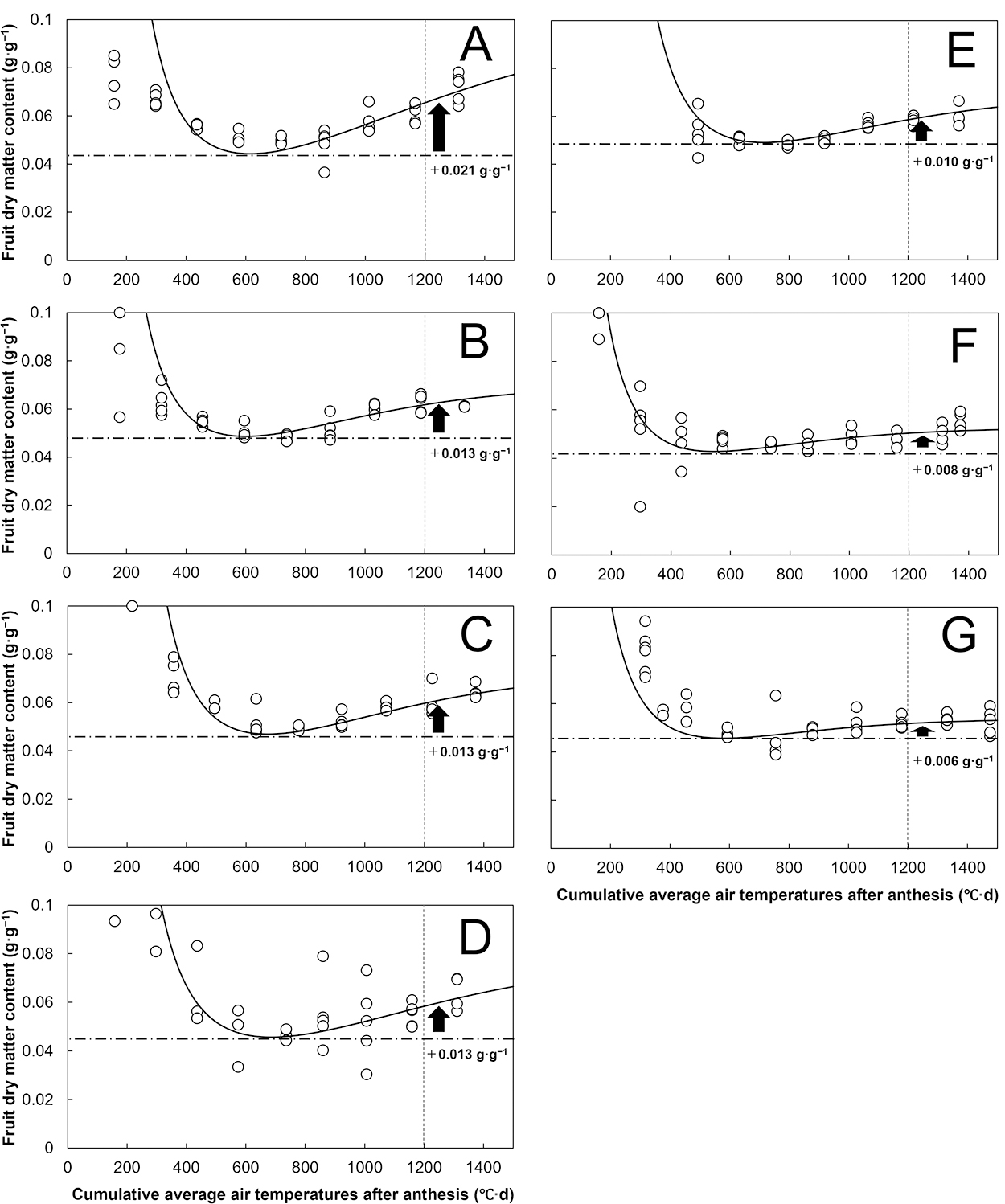
Changes in the estimated and observed fruit dry matter content during fruit growth are shown in Figure 6. The fruit dry matter content decreased to 0.04–0.05 g·g−1 until 500–600°C·d DAA for all the cultivars. From 600–1,200°C·d DAA, however, the fruit dry matter content increased to 0.06–0.08 g·g−1 for ‘Artega’, ‘Maldonado’, ‘Mavera’, ‘Nagano’, and ‘Fairway’ (Fig. 6A, B, C, D, and E), while it hardly increased for ‘Gialte’ and ‘Kaite’ (Fig. 6F, G). From 600–1,200°C·d DAA, ‘Gialte’ and ‘Kaite’, compared with the other cultivars, showed only small increase in dry weight but exhibited a similar increase in water content (Fig. 5C, D). Therefore, the fruit dry matter content of ‘Gialte’ and ‘Kaite’ were maintained at low values (i.e., 0.05–0.06 g·g−1) at 1,200°C·d DAA. In contrast, the cultivars with a high increase in fruit dry weight from 600–1,200°C·d DAA resulted in a high fruit dry matter content at 1,200°C·d DAA (e.g., ‘Artega’; Figs. 5D and 6A). Therefore, the fruit dry matter content rarely increased from 600–1,200°C·d DAA for the cultivars with small dry weight increases (e.g., ‘Gialte’), while it increased for the cultivars with large dry weight increases (e.g., ‘Artega’).
Discussion
Validity of the experimental design
In sweet pepper, previous studies reported that the fruit fresh weight and volume were mainly influenced by the cultivar type, fruit load, and air temperature (e.g., Erickson and Markhart, 2002; Pressman et al., 1998; Rylski, 1986; Wubs et al., 2009). To determine the cultivar-specific growth curves of fruit dry weight and water content, we chose 10 common sweet pepper cultivars with relatively small to large fruit size and wide ranges of fruit dry matter content, partly based on the findings of Homma et al. (2024b). Next, we removed the developing fruits from the plants and allowed only one fruit to grow on each plant to diminish the effects of fruit load on fruit growth (Chen et al., 2024). Moreover, the fruits were grown under appropriate environmental conditions, including a daily average air temperature of 19.4–22.9°C (Fig. 1); the recommended air temperature for sweet pepper growth ranges from 18–25°C (Bakker, 1989; Russo, 2012). Finally, the fruits were harvested when they completely turned red or yellow. Therefore, the experimental conditions were considered suitable for fruit growth, and the obtained data were considered valid for the analyses.
Cultivar-specific fruit dry weight growth could be due to pericarp growth related to fruit sink activity
For ‘Artega’, ‘Maldonado’, ‘Mavera’, ‘Nagano’, and ‘Fairway’, an increase in fruit dry matter content was observed in the later growth stage (600–1,200°C·d DAA) (Fig. 6A, B, C, D, E), which is consistent with previous reports (Marcelis and Baan Hofman-Eijer, 1995; Wubs et al., 2012). However, the yellow cultivars ‘Gialte’ and ‘Kaite’ did not show any marked increase in fruit dry matter content during the same period (Fig. 6F, G). Homma et al. (2024b) reported that ‘Gialte’ and ‘Kaite’ had lower fruit dry matter content than other commercial cultivars and that both cultivars had high fresh yield; additionally, the authors indicated that fresh yield and fruit dry matter content were negatively correlated. In sweet pepper, fruit dry matter content is related to soluble solids within the fruit and Brix content values (Homma et al., 2024b; Rathnayaka et al., 2021). Thus, the unexpected increase in fruit dry weight from 600–1,200°C·d DAA (Fig. 5D) was possibly related to the fruit dry matter content at harvest (Fig. 6), thereby affecting the yield potential and fruit quality (i.e., sweetness).
Sweet pepper fruit is composed of a stalk, seeds, a placenta, and a pericarp (Rylski, 1973). The previous study showed that the stalk, seeds, placenta, and pericarp accounted for about 3–4, 2–3, 8–14, and 80–85% of fruit weight, respectively. We obtained the fruit dry and fresh weight based on whole fruit without the stalk; therefore, the observed fruit weight increase was mainly due to an increase in seeds, the placenta, and pericarp. In addition, Marcelis and Baan Hofman-Eijer (1995) indicated that the dry matter fraction of seeds and placenta within fruit decreased in the later fruit growth stages, while that of the pericarp markedly increased during the same period. In other words, the observed fruit dry matter increases from 600–1,200°C·d DAA could be due to the dry matter increase in the pericarp. Therefore, a cultivar-specific dry weight increase in pericarp from 600–1,200°C·d DAA may result in different fruit dry matter content at harvest.
The reason behind the small increases in the fruit dry weight from 600–1,200°C·d DAA in ‘Gialte’ and ‘Kaite’ was unclear. However, increases in fruit dry weight are related to a plant’s ability to attract assimilates to fruits, and notably this trait is determined by sink activity and sink size (Marcelis, 1996). Sink size is associated with the cell number within fruit cells during floral development; the cell numbers are mainly determined by the assimilate supply to floral organs (Bertin et al., 2002). Photosynthate-limited conditions for fruit growth and anthesis were rare in the present study, except for the cultivar ‘Sven’ especially from 2–5 weeks; in this period, photosynthate supply was possibly limited for this cultivar due to the low solar radiation in winter. Marcelis (1996) suggested that the effect of cell numbers on assimilation attraction is weaker than that of sink activity. Therefore, the differences in the increase of fruit dry weight from 600–1,200°C·d DAA may be attributed not to sink size, but rather to sink activity (Fig. 6).
Fruit sink activity is related to phloem influxes (De Schepper et al., 2013). Additionally, phloem influxes are related to osmotically generated pressure gradients that drive mass flow from leaves to fruits, phloem resistance, and attraction of assimilates to fruits. Initially, dry matter content of leaves and stems in sweet pepper was about 0.13–0.15 and 0.17–0.21 g·g−1, respectively, whereas the fruit dry matter content at harvest was about 0.05–0.10 g·g−1 (Fig. 6; Homma et al., 2023a, 2024b). These findings imply that osmotically generated pressure gradients between the source (i.e., leaves) and sink (i.e., fruit) organs occur constantly within the plant. Second, De Schepper et al. (2013) stated that phloem resistance is mainly determined by the sieve plates within the sieve tube. Third, attraction of assimilates to fruits is related to the seeds within the fruit (Marcelis and Baan Hofman-Eijer, 1997; Yasuor and Wien, 2020). In other words, the fresh weight of individual sweet pepper fruit was positively correlated with seed number or weight within the fruit (Marcelis and Baan Hofman-Eijer, 1997; Yasuor and Wien, 2020). Therefore, the seeds within fruit or sink attractive ability per seed may have triggered the inflows of assimilates towards fruit. Overall, cultivar-specific increases in fruit dry weight from 600–1,200°C·d DAA may be linked to fruit sink activity that is related to phloem influxes, which are in turn determined by the ability to absorb assimilates into fruit and inflow resistance within sieve tubes. Therefore, this ability may determine the cultivar-specific dry matter accumulations within the pericarp, possibly deciding the cultivar-specific fruit dry matter content at harvest.
Possibility to misestimate fruit load due to differences in external and internal fruit growth
Initially, the volume of sweet pepper fruits increased, followed by an increase in water content and dry weight. These findings were partially consistent with those of Marcelis and Baan Hofman-Eijer (1995), indicating that apparent fruit weight (i.e., volume) and actual fruit weight (i.e., water content and dry weight) growths are not similar in sweet pepper. Additionally, whole plant growth is related not to fruit volume, but rather to fruit dry weight growth. For instance, flower abortion and root growth are related to the peaks and valleys of daily fruit dry weight growth (González-Real et al., 2008; Homma et al., 2022). The definition of fruit load is different in previous studies (i.e., fruit number: Fukumoto et al., 2004; Heuvelink, 1997; fruit volume: Koirala et al., 2019; fruit dry weight: Li et al., 2015; Saito et al., 2022). Therefore, crops with different growth rates for exterior fruit volume and interior fruit weight (e.g., sweet pepper) indicate that these different growths may result in a misestimation of fruit load, thereby resulting in a misestimation for whole plant growth. Thus, clarifying the relationship between external fruit volume and internal fruit weight is important to understand whole plant growth, as partly demonstrated herein.
Practical perspective
Our results have some potential applications at production sites. A first application is the option to select plants with a small or large increase in fruit dry weight from 600–1,200°C·d DAA. This selection will produce fruits with a low or high fruit dry matter content at harvest, thereby affecting yield potential and fruit sweetness. Sweet pepper cultivars with high or low fruit dry matter content at harvest are associated with high or low Brix content values, respectively (Rathnayaka et al., 2021). This trade-off is similar to that reported for tomatoes, and market acceptance of these sweet or non-sweet fruits differs depending on regions (Higashide et al., 2012). Therefore, we consider that selecting plants characterized by small or large increase in fruit dry weight from 600–1,200°C·d DAA can lead to selection of high- or low-yield plants (i.e., low- or high-Brix content fruits), in accordance with market preferences.
A second application is the mitigation of flushes (i.e., yield fluctuations characterized by long-term greenhouse sweet pepper production), leading to alleviation of plant exhaustion. The results showed that a cultivar with a high dry weight increase during later fruit growth (e.g., ‘Artega’) will result in a high fruit load during the same period. In sweet pepper, the balance between daily produced dry matter and fruit load (i.e., source-to-sink ratio; Homma et al., 2022) is closely associated with exhaustion of whole plant and flower abortion. Although the cultivar-dependent ability to produce dry matter affects the balance (Homma et al., 2024b; Marcelis et al., 2004), a high fruit load due to a cultivar-specific fruit dry weight increase probably results in plant exhaustion (Saito et al., 2022), additionally associating with flower abortions (Homma et al., 2022). Thus, selecting cultivars with low fruit dry weight during the later growth period (e.g., ‘Gialte’) or selecting cultivars with a high fruit dry weight (e.g., ‘Artega’) with relatively early harvest (i.e., from 1,000–1,200°C·d DAA) may be promising to alleviate plant exhaustion or flushes, respectively. The latter early harvest approach may produce immature fruits; however, Furuno et al. (2019) reported the effectiveness of light irradiation on color promotion for immature harvested fruits. Thus, early harvest accompanied by light irradiance after harvest may be an effective approach for cultivars that require a lot of dry matter in the later fruit growth stage (e.g., ‘Artega’). Further, removing young fruits with a low dry matter increase, but with a high fruit dry matter content (i.e., during 0–600°C·d DAA) may decrease in subsequent fruit load, possibly contributing to alleviate plant exhaustion. This application may therefore be promising to alleviate flush and plant exhaustion.
A third application is the yield prediction using the obtained fruit growth curves. Fluctuating fresh yield of sweet pepper could be precisely predicted using the cultivar-specific fruit growth curves for dry weight (Homma et al., 2023b). This report implies that the cultivar-specific fruit growth curves characterize cultivar-dependent yield changes. Therefore, obtaining a cultivar-specific fruit growth function is necessary for precise yield prediction. We thus consider that leveraging fruit growth curves obtained herein could contribute to precise predictions for yield fluctuations that are related to low productivity (Heuvelink et al., 2002). Overall, the knowledge is potentially and practically appliable at production sites focusing on breeding, alleviating plant exhaustion and flush, and yield prediction.
Limitations
Although the practical implications have been acknowledged, some limitations were also presented in this study. A first limitation is the necessity for investigating cultivars of different colors or sizes. Red and yellow cultivars comprise the majority of sweet pepper production, while orange and other colors account for less than 10% of production (Frank et al., 2001). The present study used 10 common red and yellow cultivars; however, cultivars with small fruit size or different fruit colors were excluded. A second limitation is that the effects of inadequate environmental conditions on fruit growth were not considered. Marcelis et al. (1998) reported that fruit dry matter content changed in response to environmental conditions. Pressman et al. (1998) and Yasuor and Wien (2020) mentioned that sweet pepper plant and fruit growth cease when air temperature drops below 12°C. Additionally, Bae and Kim (2004) and Tadesse et al. (1999) reported that extremely high or low electrical conductivity in nutrient solutions changed fruit dry matter content. Our experiment was conducted under conditions with sufficient water and nutrient supply and an optimal air temperature. In other words, effects of the outside ranges in presented environmental conditions (Fig. 1) on fruit growth are unclear. A third limitation is that this study used air temperature rather than fruit to estimate fruit growth. Air temperature can be considered similar to fruit temperature, but fruit temperature is influenced by not only air temperature, but also other environmental factors such as solar radiation (Helyes et al., 2007). Translocation of assimilates was partly reported to be influenced by fruit temperature changes such as quick temperature drops (Ehara et al., 2017); therefore, we consider that the obtained knowledge can be applicable in production sites for only conditions with similar air and fruit temperatures.
Despite these limitations, this study used relatively small to large, red and yellow bell-shaped cultivar types, which account for the majority of sweet pepper production (Frank et al., 2001). In addition, sweet pepper farmers usually grow plants under optimal conditions to maximize fresh yields, and the experiment has applied these conditions to grow plants. For example, air temperature and electrical conductivity were about 19.4–22.9°C (recommended to 18–25°C; Bakker, 1989; Russo, 2012) and 2.0 dS·m−1 (recommended to 1.5–2.5 dS·m−1; Maas, 1987; Tadesse et al., 1999), respectively; and daily drainage rates were over 30%. Therefore, the results are considered valid under hydroponic production and have expanded knowledge of fruit growth. Additionally, the results may be applicable to breeding or cultivar selection to increase productivity.
Conclusion
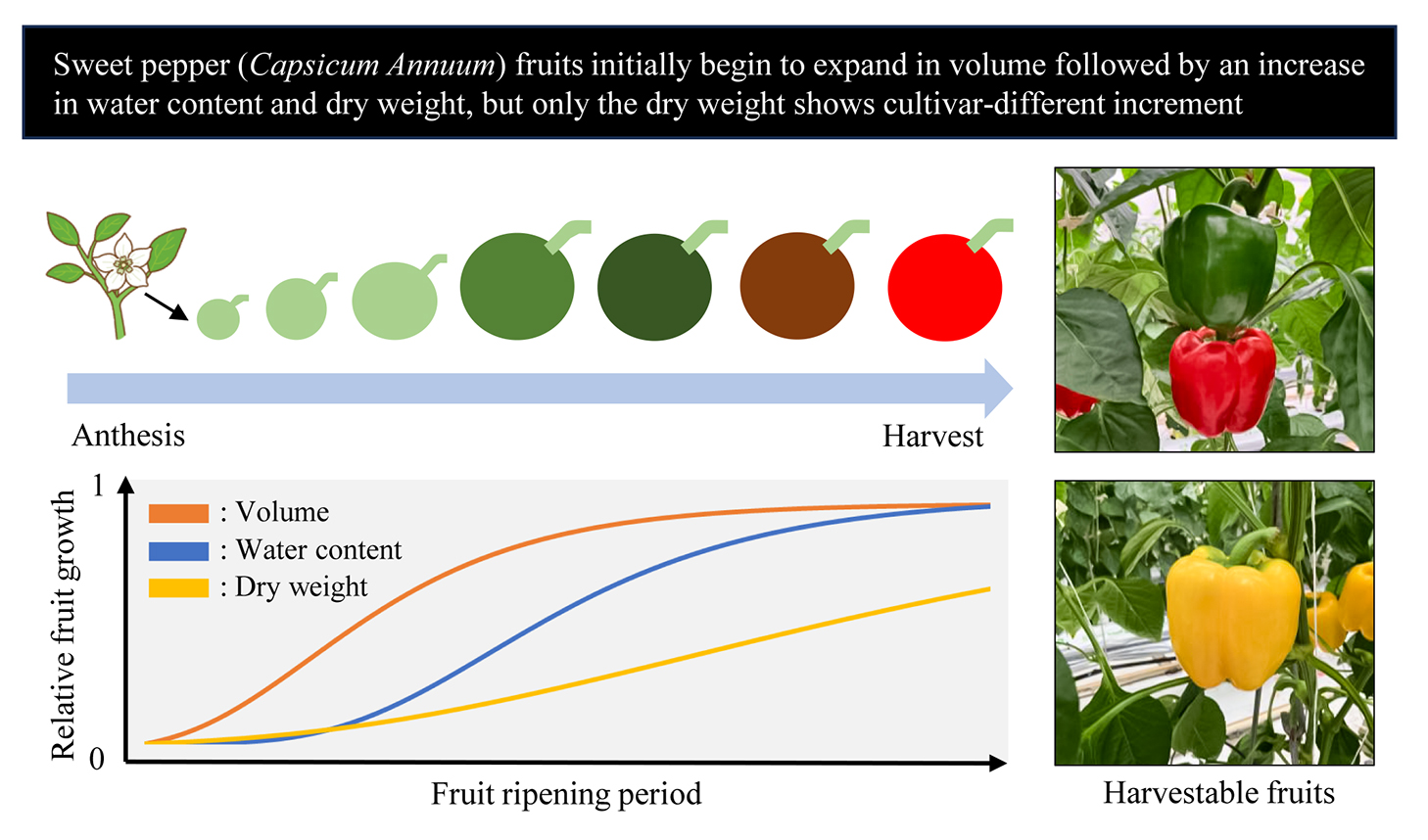
This study showed that (1) sweet pepper fruit initially started to increase in volume and then in water content and dry weight; (2) fruit volume, water content, and dry weight at harvest reached 90–100, 85–100, and 60–90% of their growth potential (i.e., coefficient A in Eq. 1), respectively, indicating that the increases in dry weight showed differences among cultivars; (3) fruit dry matter content decreased to 0.04–0.05 g·g−1 until about 600°C·d DAA, regardless of the cultivar types and thereafter increased from 600–1,200°C·d DAA for the cultivars with high dry weight increase, but hardly increased for cultivars with a low dry weight increase as mentioned in point (2). Basically, interior (i.e., water and dry weight) and exterior (i.e., volume) fruit growth are not consistent in sweet pepper. The fruit dry matter contents at harvest in different cultivars, which is related to yield potential and fruit quality, are affected by cultivar-specific increases in fruit dry weight from 600–1,200°C·d DAA.
Literature Cited
- Adams, S. R., K. E. Cockshull and C. R. J. Cave. 2001. Effect of temperature on the growth and development of tomato fruits. Ann. Bot. 88: 869–877.
- Bae, J. H. and K. H. Kim. 2004. The effect of irrigation concentration on the growth and fruit quality of sweet pepper (Capsicum annuum L.) in fertigation. J. Bio-Env. Con. 13: 167–171 (In Korean with English abstract).
- Bakker, J. C. 1989. The effects of temperature on flowering, fruit set and fruit development of glasshouse sweet pepper (Capsicum annuum L.). J. Hortic. Sci. 64: 313–320.
- Bertin, N., H. Gautier and C. Roche. 2002. Number of cells in tomato fruit depending on fruit position and source-sink balance during plant development. Plant Growth Regul. 36: 105–112.
- Bozokalfa, M. K. and M. Kilic. 2010. Mathematical modeling in the estimation of pepper (Capsicum annuum L.) fruit volume. Chil. J. Agric. Res. 70: 626–632.
- Chen, S., T. Kerstens, B. Zepeda, T. Ouzounis, S. Olschowski, L. F. M. Marcelis and E. Heuvelink. 2024. Additional far-red increases fruit yield of greenhouse sweet pepper mainly through enhancing plant source strength. Sci. Hortic. 338: 113787.
- De Schepper, V., T. De Swaef, I. Bauweraerts and K. Steppe. 2013. Phloem transport: a review of mechanisms and controls. J. Exp. Bot. 64: 4839–4850.
- Ehara, M., A. Tagawa and T. Higashide. 2017. Effects of a higher afternoon temperature followed by a quick temperature drop in the early evening on growth and fruit development in greenhouse cucumber under forcing culture. Hort. Res. (Japan) 16: 155–161 (In Japanese with English abstract).
- Erickson, A. N. and A. H. Markhart. 2002. Flower developmental stage and organ sensitivity of bell pepper (Capsicum annuum L.) to elevated temperature. Plant Cell Environ. 25: 123–130.
- Fukuda, F., Y. Tamaki, T. Kawai, K. Ushijima, K. Hirano, K. Oda, M. Hara, Y. Fukamatsu, K. Morinaga and R. Nakano. 2021. Characterization of fruit enlargement and ripening in extremely late maturing peach. Hort. Res. (Japan) 20: 65–71 (In Japanese with English abstract).
- Fukumoto, Y., Y. Nishimura and K. Shimasaki. 2004. Effects of the fruit load on fruit set and bearing habit in sweet pepper (Capsicum annuum L.). J. Japan. Sci. Hort. Sci. 73: 171–177 (In Japanese with English abstract).
- Frank, C. A., R. G. Nelson, E. H. Simonne, B. K. Behe and A. H. Simonne. 2001. Consumer preferences for color, price, and vitamin C content of bell peppers. HortScience 36: 795–800.
- Furuno, S., C. Yoshida and H. Matsunaga. 2019. Effect of light irradiation after harvest of sweet pepper (Capsicum annuum L.) fruit on yield and color promotion in summer-fall cultivation. Hort. Res. (Japan) 18: 127–132 (In Japanese with English abstract).
- Génard, M. and M. Souty. 1996. Modeling the peach sugar contents in relation to fruit growth. J. Amer. Soc. Hort. Sci. 121: 1122–1131.
- González-Real, M. M., A. Baille and H. Q. Liu. 2008. Influence of fruit load on dry matter and N-distribution in sweet pepper plants. Sci. Hortic. 117: 307–315.
- Hall, A. J. 1977. Assimilate source-sink relationships in Capsicum annuum L. I. The dynamics of growth in fruiting and deflorated plants. Aust. J. Plant Physiol. 4: 623–636.
- Helyes, L., A. Lugasi and Z. Pek. 2007. Effect of natural light on surface temperature and lycopene content of vine ripened tomato fruit. Can. J. Plant. Sci. 87: 927–929.
- Higashide, T. 2022. Review of dry matter production and growth modelling for yield improvement of greenhouse tomatoes. Hort. J. 91: 247–266.
- Higashide, T. and E. Heuvelink. 2009. Physiological and morphological changes over the past 50 years in yield components in tomato. J. Amer. Soc. Hort. Sci. 134: 460–465.
- Higashide, T., K. Yasuba, K. Suzuki, A. Nakano and H. Ohmori. 2012. Yield of Japanese tomato cultivars has been hampered by a breeding focus on flavor. HortScience 47: 1408–1411.
- Heuvelink, E. 1997. Effect of fruit load on dry matter partitioning in tomato. Sci. Hortic. 69: 51–59.
- Heuvelink, E., L. F. M. Marcelis and O. Körner. 2002. How to reduce yield fluctuations in sweet pepper? Acta Hortic. 633: 349–355.
- Ho, L. C., R. I. Grange and A. J. Picken. 1987. An analysis of the accumulation of water and dry matter in tomato fruit. Plant Cell Environ. 10: 157–162.
- Homma, M., D. H. Ahn and T. Higashide. 2024a. Effect of different fruit size and dry matter production on fruit load and fruit set ratio in a greenhouse sweet pepper. Hort. Res. (Japan) 23: 7–20 (In Japanese with English abstract).
- Homma, M., Y. Onodera, M. Kanzaki, S. Kaneko and D. H. Ahn. 2024b. Light-use efficiency and fruit dry matter content affect fresh yield in sweet pepper. HortScience 59: 1378–1385.
- Homma, M., T. Watabe and D. H. Ahn. 2023a. Evaluation of growth characteristics of long-term hydroponically grown sweet pepper based on yield components. Hort. Res. (Japan) 22: 163–172 (In Japanese with English abstract).
- Homma, M., T. Higashide and D. H. Ahn. 2023b. Modeling short-term yield changes in sweet pepper based on dry matter production and fruit growth. J. Amer. Soc. Hort. Sci. 148: 292–303.
- Homma, M., T. Watabe, D. H. Ahn and T. Higashide. 2022. Dry matter production and fruit sink strength affect fruit set ratio of greenhouse sweet pepper. J. Amer. Soc. Hort. Sci. 147: 270–280.
- Hoshi, T., K. Yasuba, H. Kurosaki and T. Okayasu. 2018. Ubiquitous environment control system: An internet-of-things–based decentralized autonomous measurement and control system for a greenhouse environment. p. 107–123. In: S. Hussmann (ed.). Automation in agriculture—Securing food supplies for future generations. InTechOpen Ltd., London, UK.
- Jones, J. W., E. Dayan, L. H. Allen, H. Van Keulen and H. Challa. 1991. A dynamic tomato growth and yield model (TOMGRO). Trans. ASAE 34: 663–672.
- Koirala, A., K. B. Walsh, Z. Wang and C. McCarthy. 2019. Deep learning for real-time fruit detection and orchard fruit load estimation: Benchmarking of ‘MangoYOLO’. Precis. Agr. 20: 1107–1135.
- Li, T., E. Heuvelink and L. F. M. Marcelis. 2015. Quantifying the source–sink balance and carbohydrate content in three tomato cultivars. Front. Plant Sci. 6: 416. DOI: 10.3389/fpls.2015.00416.
- Maas, E. V. 1987. Salt tolerance of plants. p. 57–76. In: B. R. Christie (ed.). CRC handbook of plant science in agriculture. CRC Press, FL, USA.
- Marcelis, L. F. M. 1992. Non-destructive measurements and growth analysis of the cucumber fruit. J. Hortic. Sci. 67: 457–464.
- Marcelis, L. F. M. 1996. Sink strength as a determinant of dry matter partitioning in the whole plant. J. Exp. Bot. 47: 1281–1291.
- Marcelis, L. F. M. and L. R. Baan Hofman-Eijer. 1995. Growth analysis of sweet pepper fruits (Capsicum annuum L.). Acta Hortic. 412: 470–478.
- Marcelis, L. F. M. and L. R. Baan Hofman-Eijer. 1997. Effects of seed number on competition and dominance among fruits in Capsicum annuum L. Ann. Bot. 79: 687–693.
- Marcelis, L. F. M., E. Heuvelink and J. Goudriaan. 1998. Modelling biomass production and yield of horticultural crops: a review. Sci. Hortic. 74: 83–111.
- Marcelis, L. F. M., E. Heuvelink, L. R. Baan Hofman-Eijer, J. Den Bakker and L. B. Xue. 2004. Flower and fruit abortion in sweet pepper in relation to source and sink strength. J. Exp. Bot. 55: 2261–2268.
- Mitchell, P. D. 1986. Pear fruit growth and the use of diameter to estimate fruit volume and weight. HortScience 21: 1003–1005.
- Ngouajio, M., W. Kirk and R. Goldy. 2003. A simple model for rapid and nondestructive estimation of bell pepper fruit volume. HortScience 38: 509–511.
- Pressman, E., H. Moshkovitch, K. Rosenfeld, R. Shaked, B. Gamliel and B. Aloni. 1998. Influence of low night temperatures on sweet pepper flower quality and the effect of repeated pollinations, with viable pollen, on fruit setting. J. Hortic. Sci. Biotechnol. 73: 131–136.
- R Development Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- Rathnayaka, R. M. S. M. B., M. Kobayashi, M. Minami, K. Nemoto and K. Matsushima. 2021. Relationship between the soluble solids metric Brix and other traits in Capsicum peppers. Bull. Shinshu Univ. 19: 55–69.
- Russo, V. M. 2012. Peppers: botany, production and uses. CAB International, Wallingford, UK.
- Rylski, I. 1973. Effect of night temperature on shape and size of sweet pepper (Capsicum annuum L.). J. Amer. Soc. Hort. Sci. 98: 149–152.
- Rylski, I. 1986. Pepper (Capsicum). p. 341–354. In: S. P. Monselise (ed.). Handbook of fruit set and development. CRC Press, FL, USA.
- Saito, Y., T. Ienaka, J. Tezuka and Y. Iwasaki. 2022. Estimation of photosynthesis and fruit load of strawberry. Hort. Res. (Japan) 21: 205–211 (In Japanese with English abstract).
- Tadesse, T., M. A. Nichols and K. J. Fisher. 1999. Nutrient conductivity effects on sweet pepper plants grown using a nutrient film technique: 1. Yield and fruit quality. N. Z. J. Crop. Hortic. Sci. 27: 229–237.
- United States Department of Agriculture (USDA). 2005. Sweet peppers grades and standards. https://www.ams.usda.gov/grades-standards/sweet-peppers-grades-and-standards.
- Vegetable information. 2019. Trends in Korean sweet pepper production, distribution, and exports to Japan (In Japanese). https://vegetable.alic.go.jp/yasaijoho/kaigaijoho/1901_kaigaijoho02.html.
- Watabe, T., Y. Nakano and D. H. Ahn. 2022. The effect of electrical conductivity on fruit growth pattern in hydroponically grown tomatoes. Crops 2: 323–335.
- Wolf, S., J. Rudich, A. Marani and Y. Rekah. 1986. Predicting harvesting date of processing tomatoes by a simulation model. J. Amer. Soc. Hort. Sci. 111: 11–16.
- Wubs, A. M., E. Heuvelink, L. F. M. Marcelis and L. Hemerik. 2007. Survival analysis of flower and fruit abortion in sweet pepper. Acta Hortic. 761: 617–624.
- Wubs, A. M., Y. T. Ma, L. Hemerik and E. Heuvelink. 2009. Fruit set and yield patterns in six Capsicum cultivars. HortScience 44: 1296–1301.
- Wubs, A. M., Y. T. Ma, E. Heuvelink, L. Hemerik and L. F. M. Marcelis. 2012. Model selection for nondestructive quantification of fruit growth in pepper. J. Amer. Soc. Hort. Sci. 137: 71–79.
- Yasuor, H. and H. C. Wien. 2020. Peppers. p. 179–208. In: H. C. Wien (ed.). The physiology of vegetable crops. CABI, Wallingford, UK.