Abstract
In the search for new strains of edible mushrooms in the Brazilian Amazon Forest, we found Lentinula specimens different from Lentinula raphanica. These were described morphologically and evaluated phylogenetically within the Lentinula clade. The mating system was determined, and interbreeding compatibility with L. raphanica was verified. The basidiomata have a cinnamon or deep orange to fulvous brown, moist to dry pileus, occasionally with whitish scales; crowded whitish cream lamellae; and an eccentric to lateral stipe. The typical, predominant basidiospores are 4.4-7.2 µm in length. Endogenous, elongate (7.8-14 µm) basidiospores were also found in some specimens. The long spores seem rare and occasional, but nonetheless a novelty for the group. Basidia are homogeneous in size, cheilocystidia are pyriform or bulboid, and caulocystidia are long and spheropedunculate. The hyphae of the pileipellis are pigment-encrusted. The characteristics match those of Agaricus ixodes originally described from Guyana, currently a synonym of Lentinula boryana. In the phylogenetic trees, such taxon appears distinct from L. boryana and sister to L. raphanica with strong support. This unique lineage was confirmed to be reproductively isolated from sympatric L. raphanica strains. Lentinula ixodes comb. nov. is the second species of the genus reported in the Amazon Forest.
1. Introduction
At least 52 species of edible mushrooms have been documented a from the Amazon Forest in scientific reports, including Lentinula raphanica (Murril) Mata & R. H. Petersen, eaten by indigenous people, such as the Uitoto and Andoke in Colombia (Vasco-Palacios, Suaza, Castaño-Betancur, & Franco-Molano, 2008) and the Yanomami in Brazil (Sanuma et al., 2016). Yanomami from the Awaris region, Roraima State, Brazil, refer to L. raphanica as “Naönaö amo” (Sanöma language), an onomatopoeia of the noise made when eating the mushroom (Sanuma et al., 2016). They cook L. raphanica by boiling it in salted water and serve it with a broth accompanied by “beiju” (dough made from the starch of Manihot esculenta Crantz or cassava) (Sanuma et al., 2016).
The genus Lentinula Earle (Earle, 1909) was typified by Lentinula cubensis (Berk. & M.A. Curtis) Earle ex Pegler [= Lentinula boryana (Berk. & Mont.) Pegler], but remained in Lentinus Fr. (Singer, 1941, 1952, 1955) until Pegler (1975) distinguished the two genera based on the structure of the hyphae. Lentinus boryanus (Berk. & Mont.) Singer and Lentinus edodes (Berk.) Singer (shiitake) were then recombined into Lentinula. Pegler (1983a) proposed three more combinations: Lentinula guarapiensis (Speg.) Pegler, Lentinula novae-zelandiae (Stev.) Pegler and Lentinula lateritia (Berk.) Pegler. Agreeing with Singer (1955), Pegler (1983a) accepted that Agaricus ixodes Mont. and a list of other names/taxa were conspecific (synonyms) with L. boryana. Based on morphological and mating experiments, Guzmán, Salmones, & Tapia (1997) accepted L. boryana (mostly strains from Mexico) as different from L. edodes, but they considered this latter species should include L. novae-zelandiae and L. lateritia as synonyms considering biogeography and sexual intercompatibility. Mata and Petersen (2000) then described Lentinula aciculospora Mata & R.H. Petersen from Central America based on morphological and molecular data. Mata and Petersen (2001) evaluated the morphology of the type specimens of synonymous names under L. boryana sensu Pegler (species complex), and identified Armillaria raphanica Murrill as an independent Lentinula species. Recently, Looney, Buyck, Menolli Jr., Randrianjohany & Hibbett (2021) described L. madagasikarensis Buyck, Randrianjohany & Looney from Madagascar, for a total of eight accepted species in the genus. Of these species, only the edible L. boryana and L. raphanica are reported from Brazil (Montagne, 1849; Berkeley & Curtis, 1869; Pegler, 1997; Capelari, Asai, & Ishikawa, 2010). Together with L. aciculospora in Central America and L. guarapiensis in Paraguay, they represent
the neotropical distribution of the genus. Lentinula boryana was originally described as Agaricus boryanus Berk. & Mont. from the Atlantic Forest in Northeast Brazil (Bahia State) (Montagne, 1849), with other reports from the southeastern and southern regions of the country (see citations listed in Capelari et al., 2010). Lentinula raphanica was confirmed to occur in the Brazilian Amazon Forest (Capelari et al., 2010) based on morphology and molecular (rDNA large subunit [LSU]) data.
After several studies of the phylogeny, biogeography and molecular clock including Lentinula (Hibbett, Fukumasa-Nakai, Tsuneda, & Donoghue, 1995; Hibbett, Hansen, & Donoghue, 1998; Hibbett, 2001; Moncalvo, Lutzoni, Rehner, Johnson, & Vilgalys, 2000; Moncalvo et al., 2002; Wilson & Desjardin, 2005; Matheny et al., 2006; Oliveira, Vargas-Isla, Cabral, Rodrigues, & Ishikawa, 2019; Looney et al., 2021), nine lineages are currently recognized within the genus, the majority of which have been assigned to known species in an American-African clade (L. aciculospora, L. boryana, L. madagasikarensis and L. raphanica) and an Asian-Australasian clade (L. edodes complex - Groups 1 and 5, L. lateritia complex - Groups 2 and 4, and L. novae-zelandiae). With the exception of L. madagasikarensis, these species are currently represented by 28 genome-wide drafts available in the MycoCosm of the Joint Genome Institute - JGI (Grigoriev et al., 2011; 2014), including the genomes of two L. raphanica isolates from the Central Amazon, Brazil.
To date, L. raphanica is the only species of Lentinula reported in the Amazon Forest in Brazil (Capelari et al., 2010; Vargas-Isla, Ishikawa, & Py-Daniel, 2013; Sanuma et al., 2016). In the search for new collections or strains of edible mushrooms in the Brazilian Amazon Forest, we collected Lentinula mushrooms from dried branches of Bertholletia excelsa Humb. & Bonpl. (Brazil nuts, “castanha do Brasil”) that differ from L. raphanica in terms of their deep orange-brown pileus eventually having fine scales. These were morphologically described, and nuclear rDNA internal transcribed spacer (ITS) region sequences were obtained from the collections and included in phylogenetic analyses within the Lentinula clade. The reference strain was assessed through mating experiments with L. raphanica strains.
2. Materials and methods
2.1 Collection sites
Specimens were collected in Jan 2018 and Oct 2021 in the vicinity of Itacoatiara City, on the private property “Fazenda Aruanã” (3°02ʹ21.1ʺ S, 58°471ʹ00.7ʺ W), located on road AM-010 km 215 Manaus-Itacoatiara, Amazonas State, Brazil. The area includes 12,000 ha of B. excelsa agroforestry in the Amazonas River basin. The climate type is Af in the Köppen classification (Alvares et al., 2013). Itacoatiara has 2,830 mm precipitation annually, with a rainy season from Dec to May (297-319-349 mm) and an annual mean temperature of 27.7 °C (ClimaTempo, 2022). The soil is predominantly the clayey yellow latosol (Instituto de Pesquisas e Experimentação Agropecuárias do Norte, 1969).
Three revised specimens deposited at Instituto Nacional de Pesquisas da Amazônia (INPA) herbarium were found to be similar/conspecific with the specimens referred to above. The first, determined as “Lentinus ixodes (Mont.) Singer”, was collected at “Empresa Brasileira de Pesquisa Agropecuária” (EMBRAPA) in June 1977 at km 30 of the same road, near the protected area “Reserva Florestal Adolpho Ducke” (RFAD), Manaus, Amazonas, Brazil. The second, determined as L. raphanica, was collected in Feb 2014 in the RFAD. The third, another herbarium specimen determined as Lentinula cf. boryana, was collected in the “Reserva de Desenvolvimento Sustentável do Uatumã” (RDSU) - ATTO project, São Sebastião do Uatumã City, Amazonas, Brazil. The RFAD is located north of Manaus City (02°55ʹ S 59°59ʹ W) in a 10,000 ha area of mostly non-flooded (terra firme) forest (Prance, 1978) with annual rainfall of 1,975 mm (± 332), rainy season from Nov to Jun, and mean temperature of about 26 °C (Braga-Neto, Luizão, Magnusson, Zuquim, & Castilho, 2008). The RDSU is located 150 km northeast of Manaus City and consists of an area of 424,430 ha composed of a forest mosaic, with an annual rainfall of ~2,040 mm and mean temperature of 27.4 °C (Resende et al. 2020).
The sites where the collections were obtained are shown in maps (most approximated localizations; Supplementary Material, Figs. S1 and S2).
2.2 Morphological description
Specimens were photographed in the field and macroscopically described based on fresh material. Color-coding is from Küppers (2002). The fresh specimens were dried at 40 °C. Thin sections of dried basidiomata treated in 70% ethanol were mounted in 5% (W/V) KOH or in Melzer's reagent, and observed using a DM 2500 optical microscope (Leica, Wetzlar, Germany). Measurements were taken from 30 basidiospores, 15 basidia/basidioles/cystidia and 10 hyphae of trama per category. Basidiospore measurements provided the min-max ranges of length and width, where xrm is the min-max range of the arithmetic means of length and width; xmm is the arithmetic means of length (± standard deviation, SD) and width (± SD); Qrm is the min-max range of the means of length/width; Qmm is the mean of the means of length/width (± SD); n is the number of basidiospores measured in a collection; and s is the number of different collections examined. The shape of the basidiospores was determined from the Q values. Line drawings of the microstructures were made using a drawing tube attached to a microscope. The collections were deposited at the INPA Herbarium.
2.3 Sequencing
This study was registered in SisGen (Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado) under the number A1A1061. Two ITS sequences of the reference strain (INK-04) were obtained in two different laboratories to improve resolution and minimize sequencing noise and a third ITS was obtained from RVI29 to avoid equivocal amplification of paralogues. DNA sequences of the ITS region (ITS1-5.8S-ITS2) were obtained according to the method of Oliveira et al. (2019) with the primers ITS1/ITS4 and ITS1F/ITS4. PCR was performed with an initial denaturation step at 95 °C for 2 min followed by 34 cycles of denaturation for 45 s at 94 °C, annealing for 10 min at 50 °C, and extension for 2 min at 72 °C, with a final extension step of 10 min at 72 °C followed by cooling to 4 °C.
2.4 Phylogenetic analyses
The sequences were assembled with Geneious R7 software and deposited in GenBank (NCBI). The produced ITS sequences were included in Dataset 2 of Looney et al. (2021). The outgroup was replaced with data of two Rhodocollybia species. The consolidated dataset was aligned using MUSCLE v3.8.31 (Edgar, 2004), and the alignment was inspected and manually edited in Geneious R7. Poorly or ambiguously aligned sites were removed from the alignment and the ends were trimmed. The HKY+I+G nucleotide substitution model was selected based on the Akaike information criterion calculated using MrModeltest 2.3 (Nylander, 2004).
Maximum likelihood analysis (ML) and Bayesian analysis (BA) were performed in RAxML 7.0.4 (Stamatakis, 2006) and with MrBayes 3.2.1 (Ronquist et al., 2012), respectively. Through ML analysisn, trees were reconstructed in GTRGAMMA with 1,000 rapid bootstrap inferences, to identify the best scoring tree. GAMMA model parameters were estimated to an accuracy of 0.001 log likelihood units. MC3 BA analysis consisted of two independent runs of 5,000,000 generations, sampled every 500 generations, with six independent chains and two swaps. The burn-in was 10%. The final tree was obtained through the 50% majority-rule consensus method. Phylogenetic trees were visualized in FigTree 1.3.1 and edited in CorelDRAW X7. Values supporting for the tree nodes included the bootstrap (BS) percentage (strongly supported, ≥ 80; moderately supported, ≥ 70) and posterior probability (PP) (strongly supported, ≥ 0.98; moderately supported, ≥ 0.95). All sequences from GenBank used in these analyses are identified by their accession number in Supplementary Material, Table S1.
2.5 Mating type and crossing
Monosporic isolates were obtained from the fresh basidiomata of strains INK-04 (L. ixodes), INK-05 and INPACM1701 (L. raphanica). Basidiomata of INK-05 were collected from the same site, on the same date, as INK-04. The pileus without the stipe was placed top-down in a sterilized Petri dish (Miles, Takemaru, & Kimura, 1966) and covered with a funnel at 25 °C for 4-12 h to discharge basidiospores. Basidiospores obtained from spore prints were resuspended in sterile distilled water. Next, one aliquot of this suspension was spread over potato dextrose agar (PDA) in Petri dishes (Vargas-Isla et al., 2015). Inoculated media were incubated at 25 °C for 24-72 h.
Individual germinated basidiospores were detected using an optical microscope, lifted out, and transferred to fresh PDA Petri dishes using an entomological needle with a supporting handle (Vargas-Isla et al., 2015). The germinated basidiospores were isolated by first locating those that were sufficiently separated from each other, making cuts on each side and lifting out a small PDA plug containing the germinated basidiospore (Miles et al., 1966). The germinated basidiospores were then observed under a microscope and transferred to PDA in individual Petri dishes. After five days at 25 °C, the haploid isolates developed into a mycelia that were sufficiently large to be used for mating type analysis (Miles et al., 1966); the absence of clamp connections was subsequently verified.
Mating types were determined by pairing the monosporic isolates in all possible combinations (Vargas-Isla et al., 2015). The paired cultures were incubated at 25 °C for 72-192 h and observed under a microscope to assess the formation of clamp connections. In a methodology adapted from Miles et al. (1996), plugs (2 × 2 mm) from each monosporic isolate were placed at a distance of 1-2 mm from each other to determine mating compatibility. Plug pairings were setup as four per PDA plate in all possible combinations (i.e. plate A: 1 vs. 1, 1 vs. 2, 1 vs. 3, 1 vs. 4, plate B: ...). Cross experiments consisted of INK-04 (L. ixodes) × INK-05 (L. raphanica), INK-04 (L. ixodes) × INPACM1701 (L. raphanica), and INK-05 (L. raphanica) × INPACM1701 (L. raphanica). Self-crossing was also tested for these three strains. Plates were incubated at 25 °C for 24-72 h and observed under a microscope.
3. Results
3.1 Taxonomy
Lentinula ixodes (Mont.) J.S. Oliveira, T.S. Cabral, R. Vargas-Isla, & N.K. Ishikawa, comb. nov. Fig. 1, 2, 3, 4, 5.
MycoBank no.: MB 842599.

Fig. 1 - Basidiomata of Lentinula ixodes. A, B: INK-04 (INPA 289637!). C-F: JO1637 (INPA 289982!). G-H: JO1643 (INPA 289987!). Bars: 10 mm.

Fig. 2 - Illustrations of microstructures of Lentinula ixodes (INK-04, INPA 289637!). A1: Predominant basidiospores; A2: rare basidiospores. B: Basidia. C: Basidioles. D: cheilocystidia. E: Hyphae incrusted wall outgrows in the pileus trama. F: Caulocystidia. Bars: 10 μm.
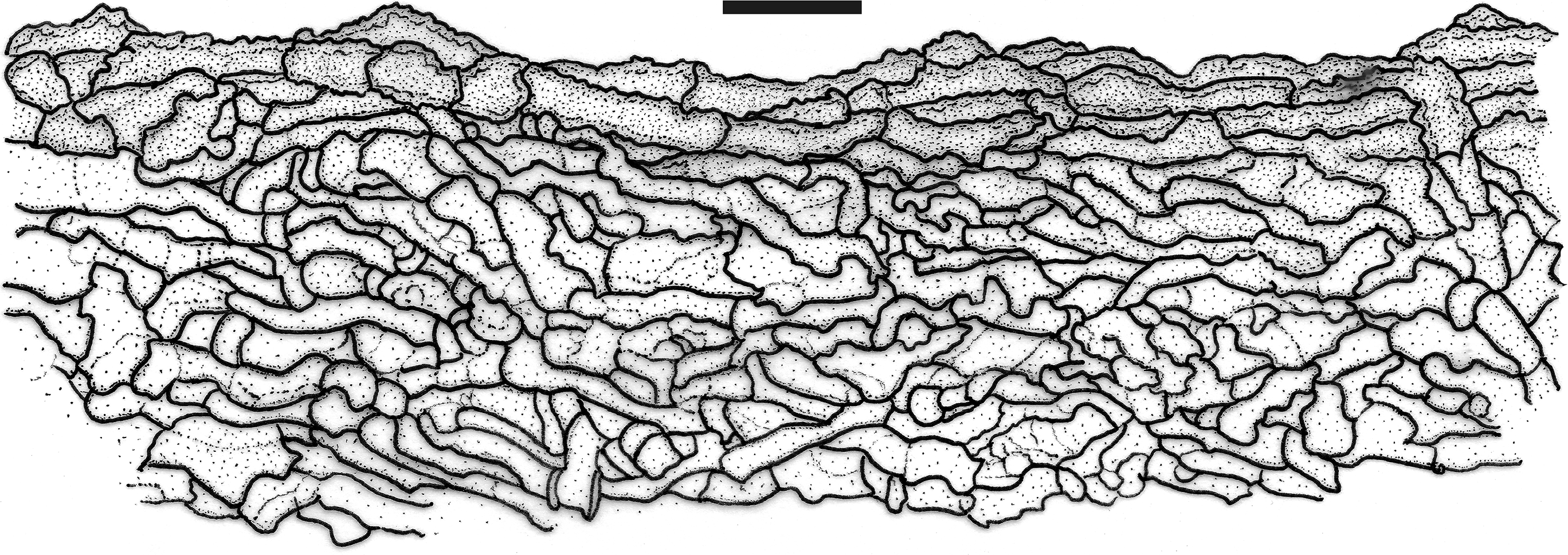
Fig. 3 - Profile section of the pileus (20×) of Lentinula ixodes (INK-04, INPA 289637!). Bar: 50 μm.

Fig. 4 - Color photographs of microstructures of Lentinula ixodes (INK-04, INPA 289637!). A: Predominant basidiospores. B: Basidium (indicated by a red arrow) and basidioles. C: Lamellae trama and hymenium. D: Cheilocystidia (indicated by red arrows). E: Pileipellis and pileus trama. F: Thick-walled hyphae of the pileus trama. F: Stipitipellis. G: Caulocystidia.

Fig. 5 - Color photographs of the rare long spores of Lentinula ixodes (A-G from INK-04, INPA 289637!; H from JO1637, INPA 289982!). A, B: Hymenium with long spores (indicated by red arrows) still attached. C-F: Long spores (indicated by a red arrow) still attached to basidia. G: A single long (indicated by a red arrow) spore among the predominant spore forms. H: Long spores (indicated by a red arrow) form attached to a basidium.
Basionym: Agaricus ixodes Mont., Annls Sci. Nat., Bot., sér. 4 1: 95. 1854.
≡ Collybia ixodes (Mont.) Sacc., Syll. fung. (Abellini) 5: 232. 1887.
Type: Guyana, on dead trunk, Leprieur 982 (K, and ex Patouillard 4055 in FH).
Pileus 8-28 mm diam (Fig. 1), when immature and dried about 5 mm diam (R. Singer & I. Araújo 10008), orbicular to hemicircular, then reniform or dimidiate, convex or planoconvex, often truncate, with an inconspicuous shallow umbo on a wide flat disc, smooth, margin decurved, often forming abrupt declivities causing zonation (at the mid-disc and margin), initially involute, edge entire to slightly crenate; surface glabrous, with rare, small, and whitish orange squamules (thin remnant tissues) at the mid-disc/margin, dry, less often appearing viscid when soaked, often ceraceous, sometimes radially fibrillose and often forming line marks, hygrophanous at some points, slightly fleshy at the disc to coriaceous or membranous at the margin, context white to cream (up to 2.5 mm), vivid orangish or fulvous brown (M40Y80-99M50-70), or ferruginous brown (M60Y80-99M60-70) on the disc and flat area, concolorous or pale beige orange (N20Y60M40) at the margin, edge orangish cream (N00Y20-30M10 to N10Y50-60M20-40), darker tan brown (N50Y90M50) at the threshold between margin and disc, sometimes with a distinctly deep ochraceous or cinnamon brown (N70Y99M60-70) center becoming paler cinnamon from the mid-disc to margin (N20Y50-60M30, N30Y70-80M40) and occasional dark fulvous brown (N90Y40-50M60) spots in random positions, chocolate brown (N70Y90-99M60-80) when dried. Lamellae (Fig. 1) free and near or moderately distant from the stipe apex (a downy area), seldom nearly adnexed or slightly sinuate into the furfuraceous stipe apex, never toothed or forming a pseudocollarium, very close, crowded, lamellae number ≅ 40-50 or more, very narrow, equal, regular, lamellulae series ≥ 3, white to cream (N00Y10M00), edge entire, non-marginate. Stipe 6-13 × 1-2.2 mm (Fig. 1), about 5 × 1.5-2 mm when immature and dried (R. Singer & I. Araújo 10008), mostly eccentric (nearly but not lateral), seldom central, cylindrical with circular caliber, equal or slightly broadened at the apex or toward the base, never bulbous, sometimes surrounded by a mycelial pad, fibrous, tough, medulla solid or spongy, surface opaque, slightly striate, more furfuraceous at the apex, becoming glabrous in the middle toward the base, generally paler than the pileus, orangish yellow (N20Y70M40) or mostly dirty white (N10Y20-30M10) covered with a downy whitish pellicle. Veil or annulus evanescent during maturation,
completely absent in mature basidiomata but with a cottony cream-colored partial veil or annulus observed on dried primordia (R. Singer & I. Araújo 10008). Odor mild. Flavor not recorded.
Basidiospores (Figs. 2A1, 4A) 4.4-7.2(-7.7) × 2-4 µm [xrm = 5.6-6.4 × 2.6-3.1 µm, xmm = 6 (± 0.3) × 2.8 (± 0.1) µm, Qrm = 1.3-3, Qmm = 2.2 (± 0.1), n = 30, s = 10], obovoid or shortly oblong, ellipsoid to subellipsoid, or lacrymoid to shortly clavate; rare and/or occasional, elongate forms (Figs. 2A2, 5) shared by INK-04, R. Singer & I. Araújo 10008, JO1637 and JO1639 (4 of 10 collections), 7.8-14 × 2-4.3 µm, fewer 16.8-21.8 µm long, oblong, clavate to subfusoid, both smooth, hyaline, thin-walled, inamyloid. Basidia (Figs. 2B, 4B, 5) 13-20.4 × 4-5 µm, cylindrical to clavate, smooth, hyaline, thin-walled, 2-4-sterigmate, inamyloid. Basidioles (Fig. 2C) (13.7-) 14.5-19 × 2.8-5 µm, cylindrical to clavate, some with a more inflated apex, smooth, hyaline, thin-walled, inamyloid. Pleurocystidia absent. Cheilocystidia (Figs. 2D, 4D) 14.7-33 × 6.6-12.7 µm, present in intermittent clusters at the sterile lamellar edge in the form of smooth cells (Globulares-type), broadly clavate, mostly pyriform or bulboid, regularly shaped, sometimes slightly pedicellate, smooth, hyaline, thin-walled to slightly thick-walled. Lamellar trama inamyloid, subregular, slightly gelatinized, hyphae 1.2-6 µm diam, cylindrical, regular in outline, occasionally branched, smooth to very slightly incrusted with excrescences of external wall surface, hyaline, thin- to moderately thick-walled. Pileus trama (Figs. 3, 4E) inamyloid, irregular, non-gelatinous, hyphae strongly interwoven, 2.3-10.5 µm diam, cylindrical, regular in outline, straight to wavy, or serpentine-contorted, regularly branched, hyaline to slightly fuscous, becoming pale yellowish brown near the pileipellis, smooth or with irregular (wavy to nodose incrustation) walls (Figs. 2E, 4F) often thicker (0.7-3.6 µm) than those of the lamellar trama, some thin-walled. Pileipellis (Figs. 3, 4E) a non-gelatinized cutis of disorganized, interwoven, non- or somewhat radially oriented hyphae, consisting of an upper stratum of pileus trama, brown to ochraceous brown in KOH, hyphae pale yellowish to brownish when observed single, 3.5-10.2 μm diam, broad like those of the pileus trama, presenting short terminal endings, sometimes cellular, obtuse and rounded, bulboid or pyriform, thick-walled, smooth or with irregular external walls, rough or slightly pigment-incrusted, inamyloid.
Stipe trama inamyloid, cortical and internal hyphae tending to parallel but interwoven, hyphae cylindrical, 1.6-8 µm diam, regular in outline, smooth, hyaline to slightly fuscous, thin- to very thick-walled. Stipitipellis a pellicle of very interwoven, prostrate, loose, slender hyphae, 2-3 µm diam., cylindrical, regular to wavy or curve-contorted, smooth, hyaline, inamyloid. Caulocystidia (Figs. 2F, 4F-G) more frequent at the stipe apex, grouped in irregular and inconspicuous fascicles, 28.4-63.7 × 5.2-9 μm, long clavate to spheropedunculate, or filamentous with broad and bulboid apex, or irregular in outline, sometimes forming sprouts, or contorted, smooth, hyaline, moderately thick-walled, inamyloid. Clamp connections present in pileus, lamellar and stipe trama and structures examined, conspicuous.
Habit and habitat. Gymnopoid (Fig. 1), gregarious, on dead wood (branch or trunk) of B. excelsa forestry, previously terra firme forest.
Examined material: BRAZIL, Amazonas State, Manaus City, road AM-010 km 30 Manaus-Itacoatiara, EMBRAPA, 23 Jun 1977, R. Singer & I. Araújo 10008 (INPA 77432!, as Lentinus ixodes), Reserva Florestal Adolpho Ducke, on wood, 11 Feb 2014, N.K. Ishikawa 2846 (INPA 272271!); Itacoatiara City, Fazenda Aruanã, P14, forestry (B. excelsa), 16 Jan 2018, N.K. Ishikawa, F. Schaedler & F. Bruno INK-04 (INPA 289637!), forestry (B. excelsa), 25 Oct 2021, J.J.S. Oliveira, N.K. Ishikawa, R. Vargas-Isla, & S.M. Benzecry JO1637 (INPA 289982!), J.J.S. Oliveira et al. JO1638 (INPA 289983!), J.J.S. Oliveira et al. JO1639 (INPA 289984!), J.J.S. Oliveira et al. JO1642 (INPA 289986!), J.J.S. Oliveira et al. JO1643 (INPA 289987!), R. Vargas-Isla, N.K. Ishikawa, J.J.S. Oliveira, & S.M. Benzecry RVI29 (INPA 290672!); São Sebastião do Uatumã City, Reserva de Desenvolvimento Sustentável do Uatumã, terra-firme forest, 80 m near the tower, 27 Feb 2019, D.L. Komura, R.P. Alves & C.G.G. Barbosa 2455 (INPA 285163!).
DNA sequences: OL329824 (ITS) and OL329825 (ITS) from INK-04; ON117629 (ITS) from RVI29.
Comment: Lentinula ixodes differs from L. boryana and L. guarapiensis in the larger basidiospores [4.4-7.2(-7.7) × 2-4 μm vs. 4.8-6.4 × 2.4-3.2 μm and 4-4.5 × 2.3-3 μm, respectively] and deeply pigment-incrusted, thick-walled hyphae forming the pileipellis. The caulocystidia (28.4-63.7 × 5.2-9 μm) are longer than those of L. boryana (12-44 × 3-8 μm), while the presence of cheilocystidia distinguishes it from L. guarapiensis that has none. Lentinula boryana has a whitish pileus (type), while L. ixodes has a deeply orange-brown pileus. Two sizes of spores are only found in L. ixodes. More details are provided in Supplementary Material, Table S2.
3.2 Phylogeny
The ITS ML and BA trees (Fig. 6) present the same consistent topology. The ingroup, Lentinula clade (BS 100/ PP 1.0), is delimited and rooted on Rhodocollybia species. This clade formed two mostly unsupported major subclades. The subclade at the bottom of both trees grouped L. boryana, L. ixodes and L. raphanica as monophyletic but without support. Lentinula boryana is basal to L. ixodes and L. raphanica, which are sister with high support (BS 100 / PP 1.0). The major subclade at the top is weakly supported (BS 70) and bears a basal small clade of L. aciculospora and L. madagasikarensis as sister with high support (BS 95 / PP 1.0) and a strongly supported small clade (BS 99 / PP 1.0) of five species-level lineages assigned to L. edodes, L. lateritia and L. novaezelandiae. Lentinula edodes and L. lateritia are each represented by two distinct lineages. Overall, 10 species-level lineages are singular and highly supported in the Lentinula clade.

Fig. 6 - Phylogeny of Lentinula including Lentinula ixodes comb. nov. based on ITS dataset inferred by ML and BA analyses. Numbers on nodes are statistical support values of bootstrap and posterior probability, respectively. Bold black stems are strongly supported and bold gray are moderately supported.
For L. ixodes (INK-04), 12 haploid isolates were used to define the tetrapolar system (Fig. 7). For L. raphanica, tetrapolarity was defined with seven haploid isolates of INPACM1701 (Fig. 8) and six of INK-05 (Fig. 9). Mating tests between L. ixodes and L. raphanica, using each recognized type, were negative for all pairings (INK-04 × INK-05 and INK-04 × INPACM1701). Intersterility was confirmed based on the absence of clamp connections in the contact region (growing edge) between paired strains and the distal sides of the inoculum plugs (Figs. 10, 11).

Fig. 7 - Haploid isolates of Lentinula ixodes comb. nov. (INK-04, INPA 289637!). Self-cross grid among twelve haploid isolates on PDA medium at 25 °C for 72-192 hours. (+) compatible pairing; (-) incompatible pairing. Haploid isolates number in bold were selected for the crosses of tested strains.

Fig. 8 - Haploid isolates of Lentinula raphanica (INPACM1701). Self-cross grid among seven haploid isolates on PDA medium at 25 °C for 72 hours. (+) compatible pairing; (-) incompatible pairing. Haploid isolates number in bold were selected for the crosses of tested strains.

Fig. 9 - Haploid isolates of Lentinula raphanica (INK-05). Self-cross grid among six haploid isolates on PDA medium at 25 °C for 72 hours. (+) compatible pairing; (-) incompatible pairing; (?) pseudo-clamp. Haploid isolates number in bold were selected for the crosses of tested strains.

Fig. 10 - Crosses of tested strains between Lentinula ixodes comb. nov. (INK-04, INPA289637!) × L. raphanica (INPACM1701and INK-05). Haploid isolates on PDA medium at 25 °C for 24-72 hours. (-) incompatible pairing.

Fig. 11 - Crosses between Lentinula raphanica (INPACM1701 and INK-05). Haploid isolates on PDA medium at 25°C for 72 hours. (+) compatible pairing; (-) incompatible pairing.
4. Discussion
Four of the ten collections of L. ixodes mentioned above are the only ones reported to date with two ranges of basidiospore sizes, with the larger being the longest in the whole genus. The elongate basidiospores still attached to basidia (Fig. 5) unequivocally indicated that they are endogenous. However, they are rare compared with the shorter basidiospores (Figs. 4A and 5F) and can be easily overlooked even by an experienced taxonomist. Basidia are homogeneously sized for both spore forms. Coordinated basidiospore sampling was conducted and the details presented in the Supplementary Material. In addition to being unexpected for the group and quite rare, they were not observed in all collections (even in different basidiomata of a single collection) of L. ixodes, and the sizes were highly variable with no clear pattern unlike those of small size. These observations suggest that this is an occasional and rare characteristic with low taxonomic significance. This does not minimize the relevance of the finding, the phenomenon itself, or the need for further study of the cause and underlying mechanisms. With the available data, we cannot consider this feature sufficient to distinguish it from A. ixodes, especially given the high correspondence of other features (Supplementary Material, Table S2). This spore dimorphism is described for the first time for Lentinula. Dimorphic basidiospores are more common in Hygrocybe species, some of which have macrospores and microspores (Pegler, 1983b, 1986; Singer, 1989; Lodge, et al., 2014). A few species of marasmioid and gymnopoid fungi from Malaysia with two spore sizes are more closely related (Corner, 1996): Marasmius gracilichorda Corner (Marasmiaceae), Marasmius indoctus Corner [≡ Collybiopsis indocta (Corner) R.H. Petersen, Omphalotaceae], and Marasmius naucoriiformis Corner (possibly Omphalotaceae in the current classification).
The specimens analyzed fit within the L. boryana complex, which is a broad morphology-based concept including taxa embedded in L. boryana sensu Pegler (1983a) and is accepted by Mata and Petersen (2001). The type specimen of L. boryana has basidiospores with compatible dimensions, of 5-6 × 2-3.5 μm (Pegler, 1983a) and 4.8-6.4 × 2.4-3.2 μm (Mata & Petersen, 2001), although they are slightly shorter than the usual (small) basidiospores of the material examined here [4.4-7.2(-7.7) × 2-4 µm]. Lentinula boryana also has smaller (14-21 × 4-10 μm) and more clavate cheilocystidia, smaller (12-44 × 3-8 μm) clavate to subventricose caulocystidia (Mata & Petersen, 2001), and a whitish pileus (Montagne, 1849). Pegler (1983a) accepted the merging of L. boryana with nine synonyms and wide variation of pileus pigmentation: “ivory white, discolouring with age, in streaks or patches to yellowish brown (M.8YR/5.9/9.5) or vinaceous rufous (M.10R/4.7/8.5)”. Mata and Petersen (2001) provided a type revision of L. boryana and its synonyms (Pegler's concept), with more detailed descriptions of each type material. Within the L. boryana complex, the specimens described here are morphologically quite compatible to A. ixodes (= L. boryana) based on the type described by Mata and Petersen (2001). Mata and Petersen (2001) did not observe caulocystidia, possibly due to the preservation conditions. For further comparison of morphological characteristics between the examined collections and a stricter species concept of L. boryana (solely the type specimen), see the Supplementary Material, Table S2.
Based on data from Montagne (1854), Singer (1955), Mata and Petersen (2001), and this study, the specimens evaluated here are A. ixodes, based mainly on the overall compatible macromorphology of the basidiomata, basidiospore size, cheilocystidia size and shape, complexity of the pileus trama, and pileipellis structure (Supplementary Material, Table S2). They only diverge from the protologue in not having a glutinous pileus. In the first revision, Singer (1955) examined the type specimen of A. ixodes from both K and FH herbaria; the characteristics match the specimens analyzed here, differing only in the smooth, thin-walled hyphae forming the pileipellis. Singer also observed that it was definitely non-gelatinized and concluded that the pileus surface could not have been glutinous even in youth stage of the basidiomata. However, he finally considered A. ixodes to be a synonym of Lentinus boryanus (Berk. & Mont.) Singer (current name Lentinula boryana), and this interpretation was accepted by Pegler (1983a). Agaricus ixodes was originally described from Guyana (Montagne, 1854), which also includes part of the Amazon Forest at the extreme north of South America. Along with the singularity of the lineage represented by the specimens examined here, which is independent from L. boryana in terms of phylogeny (Fig. 6), we combine A. ixodes in Lentinula as a distinct species. Unfortunately, we could not examine the type specimens of A. ixodes, L. boryana and L. guarapiensis, as the specimens could not be loaned.
Lentinula ixodes is also morphologically similar to L. guarapiensis (Speg.) Pegler, which is known only from its type material [Apr. 1879, Balansa 3373 (K, PC)] from Guarapí, Paraguay (Pegler, 1983a). Pegler (1983a) provided a detailed description of the species, which included the original morphological description (Spegazzini, 1883) combined with an examination of the type material currently located in the K and PC herbaria. Although limited by a lack of information on some microstructures due to the specimen's condition (heavily infected with mold and hymenophores covered with glue), this description is quite useful for comparison with L. ixodes (Supplementary Material, Table S2). Lentinula guarapiensis is somewhat similar in the basidiomata habit, with a pileus laterally attached to a curved, cylindrical stipe (lateral or nearly lateral), but the illustration in Pegler (1983a) shows an eccentric stipe (although L. ixodes also has a central stipe). In the first type review, Singer (1952) observed very scarce and inconspicuous spores among mold conidia; these were undoubtedly small (although no size information was provided), hyaline, and cylindrical, with small (although no dimensions were provided) basidia. Pegler (1983a) reported likely authentic small basidiospores (4-4.5 × 2.3-3 μm) compatible with the size of the basidia (12-14 × 3-3.5 µm), both of which were smaller than those of L. ixodes. Although found among numerous mold spores, the shape of the authentic basidiospores of L. guarapiensis seems typical for the group, as illustrated by Pegler (1983a, Fig. 2L). Pegler also observed that the hyphae of the pileus trama in L. guarapiensis has walls up to 1.5 µm thick and a cutis of interwoven, thin-walled hyphae forming the pileipellis, whereas the walls of the hyphae in the pileus trama of L. ixodes are thicker (0.7-3.6 µm), often rough, and incrusted with excrescences. No cheilocystidia were observed in L. guarapiensis and the lamellar edge was deemed likely fertile (Singer, 1952; Pegler, 1983a), in contrast to the conspicuous cheilocystidia in L. ixodes. Lentinula guarapiensis also differs in that it has adnexo-sinuate lamellae with a short decurrent tooth and denticulate edge (Pegler, 1983a). Lentinula guarapiensis is originally from a more distant site in the south of the
continent, and is not in the same biome. Therefore, the examined collections are not topotypical and are not ideal to represent L. guarapiensis. Even if A. ixodes and Agaricus guarapiensis (≡ Lentinula guarapiensis) were conspecific, A. ixodes is the older name and has nomenclatural priority. Guzmán et al. (1997) suspected that L. guarapiensis is also a synonym of L. boryana.
Lentinula lateritia, originally described from Australia (Berkeley, 1881), is morphologically similar but has a distant and quite discontinuous biogeographic distribution from L. ixodes (Pegler, 1983a). The pileus of L. lateritia is smooth and glabrous, neither squamous nor fissured, cinnamon-rufous to hazel, with a darker to cinnamon brown center and lamellae that are adnexo-sinuate and readily separate from a stipe that is central to eccentric, often curved, slender and subcylindrical (Pegler, 1983a). Lentinula lateritia also forms much larger basidiomata, with 20-50(-80) mm diam. pileus and 25-70 × 3-9 mm stipe, which occasionally shows a purplish brown tint and traces of veil at the ring zone (Pegler, 1983a). The lamellae of L. lateritia develop fulvous or purplish brown coloration at maturity, whereas no change in pigmentation occurs in the lamellae of L. ixodes. The pileus context in L. lateritia may reach 6 mm in thickness at the disc but is very thin at the margin, consisting of loosely woven, thick(1-3 μm)-walled hyphae similar to those of L. ixodes. Based on Pegler (1983a), the basidiospores in L. lateritia are similar in size but slightly broader [5.3-6(-6.5) × 3-3.7 μm] than those of L. ixodes, while the cheilocystidia are quite similar but smaller (21-24 × 7.5-10 μm vs. 14.7-33 × 6.6-12.7 µm). The pileipellis in L. lateritia is a cutis of repent, somewhat radially parallel hyphae with pale reddish brown walls, rather than a cutis of disorganized hyphae, which are mostly non-radially oriented, brown to ochraceous brown, or pale yellowish to brownish in color (Pegler, 1983a).
Lentinula ixodes is a unique clade within the Lentinula phylogeny, closer to L. raphanica (Fig. 6) as its sister species. Based on these analyses, L. ixodes is certainly not conspecific with L. boryana sensu Looney et al. (2021), and is therefore not a synonym. Although we used the same nuclear rDNA ITS dataset for the ingroup as Looney et al. (2021), our analyses yielded trees with differing topology. The resolution of their African-American clade (L. aciculospora, L. boryana, L. madagasikarensis, and L. raphanica) was inconclusive in our analyses (Fig. 6), as the small clade L. madagasikarensis + L. aciculospora was sister with moderate support to their Asian-Australasian clade strongly supported herein. Lentinula boryana was sister to L. raphanica with strong support in Looney et al. (2021); here, it formed an unsupported clade as sister to clade L. raphanica + L. ixodes. The high statistical support indicates a solid relationship, and a long-branch effect for L. raphanica + L. ixodes is quite unlikely. Lentinula ixodes anticipated in Menolli Jr. et al. (2022) refers to the combination proposed in this paper and their results from the phylogenetic analyses including ITS sequence from this study corroborate with the relationship between L. raphanica and L. ixodes presented herein.
Although phylogenetically sister and sympatric, L. ixodes is sexually incompatible with L. raphanica, and is therefore a distinct biological species. Similar complete intersterility was found among L. boryana (Group-I), L. raphanica (Group-II), L. aciculospora, and L. edodes (Mata & Petersen, 2000; Mata, Petersen, & Hughes, 2001). In contrast, L. lateritia, L. novaezelandiae, and L. edodes were found to interbreed with each other, and were therefore considered to constitute a single biological species (Shimomura, Hasebe, Nakai-Fukumasa, & Komatsu, 1992; Guzmán et al., 1997). However, these three species have since been recognized as distinct based on molecular phylogeny (Hibbett et al., 1995, 1998; Looney et al., 2021), supporting their morphology-based recognition (Pegler, 1983a). In this paper, we demonstrated that despite being collected in the same place, in the same day and in the same substrate of 2018 and 2021, L. ixodes is distinct from the L. raphanica in the morphological, molecular phylogenetic and biological species concepts.
5. Conclusion
Agaricus ixodes is not a synonym of Lentinula boryana, but rather an independent species. With new collections, Lentinula ixodes is the second unique lineage of the genus in the Amazon Forest to be morphologically described, phylogenetically evaluated, and species-level determined within Lentinula. The species is now confirmed from Guyana to the Amazonas State, Brazil. Here, its mating type was determined and it was crossed with sympatric L. raphanica strains for the first time, confirming its reproductive isolation. The morphology of L. ixodes suggests affinity with the L. boryana complex (Pegler's species concept), which likely explains why it has long been considered a synonym. The supposed viscid pileus surface is misleading in the protologue, as the pileus may appear so when soaked, but is ceraceous or dry in dried conditions. This agrees with Singer's observation that the pileipellis was never gelatinized in the type specimen. The rare and occasional, very long basidiospores in the L. ixodes examined specimens herein, along with the typical spore size, is a notable novelty in this group that merits further investigation.
Acknowledgements
J.J.S. de Oliveira thanks the Programa de Capacitação Institucional (PCI) of the Conselho Nacional de Desenvolvimento Científico Tecnológico and the Programa Nacional de Pós-doutorado (PNPD) of the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the post-doctoral scholarships. R. Vargas-Isla thanks the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) for a DCT fellowships (proc. no. 062.00219/2020), and T.S. Cabral thanks PNPD/CAPES (88882.317512/2019-01) for post-doctoral fellowships. N. Menolli Jr. thanks the Conselho Nacional de Desenvolvimento Científico e Tecnologico (CNPq), Brazil (Research Productivity grant 314236/2021-0) The authors thank Financiadora de Estudos e Projetos (FINEP) for the funding the project nº 01.18.0117.00 “Modernização do acervo do Herbário do INPA e suas capacidades para estudos de taxonomia”. The authors also thank FAPEAM (“Nº Processo 01.02.016301.03240/2021-32” of the “Edital Nº 007/2021 - BIODIVERSA” and “Nº Processo 01.02.016301.03246/2021-00” of the “Edital N. 008/2021 - PROSPAM”), Centro de Estudos Integrados da Biodiversidade Amazônica (INCT-CENBAM), CNPq, the Biodiversity Research Program (PPBio), “Fundação de Amparo à Pesquisa do Estado de São Paulo” (FAPESP) grant #2018/15677-0, Sergio Vergueiro (Agropecuária Aruanã S/A), Felipe Schaedler (Restaurante Banzeiro). Samuel M. Benzecry for finantial support. The authors thank the INPA Herbarium and the anonymous peer-reviewers.
References
- Alvares, C. A., Stape, J. L., Sentelhas, P. C., De Moraes Gonçalves, J. L., & Sparovek, G. (2013). Köppen's climate classification map for Brazil. Meteorologische Zeitschrift, 22, 711-728. https://doi.org/10.1127/0941-2948/2013/0507
- Berkeley, M. J. (1881). Australian fungi - II. Journal of the Linnean Society of London, Botany, 18, 383-389.
- Berkeley, M. J., & Curtis, M. A. (1869). Fungi Cubenses (Hymenomycetes). Journal of the Linnean Society, 10, 280-392.
- Braga-Neto, R., Luizão, R. C. C., Magnusson, W. E., Zuquim, G., & Castilho, C. V. (2008). Leaf litter fungi in a Central Amazonian forest: the influence of rainfall, soil and topography on the distribution of fruiting bodies. Biodiversity and Conservation, 17, 2701-2712. https://doi.org/10.1007/s10531-007-9247-6
- Capelari, M., Asai, T., & Ishikawa, N. K. (2010). Occurrence of Lentinula raphanica in Amazonas State, Brazil. Mycotaxon, 113, 355-364. https://doi.org/10.5248/113.355
- ClimaTempo (2021). Climatologia em Itacoatiara, BR. A StormGeo Company. Retrieved in Apr 13, 2022, from https://www.climatempo.com.br/climatologia/20/itacoatiara-am
- Corner, E. J. H. (1996). The agaric genera Marasmius, Chaetocalathus, Crinipellis, Heimiomyces, Resupinatus, Xerula and Xerulina in Malesia. Beiheft Nova Hedwigia, 111, 1-164.
- Earle, F. S. (1909). The genera of North American gill fungi. Bulletin of the New York Botanical Garden, 5, 373-451.
- Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5, 113. https://doi.org/10.1186/1471-2105-5-113
- Grigoriev, I. V., Cullen, D., Goodwin, S. B., Hibbett, D., Jeffries, T. W., Kubicek, C. P., Kuske, C., Magnuson, J. K., Martin, F., Spatafora, J. W., Tsang, A., & Baker S. E. (2011). Fueling the future with fungal genomics. Mycology, 2, 192-209. https://doi.org/10.1080/21501203.2011.584577
- Grigoriev, I. V., Nikitin, R., Haridas, S., Kuo, A., Ohm, R., Otillar, R., Riley, R., Salamov, A., Zhao, X., Korzeniewski, F., Smirnova, T., Nordberg, H., Dubchak, I., & Shabalov, I. (2014). MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic Acids Research, 42, D699-D704. https://doi.org/10.1093/nar/gkt1183
- Guzmán, G., Salmones, D., & Tapia, F. (1997). Lentinula boryana: morphological variations, taxonomic position, distribution and relationships with Lentinula edodes and related species. The Reports of the Tottori Mycological Institute, 35, 1-28.
- Hibbett, D. S. (2001). Shiitake mushrooms and molecular clocks: historical biogeography of Lentinula. Journal of Biogeography, 28, 231-241. https://doi.org/10.1046/j.1365-2699.2001.00528.x
- Hibbett, D. S., Fukumasa-Nakai, Y., Tsuneda, A., & Donoghue, M. J. (1995). Phylogenetic Diversity in Shiitake Inferred from Nuclear Ribosomal DNA Sequences. Mycologia, 87, 618-638. https://www.jstor.org/stable/3760806
- Hibbett, D. S., Hansen, K., & Donoghue, M. J. (1998). Phylogeny and biogeography of Lentinula inferred from an expanded rDNA dataset. Mycological Research, 102, 1041-1049. https://doi.org/10.1017/S0953756297005996
- Instituto de Pesquisas e Experimentação Agropecuárias do Norte (1969). Os solos da área Manaus-Itacoatiara. Séries Estudos e Ensaios nº 1, Edição do Setor de Relações Públicas. Rio de Janeiro: Impressora POLAR.
- Küppers, H. (2002). Atlas de los colores. Barcelona: Editorial Blume.
- Lodge, D. J., Padamsee, M., Matheny, P. B., Aime, M. C., Cantrell, S. A., Boertmann, D., Kovalenko, A., Vizzini, A., Dentinger, B. T. M., Kirk, P. M., Ainsworth, A. M., Moncalvo, J.-M., Vilgalys, R., Larsson, E., Lücking, R., Griffith, G. W., Smith, M. E., Norvell, L. L., Desjardin, D. E., … & Hattori, T. (2014). Molecular phylogeny, morphology, pigment chemistry and ecology in Hygrophoraceae (Agaricales). Fungal Diversity, 64, 1-99. https://doi.org/10.1007/s13225-013-0259-0
- Looney, B. P., Buyck, B., Menolli Jr. N., Randrianjohany, E., & Hibbett, D. (2021). Lentinula madagasikarensis sp. nov., a relative of shiitake mushrooms from Madagascar. Fungal Systematics and Evolution, 92, 881-882. https://doi.org/10.3114/fuse.2021.08.01
- Mata, J. L., & Petersen, R. H. (2000). A new species of Lentinula (Agaricales) from Central America. Mycoscience, 41, 351-355. https://doi.org/10.1007/BF02463948.
- Mata, J. L., & Petersen, R. H. (2001). Type specimens studies in new world Lentinula. Mycotaxon, 79, 217-229.
- Mata, J. L., Petersen, R. H., & Hughes, K. W. (2001). The genus Lentinula in the Americas. Mycologia, 93, 1102-1112. https://doi.org/10.2307/3761671
- Matheny, P. B., Curtis, J. M., Hofstetter, V., Aime, M. C., Moncalvo, J.-M., Ge, Z. W., Yang, Z. L., Slot, J. C., Ammirati, J. F., Baroni, T. J., Bougher, N. L., Hughes, K.W., Lodge, D. J., Kerrigan, R. W., Seidl, M. T., Aanen, D. K., DeNitis, M., Daniele, G. M., Desjardin, D. E., … Hibbett, D. S. (2006). Major clades of Agaricales: a multilocus phylogenetic overview. Mycologia, 98, 984-997. https://doi.org/10.3852/mycologia.98.6.982
- Menolli Jr., N., Sánchez-Ramírez, S., Sánchez-García, M., Wang, C., Patev, S., Ishikawa, N. K., Mata, J. L., Lenz, A. R., Vargas-Isla, R., Liderman, L., Lamb, M., Nuhn, M., Hughes, K. W., Xiao, Y., & Hibbett, D. S. (2022). Global phylogeny of the Shiitake mushroom and related Lentinula species uncovers novel diversity and suggests an origin in the Neotropics. Molecular Phylogenetics and Evolution, 173, 107494. https://doi.org/10.1016/j.ympev.2022.107494
- Miles, P. G., Takemaru, T., & Kimura, K. (1966). Incompatibility factors in the natural population of Schizophyllum commune. I. Analysis of the incompatibility factors present in fruit bodies collected within a small area. Botanical Magazine (Tokyo), 79, 693-705.
- Moncalvo, J. -M., Lutzoni, F., Rehner, S. A., Johnson, J., & Vilgalys, R. (2000). Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Systematic Biology, 49, 278-305.
- Moncalvo, J. -M., Vilgalys, R., Redhead, S. A., Johnson, J. E., James, T. Y., Aime, M.C., Hofstetter, V., Verduin, S. J. W., Larsson, E., Baroni, T. J., Thorn, R. G., Jacobsson, S., Clémençon, H., & Miller Jr., O. K. (2002). One hundred and seventeen clades of euagarics. Molecular Phylogenetics and Evolution, 23, 357-400. https://doi.org/10.1016/S1055-7903(02)00027-1
- Montagne, J. P. F. C. (1849). Sixièmes Centurie de plantes cellulaires nouvelles, tant indigènes qu'exotiques. Décade VII. Annales des Sciences Naturelles Botanique, 11, 235-246.
- Montagne, J. P. F. C. (1854). Cryptogamia Guyanensis seu plantarum cellularium in Guyana gallica annis 1835-1849 a cl. Leprieur collectarum enumeratio universalis. Annales des Sciences Naturelles Botanique, 1, 91-144.
- Nylander, J. A. A. (2004). Mr. Modeltest v2. Program distributed by the author. Uppsala: Evolutionary Biology Centre, Uppsala University.
- Oliveira, J. J. S., Vargas-Isla, R., Cabral, T. S., Rodrigues, D. P., & Ishikawa, N. K. (2019). Progress on the phylogeny of the Omphalotaceae: Gymnopus s. str., Marasmiellus s. str., Paragymnopus gen. nov. and Pusillomyces gen. nov. Mycological Progress, 18, 713-739. https://doi.org/10.1007/s11557-019-01483-5
- Pegler, D. N. (1975). The classification of the genus Lentinus Fr. (Basidiomycota). Kavaka, 3, 11-20.
- Pegler, D. N. (1983a). The genus Lentinula (Tricholomataceae tribe Collybieae). Sydowia, 36, 227-239.
- Pegler, D. N. (1983b). Agaric flora of the Lesser Antilles. Kew Bulletin, Additional Series, 9, 1-668.
- Pegler, D. N. (1986). Agaric flora of Sri Lanka. Kew Bulletin, Additional Series, 12, 1-519.
- Pegler, D. N. (1997). The Agarics of São Paulo, Brazil: an account of the agaricoid fungi (Holobasidiomycetes) of São Paulo State, Brazil. Royal Botanic Gardens, Kew.
- Prance, G. T. (1978). The origin and evolution of the Amazon flora. Interciencia, 3, 207-222.
- Resende, A. F., Piedade, M. T. F., Feitosa, Y. O., Andrade, V. H. F., Trumbore, S. E., Durgante, F. M., Macedo, M. O. & Schöngart, J. (2020). Flood-pulse disturbances as a threat for long-living Amazonian trees. New Phytologist, 227, 1790-1803. http://dx.doi.org/10.1111/nph.16665
- Ronquist, F., Teslenko, M., van der Mark P., Ayres, D., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M. A., & Huelsenbeck, J. P. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61, 539-542. http://dx.doi.org/10.1093/sysbio/sys029
- Sanuma, O. I., Tokimoto, K., Sanuma, C., Autuori, J., Sanuma, L. R., Sanuma, M., Martins, M. S., Menolli Jr., N., Ishikawa, N. K., & Apiamö, R.M. (2016). Sanöma samakönö sama tökö nii pewö oa wi ĩ tökö waheta: Ana amopö = Enciclopédia dos alimentos Yanomami (Sanöma): Cogumelos. São Paulo: Hutukara Associação Yanomami e Instituto Socioambiental.
- Shimomura, N., Hasebe, K., Nakai-Fukumasa, Y., & Komatsu, K. (1992). Intercompatibility between geographically distant strains of shiitake. Reports of the Tottori Mycological Institute, 30, 26-29.
- Singer, R. (1941). Is Shiitake a Cortinellus? Mycologia, 33, 449-451.
- Singer, R. (1952 [1951]). Type studies on agarics: III. Lilloa, 25, 464-551.
- Singer, R. (1955). Type studies on Basidiomycetes: VIII. Sydowia, 9, 367-431.
- Singer, R. (1989). New taxa and new combinations of Agaricales (Diagnoses fungorum novorum agaricalium IV). Fieldiana Botany, 21, 1-133.
- Spegazzini, C. L. (1883). Fungi Guaranitici. Pugillus 1. Anales de la Sociedad Científica Argentina, 16, 242-248.
- Stamatakis, S. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22, 2688-2690. https://doi.org/10.1093/bioinformatics/btl446
- Vargas-Isla, R., Capelari, M., Menolli Jr., N., Nagasawa, E., Tokimoto, K., & Ishikawa, N. K. (2015). Relationship between Panus lecomtei and P. strigellus inferred from their morphological, molecular and biological characteristics. Mycoscience, 56, 561-571. http://dx.doi.org/10.1016/j.myc.2015.05.004
- Vargas-Isla, R., Ishikawa, N. K., & Py-Daniel, V. (2013). Contribuições etnomicológicas dos povos indígenas da Amazônia. Biota Amazônia, 3, 58-65.
- Vasco-Palacios, A. M., Suaza, S. C., Castaño-Betancur, M., & Franco-Molano, A. E. (2008). Conocimiento etnoecológico de los hongos entre los indígenas Uitoto, Muinane y Andoke de la Amazonía Colombiana. Acta Amazonica, 38, 17-30. http://dx.doi.org/10.1590/S0044-59672008000100004
- Wilson, A. W., & Desjardin, D. E. (2005). Phylogenetic relationships in the gymnopoid and marasmioid fungi (Basidiomycetes, euagarics clade). Mycologia, 97, 667-679. https://doi.org/10.3852/mycologia.97.3.667
