Use of Pawpaw (Carica papaya) Seed in Tilapia Sex Reversal
2020 Volume 8 Pages 230-242
Details
2020 Volume 8 Pages 230-242
Tilapia breeds effortlessly in captivity, with this attribute which is considered as the “Achilles heel” of the species, because it predisposes pond systems to overcrowding and low weight at harvest. Efforts to mitigate this shortcoming include mono-sex culture using exogenous hormone to reverse the sex of sexually undifferentiated fish. This is premised on the fact that improvement in the growth by mono-sex culture will lead to shortened production times and a more uniform weight at harvest, which will ultimately benefit the producers. However, the use of exogenous hormones in aquaculture has recently raised concerns about the effect on farm workers, consumers and on the environment. Recently research has focused on the use of substances of plants origin which mimic the action of hormones as a potential approach to achieve sex reversal in fish. Pawpaw (Carica papaya) seed contains phytochemicals that hold great promise as a sex reversal and a reproductive inhibition agent in aquaculture. The purpose of this study was to conduct a systematic review of the relevant literature on the possibility of using a less harmful phytochemicals abundant in the plant to effect sex reversal of sexually undifferentiated tilapia.
Cichlids are a highly diversified group of fish and include amongst others tilapia, which is one of the most important fish species in freshwater aquaculture (Siddiqui and Al-harbi, 1995). Because of its widespread occurrence, growth on natural grazing or formulated feeds with no constraint for seed production, disease resistance and high consumer acceptability, cichlid species is extremely suitable as a food fish species that can potentially alleviate food security in rural communities in Sub-Saharan Africa (Little, 1998). Tilapia species are hardy and highly prolific, and breeds effortlessly in captivity (Reed et al., 1967). In tilapia aquaculture, the consequence of early maturity on the overall reproductive performance is of great importance in culture systems. Precocious maturation and indiscriminate breeding result in overcrowding of ponds and stunted growth, which in turn results in a low percentage of marketable size fish obtained in mixed sex culture systems (Toguyeni, et al., 2002). Implementing all-male tilapia production systems offers a potential means to overcome the problems associated with overstocking and stunted growth, and is based on the premise that male tilapia grows faster and bigger than females (Beardmore et al., 2001). The techniques employed in tilapia production systems to achieve all-male populations range from manual separation of sexes, genetic/chromosomal manipulations (super male tilapia), environmental manipulation (such as heat shock), and endocrine manipulation of gender (including administration of 17-methyltestosterone) (Abucay et al., 1999; Beardmore et al., 2001; Desprez et al., 2003; Abad et al., 2007). The endocrine manipulation of fish gender is the most effective method to create all-male tilapia populations (Beardmore et al., 2001) and subsequent increase in overall fish production, but the use of the meat is prohibited in many countries since hormone residues remaining in the meat may adversely affect the health of consumers, farm workers and on the environment (Curtis et al., 1991; Khalil et al., 2011). Apart from the suspected health hazards of employing synthetic hormones in food fish production, concerns have also been expressed on the fact that they are expensive and difficult to obtain (Jegede, 2010). Because of these reasons efforts are being made to substitute synthetic hormones with less harmful and environmental friendly plants sources to manipulate the gender of fish (Omeje et al., 2018). Carica papaya seed meal is one of the plants phytochemicals that have attracted research in this regard (Ampofo-yeboah, 2013).
By improving our understanding of the use of pawpaw seed powder as part of basal diets of tilapia to skew the gender of production populations, we will be able to formulate a treatment protocol for tilapia production systems that can be used by rural communities to farm profitably with tilapia, thus addressing food and household security. It will also assist in minimizing the negative effect of exogenous hormones on the environment, thereby ensuring the production of a sustainable and safe food source, without compromising the environment and human health.
Commercial fish farming especially in low income food deficit countries (LIFDC) for example Benin, Ghana, Nigeria, Sri Lanka and Bangladesh, have been recognized as an important tool to increase food fish availability and accessibility, and also income generation through employment (Hishamunda and Ridler, 2006). Although aquaculture practices in developing countries are subsistence in nature and may not provide substantial employment to the teeming population, its impact in poverty alleviation cannot be ignored. Fish contributes over 25% of total animal protein intake worldwide especially in low income and developing countries (Bondad-Reantaso et al., 2005). It is a good source of vitamins especially A, D, E and B- complex vitamins and also omega-3 fatty acids (Bondad-Reantaso et al., 2005). Tilapia species is one of the most cultured food fish worldwide, second only to carp (Coward and Bromage, 2000).
Tilapia refers to several freshwater species that belong to the family Cichlidae (Coward and Bromage, 2000). Tilapia originates from Africa, and because of its easy adaptability to various environmental conditions, has been introduced to many countries. The low cost of production combined with the fact that tilapia is widely accepted by consumers as a food fish, promotes its culture worldwide (Siddiqui and Al-harbi, 1995; De la Fuente et al., 1999; Rad et al., 2006; Shalloof and Salama, 2008; El-Kashief et al., 2013). Statistics generated by the Food and Agriculture Organization (FAO) has indicated that tilapia production in 2001 amounted to 1.5 million tonnes with an expectation to increase progressively over the years (FAO, 2002). World tilapia production for the year 2012 exceeded 4.51 million metric tonnes (FAO, 2014) and by 2017 it has increased to 6.5 million tonnes (Figure 1).
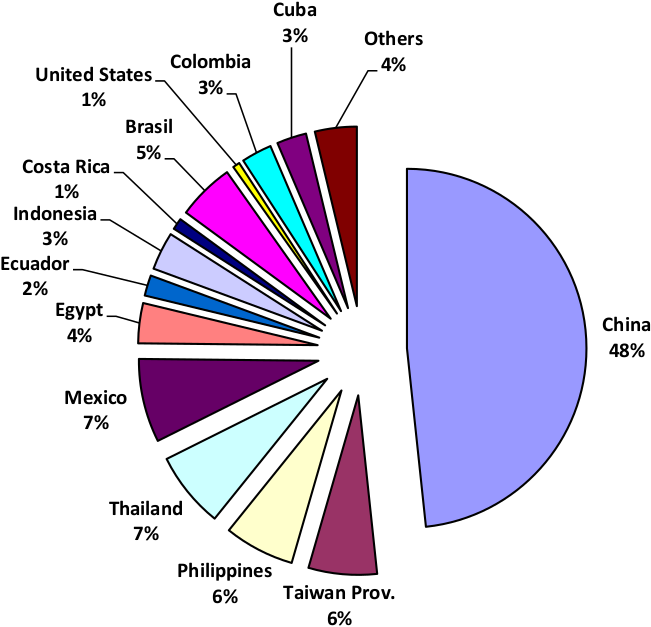
Figure 1: World Tilapia production of 6.5 million tonnes in 2017 (adapted from FAO Fisheries and Aquaculture Department, 2018)
When Africa is compared to other countries that farm with tilapia, e.g. countries like China, the Philippines, Taiwan, Thailand, USA and Belgium, Africa is outperformed in terms of production, with these countries collectively producing more than 850,000 tonnes annually (Coward and Bromage, 2000). Tilapia production in 2010 exceeded 3.2 million metric tons, exceeding the production of both salmon and catfish. Although second to carp in terms of production, it is expected that in the near future with the greater acceptance and wider distribution enjoyed by tilapia, it will become the most important aquaculture species. In Africa, Egypt is perhaps the first country to venture into the culture of fish through using freshwater ponds for fish production activities (Bondad-Reantaso et al., 2005). Incidentally tilapia was the first cultured fish in Egypt, about 2500 years ago, and today Egypt is considered as the biggest producer of food fish, contributing 72% of Africa’s production (Brummett and Williams, 2000; FAO, 2010). According to the FAO (2010), Nigeria and Uganda is the second and third biggest producers, producing 16% and 7% respectively, of Africa’s production. Together these three countries produce about 94% of the entire continent’s production as indicated in Table 1.
| Ranking | Country | Production (tons) | Percentage |
| 1 | Egypt | 919,585 | 71.51 |
| 2 | Nigeria | 200,535 | 15.59 |
| 3 | Uganda | 95,000 | 7.39 |
| 4 | Kenya | 12,154 | 0.95 |
| 5 | Zambia | 10,290 | 0.80 |
| 6 | Ghana | 10,200 | 0.79 |
| 7 | Madagascar | 6,886 | 0.54 |
| 8 | Tunisia | 5,424 | 0.42 |
| 9 | Malawi | 3,163 | 0.25 |
| 10 | South Africa | 3,133 | 0.24 |
| 11 | Democratic Republic of the Congo | 2,970 | 0.23 |
| 12 | Zimbabwe | 2,702 | 0.21 |
| 13 | Sudan | 2,200 | 0.17 |
| 14 | Mali | 2,083 | 0.16 |
| 15 | Algeria | 1,759 | 0.14 |
| 16 | Cote d’Ivoire | 1,700 | 0.13 |
| 17 | Rocco | 1,522 | 0.12 |
| 18 | Mozambique | 864 | 0.07 |
| 19 | Cameroon | 628 | 0.06 |
| 20 | Rwanda | 628 | 0.05 |
| 21 | Others | 2 339 | 0.18 |
| TOTAL PRODUCTION | 1,285,972 | 100 | |
The family Cichlidae has three main genera Oreochromis, Tilapia and Sarotherodon though the members of the family are generally referred to as tilapia. Even though the family is made up of more than 80 species of fish, only 8 or 9 species are considered important in terms of production (Coward and Bromage, 2000) with three species in the genus Oreochromis (O. niloticus, O. mossambicus, and O. aureus), two species in the genus Tilapia (T. rendalli and T. zilli), and one species in the genus Sarotheridon (S. galilaeus) being the most cultivated of the family (Siddiqui and Al-harbi, 1995; El-Kashief et al., 2013). Tilapias are considered as a good source of food fish especially for the low income food deficit countries. Previously all tilapia fish were grouped together under the genus Tilapia, but recently two other groups were established based on the parental care investment of the particular species. The genus Oreochromis exhibit maternal mouth brooding, the genus Sarotherodon is characterized according to the mouth brooding behaviour exhibited by both parents, and the Tilapia genus are classified as substrate spawners (Coward and Bromage, 2000; Specker and Kishida, 2000; Fishelson and Bresler, 2002). Fecundity of tilapia varies inversely with the parental care exhibited by the species, which means that the more the parental care is invested, the lower the fecundity of the species will be. The fecundity of species as O. mossambicus, which is a mouth brooder, is as low as 350, compared to substrate spawners such as Tilapia zilli, which has a fecundity of 12,000 eggs (Coward and Bromage, 2000). Members of the Tilapia family exhibit various degrees of parental care for their offspring. The genus Oreochromis orally incubates eggs and larvae, and the mouth brooding practice continues up to juvenile stage where the female at any threat or danger, takes the young into her mouth for safety (Tacon et al., 1996).
The reproductive performance of a species is of particular interest in aquaculture, since production indices are based on the availability, and number and quality of sperm and ova produced. Since egg size increases with maternal age and size, fish maturing early and at a smaller size will produce relatively more but smaller eggs per unit body weight than larger fish (Coward and Bromage, 1999). There is a decrease in the growth rate during the time of sexual maturity, and as a consequence of this, late maturing fish are larger than their early maturing counterparts (Schreibman et al., 1989). Species like tilapia exhibit early sexual maturation, which result in the overcrowding of and stunted growth observed in tilapia culture systems. Tilapia species become sexually mature at a very small size, i.e. with an average live weight of 15 g, with no growth occurring after an animal has attained sexual maturity (Popma and Lovshin, 1995). Maternal size is one of the factors believed to be responsible for egg size. Within a given fish species, the production of larger eggs by bigger individuals is well documented (Zonneveld and Van Zon, 1985). However, it is not clear whether maternal age or size is the primary factor influencing egg size. Al-Ahmad et al. (1988) stated that the age of brood stock does not influence the fecundity of tilapia, whereas fecundity decreases with an increase in the salinity of the culture water. Egg size is influenced by the species of fish, i.e. in the tilapia family egg size is species-specific, regardless of female age. Rana (1988) reported that when females of similar age and irrespective of size, were reared and spawned under similar conditions, mean egg size of Oreochromis niloticus females was found to be significantly larger than that of Oreochromis mossambicus.
Fecundity has been defined as the number of maturing oocytes in the ovaries prior to spawning (Bagenal, 1978). The number, diameter and volume of eggs produced by female fish increases with their age, length and weight, whereas their relative fecundity which is the number of eggs spawned per kilogram body weight of the female, decreases (Coward and Bromage, 1999). In fish of the same weight and total length, the size of their eggs may vary (Pena-Mendoza et al., 2005). Rana (1988) was of the opinion that total fecundity is more closely related to maternal size than age. According to the author, unlike egg size, the number of eggs spawned by Oreochromis niloticus brooders of similar age, increased significantly with their size. Consequently in a mixed age structure, studies suggest that larger fish of the same age may lay more eggs. Maternal growth rate and factors such as their nutritional status will also be of crucial importance in increasing egg yields for fry production. Variation in the egg size of the same individual fish indicates the multiple spawning natures of the species.
However, despite the good culture potential of the tilapias as a food fish, precocious maturation and indiscriminate breeding in mixed sex populations result in overcrowding of ponds and stunted growth of the fish (Toguyeni et al., 2002). The concept of mono-sex culture production of tilapia presents an opportunity to overcome the limitation of precocious breeding, and it is further supported by the hypothesis that male tilapia grows faster than females (Otubusin, 1988; Davis et al., 2010). The establishment of all-male tilapia populations can also prevent the problem of undesired indiscriminate spawning, which will contribute to minimizing stunted growth, and thus optimizing the time to harvest. Various methods are available for the production of all-male tilapia populations as shown in Table 2.
| Method | Description | References |
| Chromosomal manipulation | Polyploidy | Piferrer et al., 2009 |
| Manual sexing | Separating the fish according to sex | Pandian and Varadaraj, 1987 |
| Hybridization | Offspring of crosses between two related species | Pruginin et al., 1975 |
| Genetic manipulation | YY super male technology | Mair et al. 1997 |
| Environmental temperature manipulation | Heat shock (masculinization) Cold shock (feminization) |
D’Cotta et al., 2001 Wang and Tsai, 2000 |
| Environmental pH manipulation | Low pH (masculinization) High pH (feminization) |
Guerrero-Este’ves and Moreno-Mendoza, 2010 |
| Administration of hormones | 17α-methyltestosterone, 19-norethynyltestosterone, 11-ketotestosterone (Masculinization) 17β-estradiol, estrone and di-ethylstilbesterole (Feminization) |
Yamazaki, 1983; Vera Cruz and Mair, 1994; Pandian and Varadaraj, 1987; Rosenstein and Hulata, 1993 |
Sex reversal converts genotypic females into phenotypic males (Toguyeni et al., 2002) or vice versa. In masculinization also known as all-male or mono-sex production, the genetic females are induced to develop as functional males but the genetic males are not affected and therefore develop normally as functional males (Pandian and Varadaraj, 1987). Also in all female monosex production (feminization), the genetic males are induced to develop as functional females but the genetic females are not affected and so develop normally as functional females. Hormonal sex reversal or inhibition of gonadal growth is achieved through application of steroid before sexual differentiation or during the period of gametogenesis. Sex differentiation in fish is the change from primordial germ cells (gonads) to either ovary or testes (Viñas et al., 2013). According to Afonso et al. (2001), male and female sex differentiation is modulated by androgen and estrogen hormones, respectively. Sex reversal can be achieved by the application of hormones during the sexually undifferentiated stage (Rothbard et al., 1987). Since sex reversal processes takes place during the period of sex differentiation the sex reversed fish is permanent. Sex reversal processes are undertaken in food fish production to favour the sex that has the greatest growth potential which will ultimately lead to greater profitability of the enterprise. For example, the male grows bigger in cichlids, while in the salmonids and cyprinids it is the females that exhibit greatest growth (Piferrer, 2001). Male tilapia grows faster than their female counterparts and therefore all male culture is usually preferred when production of food fish is the primary concern (Macintosh et al., 1985). Many factors have been attributed with the differential growth capabilities between the male and female tilapia, these factors include diversion of greater percentage of metabolic energy to breeding processes in females and androgens in male having higher anabolic effects (Toguyeni et al., 2002). The advantages of production of monosex tilapia include elimination of reproduction, increased growth rate and possible reduction of variation in harvest size (Beardmore et al., 2001).
Carica papaya Linn (Caricaceae family) popularly called pawpaw, is widely available all year round in sub-Saharan Africa and other tropics and subtropics worldwide (Oliveira and Vitoria, 2011). The species originated from Southern Mexico and Nicaragua (Chan and Paul, 2008). Brazil is reported to be the world’s biggest producer followed by Mexico and Nigeria in the second and third place respectively while Europe and USA are the principal markets for its consumption (Oliveira and Vitória, 2011). The popularity of the plant is due to its fruit which when ripe are yellow and succulent and very nutritive and is available throughout the year (Emeruwa, 1982). Apart from the nutritive aspect, different part of C. papaya tree and even the seeds have been known to possess a wide range of pharmacological properties especially antibacterial, antifungal, pesticidal and anti-fertility (Eno et al., 2000; Doughari et al, 2007). Doughari et al. (2007) reported antibacterial activity of bioactive compounds of root extract of pawpaw against some pathogenic bacteria, they opined that the extract may be used to treat gastroenteritis, urethritis, otitis media, and typhoid fever and wound infections. Emeruwa (1982) agreed that fruit and seed extracts of pawpaw has antibacterial properties. It is also claimed that the fruit juice of pawpaw has a depressant effect on the blood pressure (Eno et al., 2000). Seed powder of C. papaya has also been credited with molluscicidal activities against freshwater snail Lymnaea acuminata (Jaiswal and Singh, 2008). Farias et al. (2007) reported that crude extract of seed of C. papaya exhibited significant inhibitory activity against cowpea weevil (Callosobruchus maculatus). Ayotunde and Ofem (2008) opined that pawpaw seed are poisonous to fish and have been used to control over population in tilapia culture. Mature unripe pulp of pawpaw contains the phytochemicals Saponins, alkaloids, terpenoids, flavonoids, glycosides, steroids and cardenolides which are responsible for its medicinal properties (Oloyede, 2005; Ezike et al., 2009). Likewise, the chemical composition of the seed of pawpaw is as shown in Table 3.
| Constituents of pawpaw seed | Recorded level / concentration | References | |
| 1 | Fatty acid | 1.33% | Puangsri et al., 2005; Nwofia et al., 2012 |
| 2 | Crude Protein | 3.04% | Afolabi et al., 2011; Nwofia et al., 2012 |
| 3 | Crude fibre | 1.87% | Afolabi et al., 2011; Nwofia et al., 2012 |
| 4 | Papaya oil | 9.95% | Afolabi et al., 2011; Aravind et al., 2013 |
| 5 | Carpasamine | 3,500ppm | Olafsdottir et al., 2002; Aravind et al., 2013 |
| 6 | Oleanolic glucoside | No data | Ampofo-Yeboa, 2013; Kumar et al., 2012 |
| 7 | Benzyl isothiocynate | 2,000–5000ppm | Olafsdottir et al., 2002 |
| 8 | Benzyl glucosinate | No data | Bennett et al., 1997 |
| 9 | Benzyl thiourea | No data | Krishna et al., 2008 |
| 10 | Hentriaconatane | No data | Krishna et al., 2008 |
| 11 | β-sitosterol | No data | Krishna et al., 2008, Kumar et al., 2012 |
| 12 | Caricine | No data | Krishna et al., 2008 |
| 13 | Myrosin | No data | Krishna et al., 2008 |
It is believed that the benzyl isothiocyanate, β-sitosterol and oleanolic glucoside component of pawpaw seeds are responsible for the sex reversal and antifertility properties of the plant (Wilson et al., 2002; Jegede and Fagbenro, 2008; Krishna et al., 2008; Kumar et al., 2012; Ampofo-yeboah, 2013). Benzyl isothiocyanate, β-sitosterol and oleanolic glucoside like other bioactive compounds produced by plants are secondary metabolites that are beneficial to the plant that produce it. However, they may have pharmacological, antifertility or toxicological effect on humans and animals including fish. Bioactive compounds produced by plants such as isothiocyanate, β-sitosterol and oleanolic glucoside are referred to as phytochemicals or phytoestrogens. Phytoestrogens are natural products of some cultured and wild plants which have the ability to modulate the endocrine system by exhibiting estrogenic and /or androgenic activity (Rearick et al., 2014). Because they modulate the endocrine system, phytoestrogens are sometimes referred to as endocrine disrupting compounds (EDC). The mode of action of EDC includes interfering with the enzymes responsible for biosynthesis of steroid hormones (Ribeiro et al., 2012). Since the steroid hormones are involved in the regulation of the GnRH-FSH/LH system through positive and negative feedback mechanism (Zohar et al., 2010), these anthropogenic chemicals equally affects the reproductive capacity of an organism by interfering in the neuroendocrine system. Sexually undifferentiated gonads are more prone to disruption by endocrine active chemicals (Van Aerle et al., 2002). The effects of endocrine disrupting chemicals include reduced viability of eggs, high mortality rate of eggs and reduced hatching rate (Schwaiger et al., 2002). There are also reports on the activities of these phytochemicals from various plants on fertility and wellbeing of fish. Dongmeza et al. (2006) reported reduced growth performance in Nile tilapia feed diet containing Moringa oleifera while Makkar et al. (2007) used diet containing 300 ppm of quillaja saponin to suppress reproductive activities in Nile tilapia. According to Casanova-Nakayama et al. (2011), endocrine disrupting compounds exert their biological activity either by interacting with endogenous hormone receptors or by disturbing endogenous hormone metabolism. Endocrine disruption in fish is indicated by the presence of significant level of vitellogenin in males or elevation of its levels in females. Unlike the female fish where vitellogenin is always present and easily detected in the blood especially in sexually matured fish, its presence in the male is usually very insignificant. Vitellogenin synthesized in the liver is incorporated to the growing oocytes where it serves as egg yolk precursor. The level of the vitellogenin in the serum of fish is measured using enzyme linked immuno-sorbent assay (Van Aerle et al., 2002). Also significant increase in the levels of estradiol in males and testosterone in females demonstrates the exposure of fish to anthropogenic chemicals (Schwaiger et al., 2002). Studies have shown that many antifertility compounds contained in parts of plants have the potential to disrupt reproductive endocrine pathways (Biswas et al., 2002; Huang and Chen, 2004), similar to the influence of dopamine.
Alternative treatment strategies that include phytochemicals such as phytoestrogens that mimic the action of the endogenous fish hormones, is considered as a suitable substitute for 17-methyltestosterone to produce all-male tilapia populations (Ampofo-yeboah, 2013). The effect of phytoestrogen exposure in fish includes impairment of the reproductive system in adults and outright sex reversal in larvae (Ampofo-yeboah, 2013). The impairment of the reproductive activities manifests as infertility, reduced fecundity, ovo-testes in females and vitellogenin induction in males (Ribeiro et al., 2012). Estrogenic phytoestrogens are capable of exhibiting anti-estrogenic tendencies in the presence of endogenous estrogens (Clotfelter and Rodriguez, 2006) and that may explain the sex reversal action of some phytoestrogens like those contained in pawpaw seeds. Crude extracts of different parts of pawpaw (Carica papaya) plants have been used for the partial reduction or complete suppression of reproduction in fish (Mousa et al., 2008; Hossam and Wafaa, 2011).
Compared to the literature on the anti-fertility effect of C. papaya on laboratory animals (Joshi and Chinoy, 1996; Udoh and Kehinde, 1999; Adebiyi et al., 2003; Manivannan et al., 2004; Verma et al., 2006), there are relatively few information on the effect of pawpaw seed on the reproductive potential of fish. However, Hossam and Wafaa (2011) used 6 g/kg/day to induce permanent sterility in O. niloticus and 3 g/kg/day to induce reversible sterility. Ekanem and Okoronkwo (2003) used an inclusion level of 9.8 g/kg of pawpaw seed meal per day to induce permanent sterility, and 4.9 g/kg of pawpaw seed meal per day to induce reversible infertility in male Nile tilapia (Oreochromis niloticus). In the study of Ampofo-Yeboah (2013), sexually undifferentiated O. mossambicus received diets containing 15 g of pawpaw seed powder per kg of basal diet, and this inclusion level resulted in 65% males. There is the need to quantify the optimal inclusion level and duration of treatment to establish the dosage that will provide the maximum percentage of sex reversal. Omeje et al. (2018) reported that pawpaw seed meal (PSM) fed sexually undifferentiated fry of O. mossambicus was able to skew the sex ratio in favour of males. At an inclusion level of 10 g/kg of basal diet (BD), PSM was able to skew the sex ratio in favour of males (60% males to 40% females). The proportion of males increased with an increasing dosage of PSM, with the maximum masculinization achieved at an inclusion level of 20 g/kg BD, resulting in 77.8% males produced. In a related research, Iipinge (2019) fed sexually undifferentiated three spotted tilapia (Oreochromis andersonii) PSM at a rate of 5, 15, 25, and 35 g/kg and reported 82% masculinization with no significant difference in the percentage masculinization obtained between 15 and 35 g/kg PSM. In another research, Ugonna et al. (2018) recorded 82.19% masculinization of Nile tilapia (Oreochromis niloticus) fed diet containing 4.27 g/kg PSM. It has also been reported that C. papaya seed included in the diet of pre-vitellogenic tilapia significantly reduced the 17β-estradiol levels, gonad weight, gonado-somatic index, fecundity and egg diameter of females (Abdelhak et al., 2013; Omeje et al., 2019). Most of the researchers (Ekanem and Okoronkwo, 2003; Hossam and Wafaa, 2011; Ampofo-yeboah, 2013; Omeje et al., 2018) reported that the pawpaw seed meal were well tolerated by the fish and lesions (if any) it impacted on the tissues of fish were negligible and also reversible on withdrawal of treatment which shows that PSM is safe for use in tilapia culture. They also opined that using pawpaw seeds as reproductive inhibitor has no negative effect on survival, growth or health status of tilapia.
The use of phytochemicals as a viable alternative to 17-methyltestosterone in the production of monosex tilapia an economically important food fish holds a great prospect. Although Carica papaya seed have been reported to skew the sex ratio in favour of males (Ampofo-yeboah, 2013; Omeje et al., 2018; Ugonna et al., 2018; Iipinge, 2019), the percentage masculinization reported by the authors may not be ideal since as little as 5% females in a culture facility can reproduce indiscriminately thereby mitigating the benefits inherent in monoculture tilapia production. Therefore, there is need for further improvement so as to enhance the percentage masculinization reported by the authors. However, this is promising for fish farmers and aquaculture scientists who may now use cost-effective and environmentally friendly PSM for sex reversal and reproductive inhibition instead of expensive exogenous steroid hormones. The same goes for the consumers of tilapia fish, the benefit of eating tilapia fed harmless PSM instead of exogenous hormones cannot be overemphasized.
The authors express their appreciation to the Stellenbosch University, Western Cape, South Africa for providing the facilities used for the study. Also sincere thanks to the West African Agricultural Productivity Program (WAAPP-Nigeria) for providing the funding.